
Вопрос 50. Мобилизация жира в жировой ткани (липолиз). Триацилглицерол, диацилглицерол и моноацилглицероллипазы. Гормональная регуляция липолиза в адипоцитах.
Адипоциты (место депонирования жиров) располагаются в основном под кожей, образуя подкожный жировой слой, и в брюшной полости, образуя большой и.малый сальники. Мобилизация жиров, т.е. гидролиз до глицерола и жирных кислот, происходит в постабсорбтивный период, при голодании и активной физической работе. Гидролиз внутриклеточного жира осуществляется под действием фермента гормончувствительной липазы — ТАГ-липазы. Этот фермент отщепляет одну жирную кислоту у первого углеродного атома глицерола с образованием диацилглицерола, а затем другие липазы гидролизуют его до глицерола и жирных кислот, которые поступают в кровь. Глицерол как водорастворимое вещество транспортируется кровью в свободном виде, а жирные кислоты (гидрофобные молекулы) в комплексе с белком плазмы — альбумином. Гидролиз триацилглицеролов до жирных кислот и глицерола катализируют три липазы. Первой действует триацилглицероллипаза, затем диацил-глицероллипаза и последней — моноацилглицероллипаза, гидролизующая сложноэфирную связь в моноацилглицеролах. Эти липазы локализованы в разных органоидах. Триацилглицероллипаза – в мембране липидных сферосом, моноацилглицероллипаза — в мембранах глиоксисом прорастающих семян. Указанные липазы различаются не только по специфичности, но и по оптимуму рН и температуре действия. Мобилизация депонированных жиров стимулируется глюкагоном и адреналином и, в меньшей степени, некоторыми другими гормонами (соматотропным, кортизолом). В постабсорбтивный период и при голодании глюкагон, действуя на адипоциты через аденилатциклазную систему, активирует протеинкиназу А, которая фосфо-рилирует и, таким образом, активирует гормончувствительную липазу, что инициирует липо-лиз и выделение жирных кислот и глицерина в кровь. При физической активности увеличивается секреция адреналина, который действует через β-адренергические рецепторы адипоцитов, активирующие аденилатциклазную систему (рис. 8-24). В настоящее время обнаружено 3 типа β-рецепторов: β1, β2, β3, активация которых приводит к липолитическому действию. К наибольшему липолитическому действию приводит активация β3-рецепторов. Адреналин одновременно действует и на α2-рецепторы адипоцитов, связанные с ингибирующим G-белком, что инактивирует аденилатциклазную систему. Вероятно, действие адреналина двояко: при низких концентрациях в крови преобладает его антилиполитическое действие через α2-рецепторы, а при высокой — преобладает липолитическое действие через β-рецепторы. Для мышц, сердца, почек, печени при голодании или физической работе жирные кислоты становятся важным источником энергии. Печень перерабатывает часть жирных кислот в кетоновые тела, используемые мозгом, нервной тканью и некоторыми другими тканями как источники энергии. В результате мобилизации жиров концентрация жирных кислот в крови увеличивается приблизительно в 2 раза, однако абсолютная концентрация жирных кислот в крови невелика даже в этот период. Т1/2 жирных кислот в крови тоже очень мал (менее 5 мин), что означает существование быстрого потока жирных кислот из жировой ткани к другим органам. Когда постабсорбтивный период сменяется аборбтивным, инсулин активирует специфическую фосфатазу, которая дефосфорилирует гормончувствительную липазу, и распад жиров останавливается.
Тут вы можете оставить комментарий к выбранному абзацу или сообщить об ошибке.
20 почему жировая ткань мало использует глицерол для синтеза триацилглицеролов
а) Синтез триглицеридов из углеводов. Если когда-нибудь большие количества углеводов попадают в организм, они либо сразу используются для получения энергии, либо запасаются в виде гликогена, а избыток их быстро превращается в триглицериды и хранится в таком виде в жировой ткани. У человека большая часть триглицеридов образуется в печени, но очень небольшие количества могут образовываться и в самой жировой ткани. Триглицериды, образуемые в печени, транспортируются главным образом в виде липопротеинов очень низкой плотности в жировую ткань, где и хранятся.
б) Превращение ацетил-КоА в жирные кислоты. Первым этапом синтеза триглицеридов является превращение углеводов в ацетил-КоА.
Это происходит во время обычного расщепления глюкозы гликолитической системой. Вследствие того, что жирные кислоты являются крупными полимерами уксусной кислоты, легко представить, каким образом ацетил-КоА может быть превращен в жирную кислоту. Однако синтез жирных кислот не обеспечивается просто изменением направления реакции окислительного расщепления. Этот синтез осуществляется двуступенчатым процессом, показанным на рисунке ниже, с использованием малонил-КоА и НАДФ-Н в качестве основных посредников процесса полимеризации.
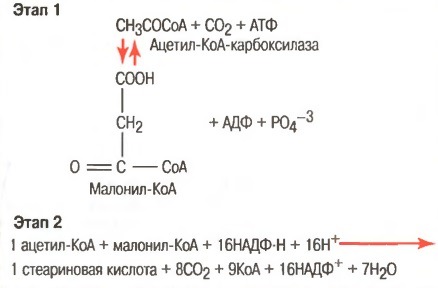
Синтез жирных кислот
в) Объединение жирных кислот с α-глицерофосфатом при образовании триглицеридов. Как только синтезируемые цепочки жирных кислот начинают включать от 14 до 18 атомов углерода, они взаимодействуют с глицеролом, образуя триглицериды. Ферменты, катализирующие эту реакцию, высокоспецифичны для жирных кислот с длиной цепочки от 14 атомов углерода и выше, что является фактором, контролирующим структурное соответствие триглицеридов, хранящихся в организме.
Читайте также: Распределения гормонов по тканям тела
Как показано на рисунке ниже, образование глицероловой части молекулы триглицерида обеспечивается α-глицерофосфатом, который является побочным продуктом реакции гликолитического расщепления глюкозы.
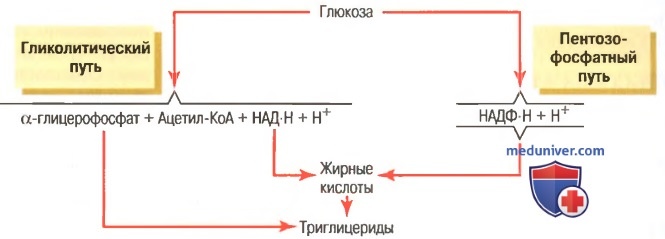
Общая схема синтеза триглицеридов из глюкозы
г) Эффективность превращения углеводов в жиры. Во время синтеза триглицеридов только 15% потенциально содержащейся в глюкозе энергии теряется в виде тепла. Остальные 85% преобразуются в энергию запасаемых триглицеридов.
д) Важность синтеза и хранения жиров. Синтез жиров из углеводов особенно важен в связи с двумя обстоятельствами.
1. Способность различных клеток организма запасать углеводы в виде гликогена выражена слабо. Только несколько сотен граммов гликогена может запасаться в печени, скелетных мышцах и всех других тканях организма, вместе взятых. В то же время могут запасаться килограммы жира, поэтому синтез жиров является способом, с помощью которого энергия, содержащаяся в избыточном количестве поступивших в организм углеводов (и белков), может запасаться, чтобы использоваться позднее. Количество энергии, которую запасает организм человека в виде жиров, приблизительно в 150 раз превышает количество энергии, запасаемой в виде углеводов.
2. Каждый грамм жиров содержит почти в 2,5 раза больше энергии, чем каждый грамм углеводов. Следовательно, при одном и той же массе тела организм может запасать в несколько раз больше энергии в виде жиров, чем в виде углеводов, что особенно важно, если необходима высокая степень подвижности, чтобы выжить.
е) Снижение синтеза жиров из углеводов при отсутствии инсулина. При отсутствии инсулина, как это бывает при тяжелом сахарном диабете, жиров синтезируется мало, если они вообще синтезируются, по следующим причинам. Во-первых, при отсутствии инсулина глюкоза не может попадать в сколько-нибудь существенных количествах в жировые ткани и клетки печени, что не обеспечивает образования достаточных количеств ацетил-КоА и НАДФ-Н, необходимых для синтеза жиров и получаемых при метаболизме глюкозы. Во-вторых, отсутствие глюкозы в жировых клетках существенно снижает количество наличного глицерофосфата, что также затрудняет образование триглицеридов.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
IV. Обмен триацилглицеролов
Приём пищи человеком происходит иногда со значительными интервалами, поэтому в организме выработались механизмы депонирования источников энергии. Жиры — наиболее выгодная и основная форма депонирования энергии. Запасы гликогена в организме не превышают 300 г и обеспечивают организм энергией не более суток. Депонированный жир может обеспечивать организм энергией при голодании в течение длительного времени (до 7-8 нед). Синтез жиров активируется в абсорбтивный период и происходит в основном в жировой ткани и печени. Но если жировая ткань — место депонирования жира, то печень выполняет важную роль превращения части углеводов, поступающих с пищей, в жиры, которые затем секретируются в кровь в составе ЛПОНП и доставляются в другие ткани (в первую очередь, в жировую). Синтез жиров в печени и жировой ткани стимулируется инсулином. Мобилизация жира активируется в тех случаях, когда глюкозы недостаточно для обеспечения энергетических потребностей организма: в постабсорбтивный период, при голодании и физической работе под действием гормонов глюкагона, адреналина, соматотропина. Жирные кислоты поступают в кровь и используются тканями как источники энергии.
А. Синтез жиров в жировой ткани и печени
Синтез жиров происходит в абсорбтивный период в печени и жировой ткани. Непосредственными субстратами в синтезе жиров являются ацил-КоА и глицерол-3-фосфат. Метаболический путь синтеза жиров в печени и жировой ткани одинаков, за исключением разных путей образования глицерол-3-фосфата.
Синтез жиров в печени и жировой ткани идёт через образование промежуточного продукта — фосфатидной кислоты (рис. 8-21).
Предшественник фосфатидной кислоты — глицерол-3-фосфат, образующийся в печени двумя путями:
восстановлением дигидроксиацетонфосфата — промежуточного метаболита гликолиза;
фосфорилированием глицеролкиназой свободного глицерола, поступающего в печень из крови (продукт действия ЛП-липазы на жиры ХМ и ЛПОНП).
В жировой ткани глицеролкиназа отсутствует, и восстановление дигидроксиацетонфосфата — единственный путь образования глицерол-3-фосфата. Следовательно, синтез жиров в жировой ткани может происходить только в абсорбтивный период, когда глюкоза поступает в адипоциты с помощью белка-переносчика глюкозы ГЛЮТ-4, активного только в присутствии инсулина, и распадается по пути гликолиза.
Читайте также: Баллистическая ткань тсвм изготавливается из чего
Синтез жиров в жировой ткани
В жировой ткани для синтеза жиров используются в основном жирные кислоты, освободившиеся при гидролизе жиров ХМ и ЛПОНП (рис. 8-22). Жирные кислоты поступают в адипоциты, превращаются в производные КоА и взаимодействуют с глицерол-3-фосфатом, образуя сначала лизофосфатидную кислоту, а затем фосфатидную. Фосфатидная кислота после дефосфорилирования превращается в диацилглицерол, который ацилируется с образованием триацилглицерола.
Кроме жирных кислот, поступающих в адипоциты из крови, в этих клетках идёт и синтез жирных кислот из продуктов распада глюкозы. В адипоцитах для обеспечения реакций синтеза жира распад глюкозы идёт по двум путям: гликолиз, обеспечивающий образование глицерол-3-фосфата и ацетил-КоА, и пентозофосфатный путь, окислительные реакции которого обеспечивают образование NADPH, служащего донором водорода в реакциях синтеза жирных кислот.
Молекулы жиров в адипоцитах объединяются в крупные жировые капли, не содержащие воды, и поэтому являются наиболее компактной формой хранения топливных молекул. Подсчитано, что, если бы энергия, запасаемая в жирах, хранилась в форме сильно гидратированных молекул гликогена, то масса тела человека увеличилась бы на 14-15 кг.

Рис. 8-21. Синтез жиров в печени и жировой ткани.
Синтез ТАГ в печени. Образование ЛПОНП в печени и транспорт жиров в другие ткани
Печень — основной орган, где идёт синтез жирных кислот из продуктов гликолиза. В гладком ЭР гепатоцитов жирные кислоты активируются и сразу же используются для синтеза жиров, взаимодействуя с глицерол-3-фосфатом. Как и в жировой ткани, синтез жиров идёт через образование фосфатидной кислоты. Синтезированные в печени жиры упаковываются в ЛПОНП и сек-ретируются в кровь (рис. 8-23).
В состав ЛПОНП, кроме жиров, входят холестерол, фосфолипиды и белок — апоВ-100. Это очень «длинный» белок, содержащий 11 536 аминокислот. Одна молекула апоВ-100 покрывает поверхность всего липопротеина.
ЛПОНП из печени секретируются в кровь (рис. 8-23), где на них, как и на ХМ, действует ЛП-липаза. Жирные кислоты поступают в ткани, в частности в адипоциты, и используются для синтеза жиров. В процессе удаления жиров из ЛПОНП под действием ЛП-липазы ЛПОНП сначала превращаются в ЛГШП, а затем в ЛПНП. В ЛПНП основными липидными компонентами служат холестерол и его эфиры, поэтому ЛПНП являются липопротеинами, доставляющими холестерол в периферические ткани. Глицерол, освободившийся из липопротеинов, кровью транспортируется в печень, где опять может использоваться для синтеза жиров.
Скорость синтеза жирных кислот и жиров в печени существенно зависит от состава пищи. Если в пище содержится более 10% жиров, то скорость синтеза жиров в печени резко снижается.
Б. Мобилизация жиров из жировой ткани
Адипоциты (место депонирования жиров) располагаются в основном под кожей, образуя подкожный жировой слой, и в брюшной полости, образуя большой и.малый сальники. Мобилизация жиров, т.е. гидролиз до глицерола и жирных кислот, происходит в постабсорбтивный период, при голодании и активной физической работе. Гидролиз внутриклеточного жира осуществляется под действием фермента гормончувствительной липазы — ТАГ-липазы. Этот фермент отщепляет одну жирную кислоту у первого углеродного атома глицерола с образованием диацилглицерола, а затем другие липазы гидролизуют его до глицерола и жирных кислот, которые поступают в кровь. Глицерол как водорастворимое вещество транспортируется кровью в свободном виде, а жирные кислоты (гидрофобные молекулы) в комплексе с белком плазмы — альбумином.
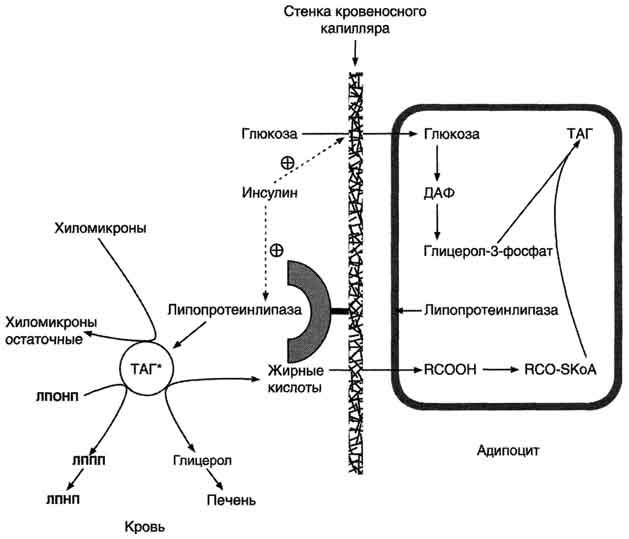
Рис. 8-22. Депонирование жира в адипоцитах в абсорбтивном периоде. После еды при повышении концентрации глюкозы в крови увеличивается секреция инсулина. Инсулин активирует транспорт глюкозы внутрь адипоцитов, действуя на ГЛЮТ-4, и синтез ЛП-липазы в адипоцитах и её экспонирование на поверхности стенки капилляров. ЛП-липаза, связанная с эндотелием сосудов, гидролизует жиры в составе ХМ и ЛПОНП. АпоС-II на поверхности ХМ и ЛПОНП активирует ЛП-липазу. Жирные кислоты проникают в адипоцит, а глицерол транспортируется в печень. Так как в адипоцитах нет фермента глицеролкиназы, то свободный глицерол не может использоваться для синтеза ТАГ в этой ткани. Активированные жирные кислоты взаимодействуют с глицерол-3-фосфатом, образующимся из дигидроксиацетонфосфата, и через фосфатидную кислоту превращаются в ТАГ, которые депонируются в адипоцитах. Сокращения: ТАГ* — триацилглицеролы в составе ХМ и ЛПОНП; ДАФ — дигидроксиацетонфосфат.
Читайте также: Ткань флис его свойства
В. Гормональная регуляция синтеза и мобилизации жиров
Какой процесс будет преобладать в организме — синтез жиров (липогенез) или их распад (липолиз), зависит от поступления пищи и физической активности. В абсорбтивном состоянии под действием инсулина происходит липогенез, в постабсорбтивном состоянии — липолиз, активируемый глюкагоном. Адреналин, секреция которого увеличивается при физической активности, также стимулирует липолиз.
Регуляция синтеза жиров. В абсорбтивный период при увеличении соотношения инсулин/

Рис. 8-23. Синтез и секреция ЛПОНП в печени. Белки, синтезированные в шероховатом ЭР (1), в аппарате Гольджи (2), формируют комплекс с ТАГ, называемый ЛПОНП, ЛПОНП комплектуются в секреторных гранулах (3), транспортируются к клеточной мембране и секретируются в кровь.
глюкагон в печени активируется синтез жиров. В жировой ткани индуцируется синтез ЛП-липазы в адипоцитах и осуществляется её экспонирование на поверхность эндотелия; следовательно, в этот период увеличивается поступление жирных кислот в адипоциты. Одновременно инсулин активирует белки-переносчики глюкозы — ГЛЮТ-4. Поступление глюкозы в адипоциты и гликолиз также активируются. В результате образуются все необходимые компоненты для синтеза жиров: глицерол-3-фосфат и активные формы жирных кислот. В печени инсулин, действуя через различные механизмы, активирует ферменты путём дефосфорилирования и индуцирует их синтез. В результате увеличиваются активность и синтез ферментов, участвующих
в превращении части глюкозы, поступающей с пищей, в жиры. Это — регуляторные ферменты гликолиза, пируватдегидрогеназный комплекс и ферменты, участвующие в синтезе жирных кислот из ацетил-КоА. Результат действия инсулина на обмен углеводов и жиров в печени — увеличение синтеза жиров и секреция их в кровь в составе ЛПОНП. ЛПОНП доставляют жиры в капилляры жировой ткани, где действие ЛП-липазы обеспечивает быстрое поступление жирных кислот в адипоциты, где они депонируются в составе триацилглицеринов.
Запасание жиров в жировой ткани — основная форма депонирования источников энергии в организме человека (табл. 8-6). Запасы жиров в организме человека массой 70 кг составляют 10 кг, но у многих людей количество жиров может быть значительно больше.
Жиры образуют в адипоцитах жировые вакуоли. Жировые вакуоли иногда заполняют значительную часть цитоплазмы. Скорость синтеза и мобилизации подкожного жира происходит неравномерно в разных частях организма, что связано с неодинаковым распределением рецепторов гормонов на адипоцитах.
Регуляция мобилизации жиров. Мобилизация депонированных жиров стимулируется глюкагоном и адреналином и, в меньшей степени, некоторыми другими гормонами (соматотроп-ным, кортизолом). В постабсорбтивный период и при голодании глюкагон, действуя на адипоциты через аденилатциклазную систему, активирует протеинкиназу А, которая фосфо-рилирует и, таким образом, активирует гормончувствительную липазу, что инициирует липо-лиз и выделение жирных кислот и глицерина в кровь. При физической активности увеличивается секреция адреналина, который действует через β-адренергические рецепторы адипоцитов, активирующие аденилатциклазную систему (рис. 8-24). В настоящее время обнаружено 3 типа β-рецепторов: β1, β2, β3, активация которых приводит к липолитическому действию. К наибольшему липолитическому действию приводит активация β3-рецепторов. Адреналин одновременно действует и на α2-рецепторы адипоцитов, связанные с ингибирующим G-белком, что инактивирует аденилатциклазную систему. Вероятно, действие адреналина двояко: при низких концентрациях в крови преобладает его антилиполитическое действие через α2-рецепторы, а при высокой — преобладает липолитическое действие через β-рецепторы.
Для мышц, сердца, почек, печени при голодании или физической работе жирные кислоты становятся важным источником энергии. Печень перерабатывает часть жирных кислот в кетоновые тела, используемые мозгом, нервной тканью и некоторыми другими тканями как источники энергии.
В результате мобилизации жиров концентрация жирных кислот в крови увеличивается приблизительно в 2 раза (рис. 8-25), однако абсолютная концентрация жирных кислот в крови невелика даже в этот период. Т1/2 жирных кислот в крови тоже очень мал (менее 5 мин), что означает существование быстрого потока жирных кислот из жировой ткани к другим органам. Когда постабсорбтивный период сменяется аборбтивным, инсулин активирует специфическую фосфатазу, которая дефосфорилирует гормончувствительную липазу, и распад жиров останавливается.
Г. Нарушения жирового обмена. Oжирение
Жировая ткань составляет 20-25% от общей массы тела у женщин и 15-20% у мужчин. Однако избыточное накопление жира в адипоцитах
Таблица 8-6. Запасы энергии в организме человека (масса 70 кг)