
34. Аэробный дихотомический распад глюкозы в тканях, его основные этапы. Биологическое значение. Пентозофосфатный путь распада глюкозы, его биологическая роль.
ПРОЦЕСС АЭРОБНОГО ОКИСЛЕНИЯ ГЛЮКОЗЫ разделяют на 3 этапа:
1. Расщепление глюкозы до пирувата.
2. Окислительное декарбоксилирование пирувата до ацетил-КоА.
3. Окисление ацетила в цикле Кребса (ЦТК).
Эти этапы можно представить в виде общей схемы:
1 Этап. Расщепление глюкозы до пирувата.
По современным представлениям первый этап окисления глюкозы протекает в цитозоле и катализируется надмолекулярным белковым комплексом — гликолитическим метаболоном, включающим в себя до десятка отдельных ферментов.Контроль направления потока метаболитов по данному метаболическому пути осуществляется с помощью термодинамических механизмов. Имеется три реакции, в ходе которых теряется большое количество энергии: гексокиназная, фосфофруктокиназная и пируваткиназная, эти реакции в клетке практически необратимы, и за счет их необратимости процесс становится необратимым.
2 этап. Окислительное декарбоксилирование пирувата с образованием ацетил-КоА. Это превращение катализируется надмолекулярным пируватдегидрогеназным комплексом, локализованным в матриксе митохондрий. Суммарное уравнение процесса:
2Пируват +2НАД + +2HS-КоА –– 2Ацетил-КоА +2НАДН+Н + +2СО2
В ходе окисления 2 моль пирувата высвобождается около 500 кДж энергии, из них около 420 кДж накапливается в виде энергии восстановленного НАД. Остальная энергия рассеивается в виде теплоты.
активность комплекса снижается, если клетка хорошо обеспечена энергией (много АТФ и НАДН+Н + ) или же цикл Кребса не справляется с окислением имеющегося ацетил-КоА.
Образовавшийся ацетил-КоА, как уже неоднократно упоминалось, поступает в цикл трикарбоных кислот, работа которого сопряжена с функционированием цепи дыхательных ферментов. При функционировании этих двух метаболических путей остаток ацетила окисляется до углекислого газа и воды.
Суммарное уравнение для всех трех этапов окисления молекулы глюкозы выглядит следующим образом:Глюкоза + 2 АДФ + 2 ГДФ + 4Ф + 10 НАД + + 2 ФАД +2Н2О –– 6 СО2 + 2 АТФ + 2 ГТФ + 10 НАДН+Н + + 2 ФАДН2; при окислении 1 молекулы глюкозы до углекислого газа и воды клетка получит 38 молекул АТФ (40 синтезируется и 2 расходуется).Второй важной функцией аэробного окисления глюкозы является пластическая функция. Из промежуточных продуктов ее окисления синтезируется много различных соединений, необходимых клетке: Гл-6-ф используется в клетке для синтеза пентоз и глюкуроновой кислоты;Фр-6-ф — для синтеза аминосахаров; ФГА и ФДА — для образования 3-фосфоглицерола, необходимого для синтеза глицеролсодержащих липидов; 3-фосфоглицериновая кислота — для синтеза заменимых аминокислот: серина, глицина и цистеина; ФЭП — для синтеза сиаловых кислот, используемых при синтезе гетероолигосахаридов; пируват — для синтеза аланина; ацетил-КоА — для синтеза жирных кислот и стероидов.
В пентозном цикле окисления углеводов идет образование восстановленного НАДФН+Н + и целого ряда моносахаридов, имеющих в своем составе пять атомов углерода — пентоз (рибоза, ксилоза).
Восстановительные эквиваленты и пентозы необходимы для биосинтетических процессов, протекающих в клетках.
Пентозный путь окисления углеводов может быть разделен на два этапа включает в себя достаточно много отдельных парциальных реакций:
Суммарное уравнение окислительного этапа пентозного цикла окисления :
В ходе неокислительного этапа цикла в результате изомеризации образуются необходимые для клетки фосфорилированные пентозы: рибозо-5-фосфат и ксилулозо-5-фосфат.Кроме того, важно отметить, что на этом этапе образуются промежуточные продукты, идентичные с промежуточными продуктами первого этапа аэробного окисления глюкозы: 3-фосфоглицериновый альдедид и Фруктозо-6-фосфат. За счет этих общих промежуточных соединений создается возможность переключения потока метаболитов с пентозного цикла окисления на путь аэробного (или анаэробного) окисления глюкозы и наоборот.
За шесть оборотов пентозного цикла окисления полностью сгорает один остаток глюкозы, так что суммарное уравнение окисления глюкозы в цикле, начиная с Гл-6-ф, можно представить в следующем виде: Гл-6-ф + 7 Н2О + 12 НАДФ + –– 6 СО2 + Ф + 12 НАДФН+Н +
Читайте также: Оригами из ткани рисунки
Пентозофосфатный цикл активно функционирует в печени, жировой ткани, коре надпочечников, семенниках и в молочной железе в период лактации. В этих тканях активно идут процессы синтеза высших жирных кислот, аминокислот или стероидов, нуждающиеся в восстановительных эквивалентах в виде НАДФН+Н + . Цикл интенсивно работает также в эритроцитах, в которых НАДФН+Н + используется для подавления перекисного окисления мембранных липидов.
Распад глюкозы в аэробных и анаэробных условиях
Катаболизм глюкозы – основной поставщик энергии для большинства клеток организма.
Аэробный распад протекает в условиях достаточного снабжения тканей кислородом. Он включает несколько стадий:
1. Аэробный распад глюкозы до двух молекул пирувата в гиалоплазме клеток,который можно разделить на 2 этапа:
а)подготовительный этап, в ходе которого глюкоза фосфорилируется и расщепляется на две молекулы фосфотриоз.
б) основной этап, сопряженный с синтезом АТФ. В результате серии реакций фосфотриозы превращаются в пируват.
Превращение пирувата в ацетил-КоА и окисление последнего в цикле Кребса
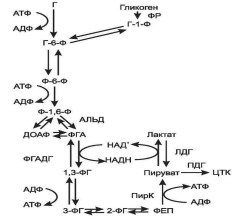
Глюкозо-6-фосфат, образованный в результате фосфорилирования глюкозы с учас-тием АТФ, в ходе следующей реакции превращается в фруктозо-6-фосфат. Это обратимая реакция изомеризации протекает при участии глюкозофосфатизомеразы. Затем следует еще одна реакция фосфорилирования за счет АТФ, катализирует ее фосфофруктокиназа. В ходе этой реакции фруктозо-6-фосфат превращается в фруктозо-1,6-бисфосфат. Этот метаболит далее расщепляется на 2 триозофосфата: фосфоглицериноый альдегид (ФГА) и диоксиацетонфосфат (ДОАФ), реакцию катализирует альдолаза. В последующих реакциях гликолиза используется только ФГА, поэтому ДОАФ превращается с участием фермента триозофосфатизомеразы в ФГА. На этом подготовительный этап заканчивается.
Основной этап аэробного распада включает реакции, связанные с синтезом АТФ. Сна-чала происходит окисление ФГАпри участии ФГА-дегидрогеназы. Этот фермент является НАД + -зависимымым. Восстановленный НАД + затем окисляется в дыхательной цепи (перенос НАДН в митохондрии, где находится дыхательная цепь, происходит при участии челночных механизмов, это связано с тем, что митохондриальная мембрана непроницаема для НАДН). Кроме того, свободная реакция окисления этой реакции концентрируется в макроэргической связи продукта реакции. Продуктом реакции является 1,3-бисфосфоглицерат. Для фосфорилирования используется Н3РО4. В следующей реакции высокоэнергетический фосфат передается на АДФ с образованием АТФ. Катализирует реакцию фосфоглицераткиназа. АТФ в данной реакции образуется путем субстратного фосфорилирования. Продуктом реакции является 3-фосфоглицерат. В следующей реакции происходит перенос фосфата из 3 положения во второе с образованием 2-фосфоглицерата. Из него при отщеплении воды образуется фосфоенолпируват – макроэргическое соединение, фосфатная группа которого в следующей реакции переносится на АДФ с образованием АТФ при участии пируваткиназы (это вторая реакция субстратного фосфорилирования). Продуктом реакции является пируват.
Во второй стадии пируват превращается в ацетил-КоА, который сгорает в цикле Кребса и дыхательной цепи (третья стадия) до СО2 и Н2О.
Всего при аэробном окислении глюкозы выделяется 32 АТФ.
Анаэробный гликолиз представляет собой распад глюкозы в анаэробных условиях (условиях недостаточного снабжения кислородом), поэтому он не зависит от работы митохондриальной дыхательной цепи. АТФ в гликолизе образуется путем субстратногофосфорилирования. Конечным продуктом является лактат (рис. 5).
Все реакции анаэробного гликолиза (11 реакций) протекают в гиалоплазме. Первые 10 реакций до пирувата идентичны аэробному распаду. В последней реакции происходит восстановление пирувата в лактат цитозольным НАДН. Реакцию эту катализирует лактатдегидрогеназа. С помощью этой реакции обеспечивается регенерация НАД + из НАДН без участия дыхательной цепи в ситуациях, связанных с недостаточным снабжением клеток кислородом. Роль акцептора водорода от НАДН (подобно кислороду в дыхательной цепи) выполняет пируват. Таким образом, значение реакции восстановления пирувата заключается не в образовании лактата, а в том, что данная цитозольная реакция обеспечивает регенерацию НАД + . К тому же лактат не является конечным продуктом метаболизма, удаляемым из организма (лактат – тупиковый метаболит). Он выводится в кровь и утилизируется: либо превращается в глюкозу (75%), либо при доступности кислорода окисляется до СО2 и Н2О (25%).
Читайте также: Дай определение понятию ткань
Энергетический выход анаэробного гликолиза составляет 2 АТФ.
Недостатками анаэробного гликолиза являются: малый энергетический выход по сравнению с аэробным распадом и накопление лактата, что приводит к ацидозу, усталости.
Достоинства гликолиза: он полезен при выполнении кратковременной работы, является единственным источником энергии для эритроцитов (в них отсутствуют митохондрии), а также необходим в разных органах при дефиците кислорода (наложение жгута, падение давления, нарушение кровотока).
Регуляция.Регуляторными ферментами гликолиза являются: гексокиназа, фосфофруктокиназа и пируваткиназа. Аллостерическими ингибиторами являются АТФ и цитрат. Стимулирует гликолиз инсулин, а тормозят глюкагон, глюкокортикостероиды. Катехоламины тормозят гликолиз в печени и активируют в мышцах.
Некоторые ткани нуждаются в постоянном поступлении глюкозы. Когда поступление углеводов в составе пищи недостаточно, содержание глюкозы в крови некоторое время поддерживается в пределах нормы за счет расщепления гликогена в печени. Однако запасы гликогена в печени невелики, они практически полностью исчерпываются после суточного голодания. В этом случае в печени начинается глюконеогенез (ГНГ) – процесс синтеза глюкозы из веществ неуглеводной природы. Его основной функцией является поддержание уровня глюкозы в крови в период длительного голодания и интенсивных физических нагрузок. В первую очередь это важно для головного мозга (он не может обеспечивать потребности в энергии за счет окисления жирных кислот) и эритроцитов – в них аэробный путь распада веществ невозможен из-за отсутствия митохондрий. Процесс в основном (на 90%) протекает в печени и менее интенсивно в корковом веществе почек и слизистой тонкого кишечника.
Субстратами ГНГ являются лактат, аминокислоты, глицерин. Аминокислоты включаются в ГНГ при длительном голодании или продолжительной мышечной работе. Глицерин освобождается в процессе липолиза из жировой ткани в период голодания или при
длительной физической нагрузке. Лактат – продукт анаэробного гликолиза. Его источником в покое являются эритроциты, а при работе – мышцы. Таким образом, лактат используется в глюконеогенезе постоянно. Большинство реакций глюконеогенеза протекает за счет обратимых реакций гликолиза и катализируется теми же ферментами. Однако три реакции гликолиза необратимы. На этих стадиях реакции ГНГ протекают другими путями (обходные пути ГНГ).
Первая необратимая реакция – образование из пирувата фосфоенолпирувата . В ГНГ это происходит в ходе двух реакций. Пируват в митохондриях карбоксилируется с образованием оксалоацетата при участии пируваткарбоксилазы – биотинзависимого фермента. Реакция протекает с использованием энергии АТФ. Дальнейшие превращения оксалоацетата протекают в цитозоле. Фосфоенолпируваткарбоксикиназа с затратой энергии ГТФ превращает оксалоацетат в фосфоенолпируват. Дальнейшие реакции ГНГ вплоть до образования фруктозо-1,6-бисфосфата протекают в цитозоле и катализируются гликолитическими ферментами. Затем следует еще одна необратимая реакция ГНГ, катализируемая фруктозо-1,6-бисфосфатазой, в которой происходит отщепление остатка фосфорной кислоты гидролитическим путем. Образующийся при этом фруктозо-6-фосфат гликолитическим ферментом изомеризуется до глюкозо-6-фосфата, от которого отщепляется остаток фосфорной кислоты при участии еще одного необратимо работающего фермента – глюкозо-6-фосфатазы. Образовавшаяся свободная глюкоза из клетки выходит в кровь. В ходе ГНГ расходуется 6 моль АТФ на синтез 1 моль глюкозы из пирувата или лактата.
Ключевыми ферментами ГНГ, катализирующими необратимые обходные реакции, являются: пируваткарбоксилаза, фосфоенолпируваткарбоксикиназа, фруктозо-1,6-бисфосфатаза и глюкозо-6-фосфатаза. Активируют ГНГ аллостерически АТФ и цитрат, обязательным активатором первого фермента является ацетил-КоА. ГНГ усиливают гормоны: глюкагон, глюкокортикостероиды. Тормозит ГНГ гормон покоя и сытости – инсулин.
Читайте также: Шуба крытая тканью как называется
Лактат, образовавшийся в ходе анаэробного гликолиза, не является конечным продуктом метаболизма. Использование лактата связано с его превращением в печени в пируват и с дальнейшим использованием последнего. Лактат, поступивший из интенсивно работающих мышц, выходит в кровь, а затем поступает в печень. В печени в лактатдегидрогеназной реакции лактат превращается в пируват, который включается в ГНГ, а образовавшаяся глюкоза поступает в кровь и поглощается скелетными мышцами. Эту последовательность событий называют «глюкозо-лактатным циклом», или «циклом Кори». Цикл Кори выполняет две важнейшие функции: обеспечивает утилизацию лактата и предотвращает лактатацидоз. Часть пирувата, образовавшегося из лактата (25%), окисляется печенью до СО2 и Н2О с освобождением энергии.
57. Аэробный гликолиз. Последовательность реакций до образования пирувата (аэробный гликолиз). Использование глюкозы для синтеза жиров. Энергетический эффект аэробного распада глюкозы.
Окисление глюкозы до СО2 и Н2О (аэробный распад). Аэробный распад глюкозы можно выразить суммарным уравнением:
Этот процесс включает несколько стадий (рис. 7-33).
Аэробный гликолиз — процесс окисления глюкозы с образованием двух молекул пирувата;
Общий путь катаболизма, включающий превращение пирувата в ацетил-КоА и его дальнейшее окисление в цитратом цикле;
ЦПЭ на кислород, сопряжённая с реакциями дегидрирования, происходящими в процессе распада глюкозы.
В определённых ситуациях обеспечение кислородом тканей может не соответствовать их потребностям. Например, на начальных стадиях интенсивной мышечной работы при стрессе сердечные сокращения могут не достигать нужной частоты, а потребности мышц в кислороде для аэробного распада глюкозы велики. В подобных случаях включается процесс, который протекает без кислорода и заканчивается образованием лактата из пировиноградной кислоты. Этот процесс называют анаэробным распадом, или анаэробным гликолизом. Анаэробный распад глюкозы энергетически малоэффективен, но именно этот процесс может стать единственным источником энергии для мышечной клетки в описанной ситуации. В даньнейшем, когда снабжение мышц кислородом будет достаточным в результате перехода сердца на ускоренный ритм, анаэробный распад переключается на аэробный. Пути катаболизма глюкозы и их энергетический эффект показаны на рис. 7-34.
Аэробным гликолизом называют процесс окисления глюкозы до пировиноградной кислоты, протекающий в присутствии кислорода. Все ферменты, катализирующие реакции этого процесса, локализованы в цитозоле клетки.
1. Этапы аэробного гликолиза
В аэробном гликолизе можно выделить 2 этапа.
Подготовительный этап, в ходе которого глюкоза фосфорилируется и расщепляется на две молекулы фосфотриоз. Эта серия реакций протекает с использованием 2 молекул АТФ.
Этап, сопряжённый с синтезом АТФ. В результате этой серии реакций фосфотриозы превращаются в пируват. Энергия, высвобождающаяся на этом этапе, используется для синтеза 10 моль АТФ.
Выход АТФ при аэробном гликолизе
На образование фруктозо-1,6-бисфосфата из одной молекулы глюкозы требуется 2 молекулы АТФ (реакции 1 и 3 на рис. 7-33). Реакции, связанные с синтезом АТФ, происходят после распада глюкозы на 2 молекулы фосфотриозы, т.е. на втором этапе гликолиза. На этом этапе происходят 2 реакции субстратного фосфорилирования и синтезируются 2 молекулы АТФ (реакции 7 и 10). Кроме того, одна молекула глицеральдегид-3-фосфата дегидрируется (реакция 6), a NADH передаёт водород в митохондриальную ЦПЭ, где синтезируется 3 молекулы АТФ путём окислительного фосфорилирования. В данном случае количество АТФ (3 или 2) зависит от типа челночной системы. Следовательно, окисление до пирувата одной молекулы глицеральдегид-3-фосфата сопряжено с синтезом 5 молекул АТФ. Учитывая, что из глюкозы образуются 2 молекулы фосфотриозы, полученную величину нужно умножить на 2 и затем вычесть 2 молекулы АТФ, затраченные на первом этапе. Таким образом, выход АТФ при аэробном гликолизе составляет (5×2) — 2 = 8 АТФ.
Выход АТФ при аэробном распаде глюкозы до конечных продуктов
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом