
Кость, os, ossis, как орган живого организма состоит из нескольких тканей, главнейшей из которых является костная.
Химический состав кости и ее физические свойства.
Костное вещество состоит из двоякого рода химических веществ: органических (1/3), главным образом оссеина, и неорганических (2/3), главным образом солей кальция, особенно фосфорнокислой извести (более половины — 51,04 %). Если кость подвергнуть действию раствора кислот (соляной, азотной и др.), то соли извести растворяются (decalcinatio), а органическое вещество остается и сохраняет форму кости, будучи, однако, мягким и эластичным. Если же кость подвергнуть обжиганию, то органическое вещество сгорает, а неорганическое остается, также сохраняя форму кости и ее твердость, но будучи при этом весьма хрупким. Следовательно, эластичность кости зависит от оссеина, а твердость ее — от минеральных солей. Сочетание неорганических и органических веществ в живой кости и придает ей необычайные крепость и упругость. В этом убеждают и возрастные изменения кости. У маленьких детей, у которых оссеина сравнительно больше, кости отличаются большой гибкостью и потому редко ломаются. Наоборот, в старости, когда соотношение органических и неорганических веществ изменяется в пользу последних, кости становятся менее эластичными и более хрупкими, вследствие чего переломы костей чаще всего наблюдаются у стариков.
Строение кости
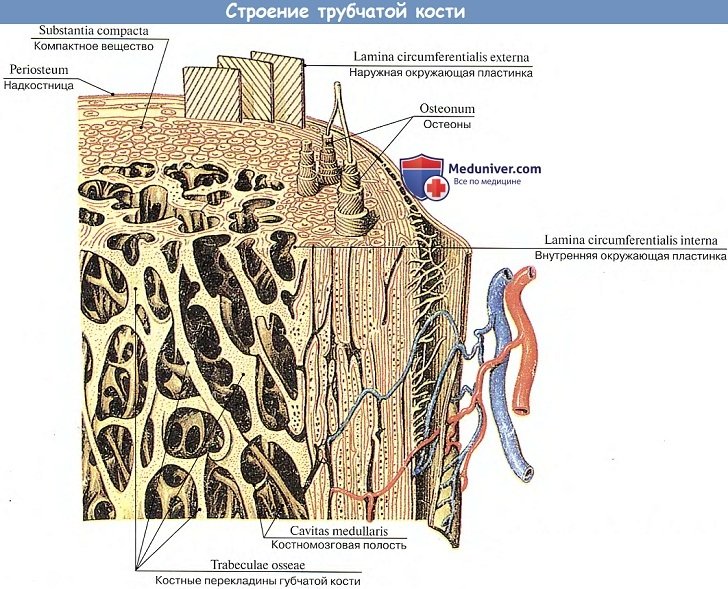
Структурной единицей кости, видимой в лупу или при малом увеличении микроскопа, является остеон, т. е. система костных пластинок, концентрически расположенных вокруг центрального канала, содержащего сосуды и нервы.
Остеоны не прилегают друг к другу вплотную, а промежутки между ними заполнены интерстициальными костными пластинками. Остеоны располагаются не беспорядочно, а соответственно функциональной нагрузке на кость: в трубчатых костях параллельно длиннику кости, в губчатых — перпендикулярно вертикальной оси, в плоских костях черепа — параллельно поверхности кости и радиально.
Вместе с интерстициальными пластинками остеоны образуют основной средний слой костного вещества, покрытый изнутри (со стороны эндоста) внутренним слоем костных пластинок, а снаружи (со стороны периоста) — наружным слоем окружающих пластинок. Последний пронизан кровеносными сосудами, идущими из надкостницы в костное вещество в особых прободающих каналах. Начало этих каналов видно на мацерирован-ной кости в виде многочисленных питательных отверстий (foramina nutricia). Проходящие в каналах кровеносные сосуды обеспечивают обмен веществ в кости. Из остеонов состоят более крупные элементы кости, видимые уже невооруженным глазом на распиле или на рентгенограмме, — перекладины костного вещества, или трабекулы. Из этих трабекул складывается двоякого рода костное вещество: если трабекулы лежат плотно, то получается плотное компактное вещество, substantia compacta. Если трабекулы лежат рыхло, образуя между собою костные ячейки наподобие губки, то получается губчатое, трабекулярное вещество, substantia spongiosa, trabecularis (spongia, греч. — губка).
Распределение компактного и губчатого вещества зависит от функциональных условий кости. Компактное вещество находится в тех костях и в тех частях их, которые выполняют преимущественно функцию опоры (стойки) и движения (рычаги), например в диафизах трубчатых костей.
В местах, где при большом объеме требуется сохранить легкость и вместе с тем прочность, образуется губчатое вещество, например в эпифизах трубчатых костей.
Перекладины губчатого вещества располагаются не беспорядочно, а закономерно, также соответственно функциональным условиям, в которых находится данная кость или ее часть. Поскольку кости испытывают двойное действие — давление и тягу мышц, постольку костные перекладины располагаются по линиям сил сжатия и растяжения. Соответственно разному направлению этих сил различные кости или даже части их имеют разное строение. В покровных костях свода черепа, выполняющих преимущественно функцию защиты, губчатое вещество имеет особый характер, отличающий его от остальных костей, несущих все 3 функции скелета. Это губчатое вещество называется диплоэ, diploe (двойной), так как оно состоит из неправильной формы костных ячеек, расположенных между двумя костными пластинками — наружной, lamina externa, и внутренней, lamina interna. Последнюю называют также стекловидной, lamina vftrea, так как она ломается при повреждениях черепа легче, чем наружная.
Костные ячейки содержат костный мозг — орган кроветворения и биологической защиты организма. Он участвует также в питании, развитии и росте кости. В трубчатых костях костный мозг находится также в канале этих костей, называемом поэтому костномозговой полостью, cavitas medullaris.
Таким образом, все внутренние пространства кости заполняются костным мозгом, составляющим неотъемлемую часть кости как органа.
Костный мозг бывает двух родов: красный и желтый.
Красный костный мозг, medulla ossium rubra (детали строения см. в курсе гистологии), имеет вид нежной красной массы, состоящей из ретикулярной ткани, в петлях которой находятся клеточные элементы, имеющие непосредственное отношение к кроветворению (стволовые клетки) и костеобразованию (костесозидатели — остеобласты и костеразруши-тели — остеокласты). Он пронизан нервами и кровеносными сосудами, питающими, кроме костного мозга, внутренние слои кости. Кровеносные сосуды и кровяные элементы и придают костному мозгу красный цвет.
Желтый костный мозг, medulla ossium flava, обязан своим цветом жировым клеткам, из которых он главным образом и состоит.
В периоде развития и роста организма, когда требуются большая кроветворная и костеобразующая функции, преобладает красный костный мозг (у плодов и новорожденных имеется только красный мозг). По мере роста ребенка красный мозг постепенно замещается желтым, который у взрослых полностью заполняет костномозговую полость трубчатых костей.
Снаружи кость, за исключением суставных поверхностей, покрыта надкостницей, periosteum (периост).
Надкостница — это тонкая, крепкая соединительнотканная пленка бледно-розового цвета, окружающая кость снаружи и прикрепленная к ней с помощью соединительнотканных пучков — прободающих волокон, проникающих в кость через особые канальцы. Она состоит из двух слоев: наружного волокнистого (фиброзного) и внутреннего костеобразующего (остеогенного, или камбиального). Она богата нервами и сосудами, благодаря чему участвует в питании и росте кости в толщину. Питание осуществляется за счет кровеносных сосудов, проникающих в большом числе из надкостницы в наружное компактное вещество кости через многочисленные питательные отверстия (foramina nutricia), а рост кости осуществляется за счет остеобластов, расположенных во внутреннем, прилегающем к кости слое (камбиальном). Суставные поверхности кости, свободные от надкостницы, покрывает суставной хрящ, cartilage articularis.
Таким образом, в понятие кости как органа входят костная ткань, образующая главную массу кости, а также костный мозг, надкостница, суставной хрящ и многочисленные нервы и сосуды.
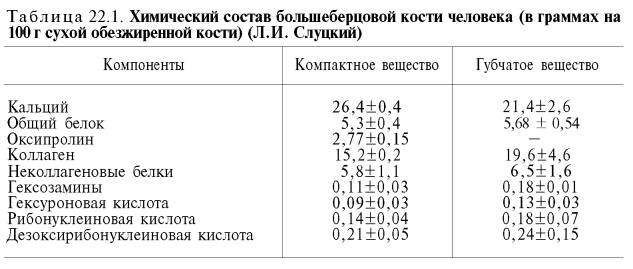
ХИМИЧЕСКИЙ СОСТАВ КОСТНОЙ ТКАНИ
Изучение химического состава костной ткани сопряжено со значительными трудностями, поскольку для выделения органического матрикса требуется провести деминерализацию кости. Кроме того, содержание и состав органического матрикса подвержены значительным изменениям в зависимости от степени минерализации костной ткани.
Известно, что при продолжительной обработке кости в разведенных растворах кислот ее минеральные компоненты растворяются и остается гибкий мягкий органический остаток (органический матрикс), сохраняющий форму интактной кости. Межклеточный органический матрикс компактной кости составляет около 20%, неорганические вещества – 70% и вода – 10%. В губчатой кости преобладают органические компоненты, которые составляют более 50%, на долю неорганических соединений приходится 33–40%. Количество воды сохраняется в тех же пределах, что и в компактной кости (Ю.С. Касавина, В.П. Торбенко).
По данным А. Уайта и соавт., неорганические компоненты составляют около 1 /4 объема кости; остальную часть занимает органический матрикс. Вследствие различий в относительной удельной массе органических и неорганических компонентов на долю нерастворимых минералов приходится половина массы кости.
Читайте также: Ткани из тканей компаньонов
Неорганический состав костной ткани. Более 100 лет назад было высказано предположение, что кристаллы костной ткани имеют структуру апатита. В дальнейшем это в значительной мере подтвердилось. Действительно, кристаллы кости относятся к гидроксилапатитам, имеют форму пластин или палочек и следующий химический состав – Са10(РО4)6(ОН)2. Кристаллы гидроксилапатита составляют лишь часть минеральной фазы костной ткани, другая часть представлена аморфным фосфатом кальция Са3(РО4)2. Содержание аморфного фосфата кальция подвержено значительным колебаниям в зависимости от возраста. Аморфный фосфат кальция преобладает в раннем возрасте, в зрелой кости преобладающим становится кристаллический гидроксилапатит. Обычно аморфный фосфат кальция рассматривают как лабильный резерв ионов Са 2+ и фосфата.
В организме взрослого человека содержится более 1 кг кальция, который почти целиком находится в костях и зубах, образуя вместе с фосфатом нерастворимый гидроксилапатит. Большая часть кальция в костях постоянно обновляется. Ежедневно кости скелета теряют и вновь восстанавливают примерно 700–800 мг кальция.
В состав минеральной фазы кости входит значительное количество ионов, которые обычно не содержатся в чистом гидроксилапатите, например ионы натрия, магния, калия, хлора и др. Высказано предположение, что в кристаллической решетке гидроксилапатита ионы Са 2+ могут замещаться другими двухвалентными катионами, тогда как анионы, отличные от фосфата и гидроксила, либо адсорбируются на поверхности кристаллов, либо растворяются в гидратной оболочке кристаллической решетки.
Органический матрикс костной ткани. Приблизительно 95% органического матрикса приходится на коллаген. Вместе с минеральными компонентами коллаген является главным фактором, определяющим механические свойства кости. Коллагеновые фибриллы костного матрикса образованы коллагеном типа 1. Известно, что данный тип коллагена входит также в состав сухожилий и кожи, однако коллаген костной ткани обладает некоторыми особенностями. Есть данные, что в коллагене костной ткани несколько больше оксипролина, чем в коллагене сухожилий и кожи. Для костного коллагена характерно большое содержание свободных ε-амино-групп лизиновых и оксилизиновых остатков. Еще одна особенность костного коллагена – повышенное по сравнению с коллагеном других тканей содержание фосфата. Большая часть этого фосфата связана с остатками серина.
В сухом деминерализованном костном матриксе содержится около 17% неколлагеновых белков, среди которых находятся и белковые компоненты протеогликанов. В целом количество протеогликанов в сформировавшейся плотной кости невелико.
В состав органического матрикса костной ткани входят гликозамино-гликаны, основным представителем которых является хондроитин-4-суль-фат. Хондроитин-6-сульфат, кератансульфат и гиалуроновая кислота содержатся в небольших количествах.
Принято считать, что гликозаминогликаны имеют непосредственное отношение к оссификации . Показано, что окостенение сопровождается изменением гликозаминогликанов: сульфатированные соединения уступают место несульфатированным. Костный матрикс содержит липиды, которые представляют собой непосредственный компонент костной ткани, а не являются примесью в результате недостаточно полного удаления богатого липидами костного мозга. Липиды принимают участие в процессе минерализации. Есть основания полагать, что липиды могут играть существенную роль в образовании ядер кристаллизации при минерализации кости.
Биохимические и цитохимические исследования показали, что остеобласты – основные клетки костной ткани – богаты РНК . Высокое содержание РНК в костных клетках отражает их активность и постоянную биосинтетическую функцию (табл. 22.1).
Своеобразной особенностью костного матрикса является высокая концентрация цитрата: около 90% его общего количества в организме приходится на долю костной ткани. Принято считать, что цитрат необходим для минерализации костной ткани. Вероятно, цитрат образует комплексные соединения с солями кальция и фосфора, обеспечивая возможность повышения концентрации их в ткани до такого уровня, при котором могут начаться кристаллизация и минерализация.
Кроме цитрата, в костной ткани обнаружены сукцинат, фумарат, малат, лактат и другие органические кислоты.
Аминокислоты и биосинтез белка
АМИНОКИСЛОТЫ БЕЛКОВ

БИОСИНТЕЗ БЕЛКА НА РИБОСОМЕ
Аминокислоты – (аминокарбоновые кислоты; амк) — органические соединения , в молекуле которых одновременно содержатся карбоксильные и аминные группы (аминогруппы). Т.е. а минокислоты могут рассматриваться , как производные карбоновых кислот, в которых один или несколько атомов водорода заменены на аминогруппы.
- Карбоксильная группа (карбоксил) -СООН — функциональная одновалентная группа, входящая в состав карбоновых кислот и определяющая их кислотные свойства.
- Аминогруппа — функциональная химическая одновалентная группа -NH2, органический радикал, содержащий один атом азота и два атома водорода.
Известно более 200 природных аминокислот, которые можно классифицировать по-разному. Структурная классификация исходит из положения функциональных групп на альфа-, бета-, гамма- или дельта- положении аминокислоты.
Кроме этой классификации, существуют еще и другие, например, классификация по полярности, рН уровню, а также типу группы боковой цепи (алифатические, ациклические, ароматические аминокислоты, аминокислоты, содержащие гидроксил или серу, и т.д.).
В виде белков аминокислоты являются вторым (после воды) компонентом мышц, клеток и других тканей человеческого организма. Аминокислоты играют решающую роль в таких процессах, как транспорт нейротрансмиттеров и биосинтезе.
Общая структура аминокислот. Альфа аминокислоты. Изомеризация аминокислот.
Аминокислоты – биологически важные органические соединения, состоящие из аминогруппы (-NH2) и карбоновой кислоты (-СООН), и имеющие боковую цепь, специфичную для каждой аминокислоты. Ключевые элементы аминокислот – углерод, водород, кислород и азот. Прочие элементы находятся в боковой цепи определенных аминокислот.

Рис. 1 — Общая структура α-аминокислот , составляющих белки (кроме пролина). Составные части молекулы аминокислоты — аминогруппа NH2, карбоксильная группа COOH, радикал (различается у всех α-аминокислот), α-атом углерода (в центре).
В структуре аминокислот боковая цепь, специфичная для каждой аминокислоты, обозначается буквой R. Атом углерода, находящийся рядом с карбоксильной группой, называется альфа-углерод, и аминокислоты, боковая цепь которых связана с этим атомом, называются альфа-аминокислотами. Они представляют собой наиболее распространенную в природе форму аминокислот.
У альфа-аминокислот, за исключением глицина , альфа-углерод является хиральным атомом углерода. У аминокислот, углеродные цепи которых присоединяются к альфа-углероду (как, например, Лизин (L-лизин)), углероды обозначаются как альфа, бета, гамма, дельта, и так далее. У некоторых аминокислот аминогруппа прикреплена к бета или гамма-углероду, и поэтому они называются бета- или гамма- аминокислоты.
По свойствам боковых цепей аминокислоты подразделяются на четыре группы. Боковая цепь может делать аминокислоту слабой кислотой, слабым основанием, или эмульсоидом (если боковая цепь является полярной), или гидрофобным, плохо впитывающим воду, веществом (если боковая цепь неполярна).
Термин «аминокислота с разветвленной цепью» относится к аминокислотам, имеющим алифатические нелинейные боковые цепи, это Лейцин , Изолейцин и Валин .

Пролин – единственная протеиногенная аминокислота, боковая группа которой прикреплена к альфа-аминогруппе и, таким образом, также является единственной протеиногенной аминокислотой, содержащей на этом положении вторичный амин. С химической точки зрения, пролин, таким образом, является иминокислотой , поскольку в нем отсутствует первичная аминогруппа, хотя в текущей биохимической номенклатуре он все еще классифицируется как аминокислота, а также «N-алкилированная альфа-аминокислота» (Иминокислоты — карбоновые кислоты, содержащие иминогруппу (NH). Входят в состав белков, их обмен тесно связан с обменом аминокислот. По своим свойствам иминокислоты близки к аминокислотам, и в результате каталитического гидрирования иминокислоты превращаются в аминокислоты. Иминогруппа — молекулярная группа NH. Двухвалентна. Содержится во вторичных аминах и пептидах. В свободном виде двухвалентный радикал аммиака не существует).
АЛЬФА-АМИНОКИСЛОТЫ
Аминокислоты, имеющие как амин-, так и карбоксильную группу, прикрепляются к первому (альфа-) атому углерода имеют особое значение в биохимии. Они известны как 2-, альфа или альфа-аминокислоты (общая формула в большинстве случаев H2NCHRCOOH, где R представляет собой органический заместитель, известный как «боковая цепь»); часто термин «аминокислота» относится именно к ним.
Это 22 протеиногенных (то есть «служащих для строительства белка») аминокислоты, которые сочетаются в пептидные цепи («полипептиды»), обеспечивая построение широкого спектра белков. Они являются L-стереоизомерами («левыми» изомерами), хотя у некоторых бактерий и в некоторых антибиотиках встречаются некоторые из D-аминокислот («правых» изомеров).
Читайте также: Ткань дама с единорогом

Рис. 2. Пептидная связь — вид амидной связи, возникающей при образовании белков и пептидов в результате взаимодействия α-аминогруппы (—NH2) одной аминокислоты с α-карбоксильной группой (—СООН) другой аминокислоты.
Из двух аминокислот (1) и (2) образуется дипептид (цепочка из двух аминокислот) и молекула воды. По этой же схеме рибосома генерирует и более длинные цепочки из аминокислот: полипептиды и белки. Разные аминокислоты, которые являются «строительными блоками» для белка, отличаются радикалом R.
ОПТИЧЕСКАЯ ИЗОМЕРИЯ АМИНОКИСЛОТ

Рис. 3. Оптические изомеры аминокислоты аланина
В зависимости от положения аминогруппы относительно 2-го атома углерода выделяют α-, β-, γ- и другие аминокислоты. Для организма млекопитающих наиболее характерны α-аминокислоты. Все входящие в состав живых организмов α-аминокислоты, кроме глицина , содержат асимметрический атом углерода ( треонин и изолейцин содержат два асимметрических атома) и обладают оптической активностью. Почти все встречающиеся в природе α-аминокислоты имеют L-конфигурацию, и лишь L-аминокислоты включаются в состав белков, синтезируемых на рибосомах .
Все стандартные альфа-аминокислоты, кроме глицина, могут существовать в форме одной из двух энантиомеров , называемых L или D аминокислоты, представляющих собой зеркальные отображения друг друга.
D, L -Система обозначения стереоизомеров.

По этой системе L -конфигурация приписывается стереозомеру, у которого в проекций Фишера реперная группа находится слева от вертикальной линии (от лат. «laevus» -левый). Надо помнить, что в проекции Фишера вверху располагают наиболее окисленный атом углерода (как правило, этот атом входит в состав карбоксильной СОOН или карбонильной СН=О групп.). Кроме того, в проекции Фишера все горизонтальные связи направлены в сторону наблюдателя, а вертикальные — удалены от наблюдателя. Соответственно, если реперная группа расположена в проекции Фишера справа, стереоизомер имеет D — конфигурацию (от лат. «dexter» — правый). В α-аминокислотах реперными группами служат группы NH2.
Энантиомеры — пара стереоизомеров , представляющих собой зеркальные отражения друг друга, не совмещаемые в пространстве. Классической иллюстрацией двух энантиомеров могут служить правая и левая ладони: они имеют одинаковое строение, но различную пространственную ориентацию. Существование энантиомерных форм связано с наличием у молекулы хиральности — свойства не совмещаться в пространстве со своим зеркальным отражением. Аминокислоты являются примерами хиральных молекул .
Энантиомеры идентичны по физическим свойствам. Они могут быть различены лишь при взаимодействии с хиральной средой, например, световым излучением. Энантиомеры одинаково ведут себя в химических реакциях с ахиральными реагентами в ахиральной среде. Однако, если реагент, катализатор либо растворитель хиральны, реакционная способность энантиомеров, как правило, различается. Большинство хиральных природных соединений ( аминокислоты , моносахариды ) существует в виде 1 энантиомера. Понятие энантиомерии важно в фармацевтике , т.к. различные энантиомеры лекарств , имеют различную биологическую активность.
БИОСИНТЕЗ БЕЛКА НА РИБОСОМЕ
СТАНДАРТНЫЕ АМИНОКИСЛОТЫ
(протеиногенные)
В процессе биосинтеза белка в полипептидную цепь включаются 20 α-аминокислот , кодируемых генетическим кодом (см. рис. 4). Помимо этих аминокислот, называемых протеиногенными, или стандартными, в некоторых белках присутствуют специфические нестандартные аминокислоты, возникающие из стандартных в процессе посттрансляционных модификаций.
Прим.: В последнее время к протеиногенным аминокислотам иногда причисляют трансляционно включаемые селеноцистеин и пирролизин. Это так называемые 21-я и 22-я аминокислоты.
Аминокислоты являются структурными соединениями (мономерами), из которых состоят белки. Они объединяются между собой, формируя короткие полимерные цепи, называемые пептидами длинной цепи, полипептидами или белками. Эти полимеры являются линейными и неразветвленными, каждая аминокислота в цепи присоединяется к двум соседним аминокислотам.

Рис. 5. Рибосома в процессе трансляции (синтеза белка)
Процесс построения белка называется трансляцией и включает в себя пошаговое добавление аминокислот к растущей цепи белка через рибозимы, осуществляемый рибосомой. Порядок, в котором добавляются аминокислоты, считывается в генетическом коде с помощью шаблона мРНК , который представляет собой копию РНК одного из генов организма.
Трансляция — биосинтез белка на рибосоме

Рис. 6 Стадии элонгации полипептида.
Двадцать две аминокислоты естественно включены в полипептиды и называются протеиногенными, или природными, аминокислотами. Из них 20 кодируются с помощью универсального генетического кода.
Оставшиеся 2, селеноцистеин и пирролизин , включаются в белки при помощи уникального синтетического механизма. Селеноцистеин образуется, когда транслируемый мРНК включает SECIS элемент, вызывающий кодон UGA вместо стоп-кодона. Пирролизин используется некоторыми метаногенными археями в составе ферментов, необходимых для производства метана. Он кодируется с кодоном UAG, который в других организмах обычно играет роль стоп-кодона. За кодоном UAG следует PYLIS последовательность.

Рис. 7. Полипептидная цепь — первичная структура белка.
Белки имеют 4 уровня своей структурной организации: первичная, вторичная, третичная и четвертичная. Первичная структура — последовательность аминокислотных остатков в полипептидной цепи. Первичную структуру белка, как правило, описывают, используя однобуквенные или трёхбуквенные обозначения для аминокислотных остатков.Вторичная структура — локальное упорядочивание фрагмента полипептидной цепи, стабилизированное водородными связями.Третичная структура — пространственное строение полипептидной цепи. Структурно состоит из элементов вторичной структуры, стабилизированных различными типами взаимодействий, в которых гидрофобные взаимодействия играют важнейшую роль. Четвертичная структура (или субъединичная, доменная) — взаимное расположение нескольких полипептидных цепей в составе единого белкового комплекса.

Рис. 8. Структурная организация белков
НЕСТАНДАРТНЫЕ АМИНОКИСЛОТЫ
(Не-протеиногенные)
Помимо стандартных аминокислот существует множество других аминокислот, которые называются не-протеиногенными или нестандартными. Такие аминокислоты либо не встречаются в белках (например, L-карнитин , ГАМК ), либо не производятся непосредственно в изоляции при помощи стандартных клеточных механизмов (например, оксипролин и селенометионин).
Нестандартные аминокислоты, находящиеся в белках, образуются путем пост-трансляционной модификации, то есть модификацией после трансляции в процессе синтеза белка. Эти модификации часто необходимы для функционирования или регуляции белка; например, карбоксилирование глутамата позволяет улучшить связывание ионов кальция, а гидроксилирование пролина важно для поддержания соединительной ткани. Другой пример – формирование гипузина в фактор инициации трансляции EIF5A посредством модификации остатка лизина . Такие модификации могут также определять локализацию белка, например, добавление длинных гидрофобных групп может вызвать связывание белка с фосфолипидной мембраной.
Некоторые нестандартные аминокислоты не встречаются в белках. Это лантионин, 2-аминоизомасляная кислота, дегидроаланин и гамма-аминомасляная кислота. Нестандартные аминокислоты часто встречаются в качестве промежуточных метаболических путей для стандартных аминокислот — например, орнитин и цитруллин встречаются в орнитиновом цикле как часть катаболизма кислоты.
Редкое исключение доминированию альфа-аминокислоты в биологии — бета-аминокислота Бета-аланин (3-аминопропановая кислота), которая используется для синтеза пантотеновой кислоты (витамина B5), компонента коэнзима А у растений и микроорганизмов. Ее, в частности, продуцируют пропионовокислые бактериии .
Функции аминокислот
БЕЛКОВЫЕ И НЕ БЕЛКОВЫЕ ФУНКЦИИ
Многие протеиногенные и непротеиногенные аминокислоты также играют важную, не связанную с образованием белка, роль в организме. Например, в головном мозге человека глутамат (стандартная глутаминовая кислота) и гамма-аминомасляная кислота ( ГАМК , нестандартная гамма-аминокислота), являются основными возбуждающими и тормозящими нейромедиаторами. Гидроксипролин (основной компонент соединительной ткани коллагена) синтезируют из п ролина ; стандартная аминокислота глицин используется для синтеза порфиринов , используемых в эритроцитах. Нестандартный карнитин используется для транспорта липидов.
Из-за своей биологической значимости аминокислоты играют важную роль в питании и обычно используются в пищевых добавках, удобрениях и пищевых технологиях. В промышленности аминокислоты используются при производстве лекарств, биоразлагаемого пластика и хиральных катализаторов.
1. Аминокислоты, белки и питание
О биологической роли и последствиях дефицита аминокислот в организме человека см. информацию в таблицах незаменимых и заменимых аминокислот.
Читайте также: Гипертрофия костной ткани это
При введении в организм человека с пищей, 20 стандартных аминокислот либо используются для синтеза белков и других биомолекул, либо окисляются в мочевину и углекислый газ в качестве источника энергии. Окисление начинается с удаления аминогруппы через трансаминазу, а затем аминогруппа включается в цикл мочевины. Другой продукт трансамидирования – кетокислота, которая входит в цикл лимонной кислоты. Глюкогенные аминокислоты также могут быть преобразованы в глюкозу посредством глюконеогенеза.
Из 20 стандартных аминокислот , 8 ( валин , изолейцин , лейцин , лизин , метионин , треонин , триптофан и фенилаланин ) называют незаменимыми потому, что человеческий организм не может синтезировать их самостоятельно из других соединений в необходимых для нормального роста количествах, их можно получить только с пищей. Однако по современным представлениям Гистидин и Аргинин также являются незаменимыми аминокислотами для детей. Другие могут быть условно незаменимы для людей определенного возраста или людей, имеющих какие-либо заболевания.
Белки́ (протеины, полипептиды) — высокомолекулярные органические вещества , состоящие из альфа- аминокислот , соединённых в цепочку пептидной связью . В живых организмах аминокислотный состав белков определяется генетическим кодом , при синтезе в большинстве случаев используются 20 стандартных аминокислот .

Рис. 9. Белки не только пища. Типы белковых соединений.
Каждый живой организм состоит из белков. Различные формы белков принимают участие во всех процессах, происходящих в живых организмах. В теле человека из белков формируются мышцы, связки, сухожилия, все органы и железы, волосы, ногти; белки входят в состав жидкостей и костей. Ферменты и гормоны, катализирующие и регулирующие все процессы в организме, также являются белками. Дефицит белков в организме опасен для здоровья. Каждый белок уникален и существует для специальных целей.
ПИТАНИЕ И БЕЛКИ

Белки — важная часть питания животных и человека (основные источники: мясо, птица, рыба, молоко, орехи, бобовые, зерновые; в меньшей степени: овощи, фрукты, ягоды и грибы), поскольку в их организмах не могут синтезироваться все необходимые аминокислоты и часть должна поступать с белковой пищей. В процессе пищеварения ферменты разрушают потреблённые белки до аминокислот, которые используются для биосинтеза собственных белков организма или подвергаются дальнейшему распаду для получения энергии.

Стоит подчеркнть, что современная наука о питании утверждает, что белок должен удовлетворять потребности организма в аминокислотах не только по количеству. Данные вещества должны поступать в организм человека в определенных соотношениях между собой.
Процесс синтеза белков идет в организме постоянно. Если хоть одна незаменимая аминокислота отсутствует, образование белков приостанавливается. Это может привести к самым различным серьезным нарушениям здоровья — от расстройств пищеварения до депрессии и замедления роста у детей. Разумеется, данное рассмотрение вопроса весьма упрощенное, т.к. функции белков в клетках живых организмов более разнообразны, чем функции других биополимеров — полисахаридов и ДНК.
Также, кроме белков, из аминокислот образуется большое количество веществ небелковой природы (см. ниже), выполняющих специальные функции. К ним, напроимер, относится холин (витаминоподобное вещество, входящее в состав фосфолипидов и являющееся предшественником нейромедиатора ацетилхолина — Нейромедиаторы — это химические вещества, передающие нервный импульс с одной нервной клетки на другую. Таким образом, некоторые аминокислоты крайне необходимы для нормальной работы головного мозга).
2. Небелковые функции аминокислот
Нейромедиатор аминокислоты

Прим.: Нейромедиаторы (нейротрансмиттеры, посредники) — биологически активные химические вещества, посредством которых осуществляется передача электрохимического импульса от нервной клетки через синаптическое пространство между нейронами, а также, например, от нейронов к мышечной ткани или железистым клеткам. Для получения информации от собственных тканей и органов организм человека синтезирует особые химические вещества – нейромедиаторы. Все внутренние ткани и органы тела человека, «подчиненные» вегетативной нервной системе (ВНС), снабжены нервами (иннервированы), т. е. функциями организма управляют нервные клетки. Они как датчики собирают информацию о состоянии организма и передают ее в соответствующие центры, а от них корректирующие воздействия идут к периферии. Любое нарушение вегетативной регуляции приводит к сбоям в работе внутренних органов. Передача информации, или управление, осуществляется с помощью специальных химических веществ-посредников, которые называются медиаторами (от лат. mediator – посредник) или нейромедиаторами. По своей химической природе медиаторы относятся к различным группам: биогенным аминам, аминокислотам, нейропептидам и т. д. В настоящее время изучено более 50 соединений, относящихся к медиаторам.
В организме человека многие аминокислоты используются для синтеза других молекул, например:
- Триптофан является предшественником нейромедиатора серотонина.
- L-Тирозин и его предшественник фенилаланин являются предшественниками нейромедиаторов дофамина катехоламинов, адреналина и норадреналина.
- Глицин является предшественником порфиринов, таких как гем.
- Аргинин является предшественником оксида азота.
- Орнитин и S-аденозилметионин являются предшественниками полиаминов.
- Аспартат, Глицин и глутамин являются предшественниками нуклеотидов.
Тем не менее, все еще известны не все функции других многочисленных нестандартных аминокислот. Некоторые нестандартные аминокислоты используются растениями для защиты от травоядных животных. Например, канаванин является аналогом аргинина, который содержится во многих бобовых, и в особо крупных количествах в Canavalia gladiata (канавалия мечевидная). Эта аминокислота защищает растения от хищников, например насекомых, и при употреблении некоторых необработанных бобовых может вызывать заболевания у людей.
Классификация протеиногенных аминокислот
Рассмотрим классификацию на примере 20 протеиногенных α-аминокислот, необходимых для синтеза белка
Среди многообразия аминокислот только 20 участвует во внутриклеточном синтезе белков (протеиногенные аминокислоты). Также в организме человека обнаружено еще около 40 непротеиногенных аминокислот. Все протеиногенные аминокислоты являются α-аминокислотами. На их примере можно показать дополнительные способы классификации. Названия аминокислот обычно сокращаются до 3-х буквенного обозначения (см. рис. полипептидной цепи вверху страницы). Профессионалы в молекулярной биологии также используют однобуквенные символы для каждой аминокислоты.
1. По строению бокового радикала выделяют:
- алифатические (аланин, валин, лейцин, изолейцин, пролин, глицин) — соединения, не содержащие ароматических связей.
- ароматические (фенилаланин, тирозин, триптофан)

Ароматические соединения (арены)
— циклические органические соединения, которые имеют в своём составе ароматическую систему. Основными отличительными свойствами являются повышенная устойчивость ароматической системы и, несмотря на ненасыщенность, склонность к реакциям замещения, а не присоединения.
Различают бензоидные (арены и структурные производные аренов, содержат бензольные ядра) и небензоидные (все остальные) ароматические соединения.
Ароматичность — особое свойство некоторых химических соединений, благодаря которому сопряженное кольцо ненасыщенных связей проявляет аномально высокую стабильность;
- серусодержащие (цистеин, метионин), содержащие атом серы S
- содержащиеОН-группу (серин, треонин, опять тирозин),
- содержащие дополнительнуюСООН-группу (аспарагиновая и глутаминовая кислоты),
- дополнительнуюNH2-группу (лизин, аргинин, гистидин, также глутамин, аспарагин).
2. По полярности бокового радикала
Существуют неполярные аминокислоты (ароматические, алифатические) и полярные (незаряженные, отрицательно и положительно заряженные).
3. По кислотно-основным свойствам
По кислотно-основным свойствам подразделяют нейтральные (большинство), кислые (аспарагиновая и глутаминовая кислоты) и основные (лизин, аргинин, гистидин) аминокислоты.
4. По незаменимости
По необходимости для организма выделяют такие, которые не синтезируются в организме и должны поступать с пищей – незаменимые аминокислоты (лейцин, изолейцин, валин, фенилаланин, триптофан, треонин, лизин, метионин). К заменимым относят такие аминокислоты, углеродный скелет которых образуется в реакциях метаболизма и способен каким-либо образом получить аминогруппу с образованием сответствующей аминокислоты. Две аминокислоты являются условно незаменимыми (аргинин, гистидин), т.е.их синтез происходит в недостаточном количестве, особенно это касается детей.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом
- Правообладателям
- Политика конфиденциальности