
Специальные механизмы в сердце обеспечивают его регулярную сократительную деятельность благодаря возникновению возбуждения и распространению его по миокарду. В этой главе мы обсудим работу сердца как насоса, начиная с особенностей сердечной мышцы.
Сердце состоит из трех основных типов мышечной ткани: миокарда предсердий, миокарда желудочков и атипического миокарда проводящей системы сердца. Сокращение миокарда предсердий и желудочков имеет тот же механизм, что и сокращение скелетных мышц, но отличается большей продолжительностью. Волокна проводящей системы содержат мало миофибрилл и сокращаются слабо. Основной функцией этих волокон является автоматическая генерация импульсов и проведение их к сократительному миокарду предсердий и желудочков, что обеспечивает контроль над ритмом сердечных сокращений.
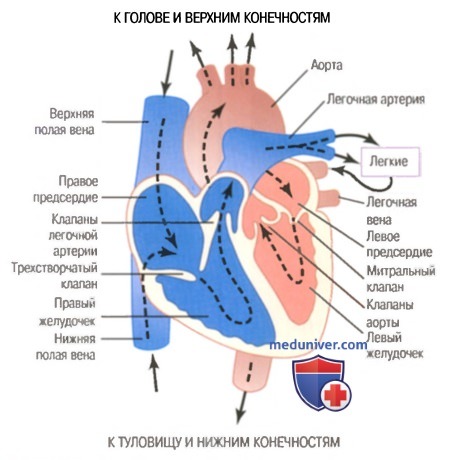 Строение сердца, направление движения крови в полостях сердца и крупных сосудах, сердечные клапаны.
Строение сердца, направление движения крови в полостях сердца и крупных сосудах, сердечные клапаны.  Синцитиальное строение миокарда.
Синцитиальное строение миокарда.
На рисунке показано гистологическое строение миокарда. Видно, что волокна сердечной мышцы образуют сеть благодаря разветвлению волокон, которые затем сливаются и разветвляются вновь. Видно также, что волокна сердечной мышцы имеют такую же поперечную исчерченность, что и волокна скелетных мышц. Более того, они содержат типичные миофибриллы, состоящие из актиновых и миозиновых филаментов. Так же, как и в волокнах скелетных мышц, эти филаменты располагаются параллельно и скользят относительно друг друга в процессе сокращения. Однако сердечная мышца имеет ряд существенных отличий от скелетной мышцы.
Сердечная мышца как синцитий. Темные зоны, пересекающие волокна сердечной мышцы на рисунке, называют вставочными дисками. Они представляют собой клеточные мембраны, которые отделяют клетки сердечной мышцы друг от друга. Таким образом, волокна миокарда состоят из большого количества отдельных кардиомиоцитов, которые соединены между собой последовательно и параллельно.
В области вставочных дисков мембраны клеток сливаются друг с другом таким образом, что формируются высокопроницаемые щелевые контакты (gap junctions), через которые свободно диффундируют ионы. Следовательно, важной функциональной особенностью миокарда является свободное движение ионов во внутриклеточной жидкости вдоль миокардиального волокна, что обеспечивает беспрепятственное распространение потенциалов действия от одной мышечной клетки к другой через вставочные диски. Таким образом, миокард представляет собой функциональное объединение (синцитий) большого количества клеток, настолько тесно взаимосвязанных между собой, что возбуждение только одной клетки приводит к распространению потенциала действия ко всем клеткам миокардиального синцития.
Сердце состоит из двух функциональных синцитиев: предсердного синцития, представленного мышечной стенкой обоих предсердий, и желудочкового синцития, представленного мышечной стенкой обоих желудочков. Предсердия отделены от желудочков фиброзной перегородкой, в которой имеются атриовентрикулярные отверстия, снабженные клапанами. Возбуждение не может пройти от предсердного синцития к желудочковому непосредственно через фиброзную ткань. Оно передается только с помощью специального атриовентрикулярного пучка диаметром в несколько миллиметров, состоящего из волокон проводящей системы сердца.
Наличие в сердце двух функциональных синцитиальных систем позволяет предсердиям сокращаться раньше, чем начнется сокращение желудочков. Это очень важно для эффективной насосной функции сердца.
Видео физиология сердечной мышцы (миокарда) — профессор, д.м.н. П.Е. Умрюхин
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Мир психологии
психология для всех и каждого
Сердечно-сосудистая система. Часть 2.





В этой части речь идет об общей схеме кровообращения, о расположении и строении сердца, о микроструктуре сердечной мышцы, об атипической ткани сердечной мышцы.
Основные особенности строения сердечно-сосудистой системы.
Общая схема кровообращения.
Система кровообращения представлена сердцем и отходящими от него сосудами, которые образуют большой и малый круги кровообращения (см. рис. 2 в правом верхнем углу).
Большой круг кровообращения начинается от левого желудочка самым крупным сосудом — аортой. Аорта разветвляется на артерии, идущие к голове (сонная артерия), верхним конечностям (подключичная артерия), туловищу (нисходящая часть аорты), ко всем внутренним органам и к нижним конечностям. Артерии разветвляются на более мелкие сосуды — артериолы, а затем капилляры, образующие густую сеть сосудов в органах и тканях.
Капилляры переходят в очень тонкие венозные сосуды — венулы. Последние идут от всех органов и тканей и соединяются в более крупные вены, которые, идя от туловища и нижних конечностей, впадают в нижнюю полую вену, а от головы и верхних конечностей — в верхнюю полую вену. Этими сосудами, впадающими в правое предсердие, заканчивается большой круг кровообращения.
Малый, или легочный, круг кровообращения начинается от правого желудочка легочной артерией, которая делится на две ветви. По этим артериям венозная кровь поступает в правое и левое легкие. Через тонкостенные капилляры легких происходит обмен газов. Кровь получает из альвеолярного воздуха кислород и отдает ему углекислый газ, т.е. превращается в артериальную. Артериальная кровь по четырем легочным венам поступает в левое предсердие, где заканчивается малый круг кровообращения (см. рис.2 в правом верхнем углу).
Кровь течет по замкнутой системе сосудов и не соприкасается с тканями. Обмен газов и питательных веществ осуществляется через жидкость, которая окружает ткани и которую называют тканевой жидкостью или тканевой плазмой.
Расположение и строение сердца.
Сердце человека расположено в грудной полости, позади грудной кости в переднем средостении, между легкими и почти полностью прикрыто ими (см. рис.3 в правом верхнем углу). Оно свободно подвешено на сосудах и может несколько смещаться.
Сердце в грудной полости располагается ассимметрично и занимает косое положение: его ось направлена справа, сверху, вперед, вниз, влево. Своим основанием сердце обращено к позвоночнику, а его верхушка упирается в пятое левое межреберье.
Сердце лежит внутри околосердечной сумки — перикарда. Она выполняет защитную роль, ограничивая растяжение сердечной мышцы. В перикарде расположены рецепторы, импульсы от которых способствуют приспособлению сердца к условиям деятельности.
Сердце состоит из двух предсердий и двух желудочков (см. рис.4 в правом верхнем углу). Правая и левая половины сердца не сообщаются между собой, и кровь через каждую из них проходит изолированно. Но правое предсердие и правый желудочек сообщаются между собой так же, как и левое предсердие с левым желудочком. Граница между предсердиями и желудочками называется атриовентрикулярной границей. В ней имеются отверстия, через которые кровь из предсердий поступает в желудочки. Эти отверстия закрыты клапанами: со стороны левого желудочка — двустворчатым (или митральным), а со стороны правого — трехстворчатым. Эти клапаны открываются только в сторону желудочков, обеспечивая поступление в них крови. При сокращении желудочков, когда в них повышается кровяное давление, клапаны плотно прилегают к отверстиям и закрывают их, препятствую поступлению крови из желудочков в предсердия. У выхода аорты и легочных артерий из желудочков расположены полулунные клапаны. Они открываются только в сосуды, обеспечивая движение крови из сердца в сосуды и препятствуя обратному току крови.
Сердечная мышца состоит из трех слоев: наружного (эпикард), внутреннего (эндокард) и среднего (миокард). Эпикард — тонкий, покрывающий сердечную мышцу слой, являющийся продолжением околосердечной сумки (ее внутренний листок). Эндокард — гладкая, эндотелиальная оболочка, выстилающая полость сердца. Миокард переставляет собой средний мышечный слой сердца, заключенный между эпикардом и эндокардом. Миокард — это особая поперечнополосатая мышца. В предсердиях он состоит из двух слоев: внутреннего, образующего правое и левое предсердия, и наружного, покрывающего оба предсердия.
Миокард желудочков состоит из трех слоев: наружного, внутреннего и среднего. Наружный мышечный слой начинается от атриовентрикулярной границы: от корней аорты и легочных артерий его волокна идут продольно к верхушке сердца, где образуют завиток, и продолжаются во внутренний мышечный слой, выстилающий полость желудочков. Средний слой миокарда образован кольцевыми мышечными волокнами, расположенными отдельно в правом и левом желудочках. Особенно сильно миокард развит в левом желудочке (см.рис.5 в правом верхнем углу).
Микроструктура сердечной мышцы.
Для понимания функциональных особенностей сердца необходимо знать строение его мышечных волокон. Клетки сердечной мышечной ткани — миоциты — почти прямоугольной формы. Их длина равна — 50-120 мк, а ширина 15-20 мк. Эти клетки имеют 1-2 ядра удлиненной формы. В периферической части цитоплазмы этих клеток особенно густо располагаются миофибриллы толщиной 1-3 мк. Миофибриллы располагаются строго прямолинейно и состоят из более мелких волокон — тонких (актиновые нити) и толстых (миозиновые нити) протофибрилл, которые создают, так же как и в поперечнополосатой скелетной мышце, поперечную исчерченность. Отличительная особенность миоцитов заключается в том, что цитоплазматическая есть у них слабее развита, чем в скелетной мышце. В сердечной мышце саркоплазматический ретикулюм сильнее выражен в волокнах, обладающих наибольшей частотой сокращения.
В сердечной мышце своеобразны контакты между двумя миоцитами — они представлены вставочными дисками, или десмосомами, в которых содержится большое количество ферментов, обеспечивающих высокий уровень энергетических процессов. Считают, что десмосомы принимают учание в передаче возбуждения от одной клетки к другой. Особенностью сердечной мышцы является наличие митохондрий. Они густо расположены между миофибриллами, в миоцитах их в 5 раз больше, чем в скелетных мышцах. Это связано с высоким уровнем обмена веществ в сердечной мышце.
Атипическая ткань сердечной мышцы.
В сердце находятся также атипичные миоциты, которые располагаются группами (узлами) и образуют проводящую систему сердца. Атипичные миоциты по своей структуре близки к эмбриональным мышечным клеткам и отличаются от миоцитов сердечной мышцы более крупными размерами ядра и самой клетки, меньшим содержанием миофибрилл и большим содержанием саркоплазмы. Их миофибриллы не имеют строгой ориентации, часто перекрещиваются друг с другом. В них мало митохондрий и рибосом. В узлах проводящей системы, помимо миоцитов, содержится много нервных клеток и волокон, их окончаний, которые образуют ганглиозную нервную сеть.
Проводящая система сердца человека представлена тремя основными узлами. Первый из них — синусно-предсердный, или синоатриальный (или, по имени исследователей, узел Кис-Флака), расположен под эпикардом в правом предсердии у места впадения верхней полой вены. От него отходит вырост, осуществляющий функциональную связь синоатриального узла со вторым узлом проводящей системы — атриовентрикулярным, или предсердно-желудочновым (или Ашофа- Товара), который расположен в правом предсердии около атриовентрикулярной перегородки и перегородки, разделяющей предсердия. Атриовентрикулярный узел переходит в пучок Гисса, начало которого располагается в верхней части межжелудочковой перегородки и называется общей ножкой пучка Гисса. Здесь же она делится на две ветви — правую и левую ножки пучка Гисса, которые, соответственно, направляются к мышцам правого и левого желудочков. Окончательные разветвления проводящей системы в виде волокон Пуркинье контактируют с мышечными волокнами миокарда.
Читайте также: Палисадная ткань с обеих сторон
Атипическая ткань сердечной мышцы.
В сердце находятся также атипичные миоциты, которые располагаются группами (узлами) и образуют проводящую систему сердца. Атипичные миоциты по своей структуре близки к эмбриональным мышечным клеткам и отличаются от миоцитов сердечной мышцы более крупными размерами ядра и самой клетки, меньшим содержанием миофибрилл и большим содержанием саркоплазмы. Их миофибриллы не имеют строгой ориентации, часто перекрещиваются друг с другом. В них мало митохондрий и рибосом. В узлах проводящей системы, помимо миоцитов, содержится много нервных клеток и волокон, их окончаний, которые образуют ганглиозную нервную сеть.
Проводящая система сердца человека представлена тремя основными узлами. Первый из них — синусно-предсердный, или синоатриальный (или, по имени исследователей, узел Кис-Флака), расположен под эпикардом в правом предсердии у места впадения верхней полой вены. От него отходит вырост, осуществляющий функциональную связь синоатриального узла со вторым узлом проводящей системы — атриовентрикулярным, или предсердно-желудочновым (или Ашофа- Товара), который расположен в правом предсердии около атриовентрикулярной перегородки и перегородки, разделяющей предсердия. Атриовентрикулярный узел переходит в пучок Гисса, начало которого располагается в верхней части межжелудочковой перегородки и называется общей ножкой пучка Гисса. Здесь же она делится на две ветви — правую и левую ножки пучка Гисса, которые, соответственно, направляются к мышцам правого и левого желудочков. Окончательные разветвления проводящей системы в виде волокон Пуркинье контактируют с мышечными волокнами миокарда.
35. Различают линейную и объемную скорость кровотока. Линейная скорость кровотока (Vлин.) это расстояние, которое проходит частица крови в единицу времени. Она зависит от суммарной площади поперечного сечения всех сосудов, образующих участок сосудистого русла. Поэтому в кровеносной системе наиболее узким участком является аорта. Здесь наибольшая линейная скорость кровотока, составляющая 0,5-0,6 м/сек. В артериях среднего и мелкого калибра она снижается до 0,2-0,4 м/сек. Суммарный просвет капиллярного русла в 500-600 раз больше чем аорты. Поэтому скорость кровотока в капиллярах уменьшается до 0,5 мм/сек. Замедление тока крови в капиллярах имеет большое физиологическое значение, так как в них происходит транскапиллярный обмен. В крупных венах линейная скорость кровотока вновь возрастает до 0,1-0,2 м/сек. Линейная скорость кровотока в артериях измеряется ультразвуковым методом. Он основан на эффекте Доплера. На сосуд помещают датчик с источником и приемником ультразвука. В движущейся среде – крови частота ультразвуковых колебаний изменяется. Чем больше скорость течения крови по сосуду, тем ниже частота отраженных ультразвуковых волн. Скорость кровотока в капиллярах измеряется под микроскопом с делениями в окуляре, путем наблюдения за движением определенного эритроцита.
Объемная скорость кровотока (Vоб.) это количество крови, проходящей через поперечное сечение сосуда в единицу времени. Она зависит от разности давлений в начале и конце сосуда и сопротивления току крови:
Vоб =———- где где Р1 и Р2 давление в начале и конце сосуда, R —
Раньше в эксперименте объемную скорость кровотока измеряли с помощью кровяных часов Людвига. В клинике объемный кровоток оценивают с помощью реовазографии. Этот метод основан на регистрации колебаний электрического сопротивления органов для тока высокой частоты, при изменении их кровенаполнения в систолу и диастолу. При увеличении кровенаполнения сопротивление понижается, а уменьшении возрастает. С целью диагностики сосудистых заболеваний производят реовазографию конечностей, печени, почек, грудной клетки. Иногда используют плетизмографию. Это регистрация колебаний объема органа, возникающих при изменении их кровенаполнения. Колебания объема регистрируют с помощью водных, воздушных и электрических плетизмографов.
Скорость кругооборота крови, это время за которое частица крови проходит оба круга кровообращения. Ее измеряют путем введения красителя флюоресцина в вену одной руки и определения времени его появления в вене другой. В среднем скорость кругооборота крови составляет 20-25 сек.Движение крови по венам.Стенки вен значительно тоньше и мягче стенок артерий. Движению крови по венам способствует сокращение скелетных мышц, окружающих вены. Кровь по венам продвигается только к сердцу. Движению ее в противоположном направлении препятствуют кармановидные полулунные клапаны, расположенные внутри вен. В единицу времени к сердцу по обеим полым венам притекает столько же крови, сколько выбрасывается им в аорту.
26. Гру́ппа кро́ви — описание индивидуальных антигенных характеристик эритроцитов, определяемое с помощью методов идентификации специфических групп углеводов и белков, включённых в мембраны эритроцитов животных. Система AB0
Известно несколько основных групп аллельных генов этой системы: A¹, A², B и 0. Генный локус для этих аллелей находится на длинном плече хромосомы 9. Основными продуктами первых трёх генов — генов A¹, A² и B, но не гена 0 — являются специфические ферменты гликозилтрансферазы, относящиеся к классу трансфераз. Эти гликозилтрансферазы переносят специфические сахара — N-ацетил-D-галактозамин в случае A¹ и A² типов гликозилтрансфераз, и D-галактозу в случае B-типа гликозилтрансферазы. При этом все три типа гликозилтрансфераз присоединяют переносимый углеводный радикал к альфа-связующему звену коротких олигосахаридных цепочек.
Субстратами гликозилирования этими гликозилтрансферазами являются, в частности и в особенности, как раз углеводные части гликолипидов и гликопротеидов мембран эритроцитов, и в значительно меньшей степени — гликолипиды и гликопротеиды других тканей и систем организма. Именно специфическое гликозилирование гликозилтрансферазой A или B одного из поверхностных антигенов — агглютиногена — эритроцитов тем или иным сахаром (N-ацетил-D-галактозамином либо D-галактозой) и образует специфический агглютиноген A или B.
В плазме крови человека могут содержаться агглютинины α и β, в эритроцитах — агглютиногены A и B, причём из белков A и α содержится один и только один, то же самое — для белков B и β.
Таким образом, существует четыре допустимых комбинации; то, какая из них характерна для данного человека, определяет его группу крови [1] :α и β: первая (0)A и β: вторая (A)α и B: третья (B)A и B: четвёртая (AB)Система Rh (резус-система)Резус-фактор — это антиген (белок), который находится на поверхности красных кровяных телец (эритроцитов). Он обнаружен в 1940 году Карлом Ландштейнером и А.Вейнером [2] . Около 85 % европейцев (99 % индейцев и азиатов) имеют резус-фактор и соответственно являются резус-положительными. Остальные же 15 % (7 % у африканцев), у которых его нет, — резус-отрицательный. Резус-фактор играет важную роль в формировании так называемой гемолитической желтухи новорожденных, вызываемой вследствие резус-конфликта иммунизованной матери и эритроцитов плода.Известно, что резус-фактор — это сложная система, включающая более 40 антигенов, обозначаемых цифрами, буквами и символами. Чаще всего встречаются резус-антигены типа D (85 %), С (70 %), Е (30 %), е (80 %) — они же и обладают наиболее выраженной антигенностью. Система резус не имеет в норме одноименных агглютининов, но они могут появиться, если резус-отрицательному человеку перелить резус-положительную кровь Переливание несовместимой крови вызывает тяжелейшее осложнение – гемотрансфузионный шок. Он возникает вследствие того, что склеившиеся эритроциты закупоривают мелкие сосуды. Кровоток нарушается. Затем происходит их гемолиз и из эритроцитов донора в кровь поступают чужеродные белки. В результате резко падает кровяное давление, угнетается дыхание, сердечная деятельность, нарушается работа почек, центральной нервной системы. Переливание даже небольших количеств такой крови может закончиться смертью реципиента.
В настоящее время допускается переливание только одногрупповой крови по системе АВО. Обязательно учитывается и ее резус-принадлежность. Поэтому перед каждым переливанием обязательно проводится определение группы и D-антигена крови донора и реципиента. Для определения групповой принадлежности, каплю исследуемой крови смешивают на предметном стекле с каплей стандартных сывороток I, II и III групп. Таким методом определяются антигенные свойства эритроцитов. Если ни в одной из сывороток не произошла агглютинация, следовательно в эритроцитах агглютиногенов нет. Это кровь I группы. Когда агглютинация наблюдается с сыворотками I и III групп, значит эритроциты исследуемой крови содержат агглютиноген А. Т.е. это кровь II группы. Агглютинация эритроцитов с сыворотками I и II групп говорит о том, что в них имеется агглютиноген В и эта кровь III группы. Если во всех сыворотках наблюдается агглютинация, значит эритроциты содержат оба антигена А и В. Т.е. кровь IV группы. Желательно проводить исследование и с сывороткой IV группы. Более точно группу крови можно определить с помощью стандартных эритроцитов I, II, III и IV групп. Для этого их смешивают с сывороткой исследуемой крови и определяют содержание в ней агглютининов. Резус принадлежность крови определяют путем ее смешивания с сывороткой, содержащей резус-агглютинины.
Это сложный физиологический процесс, протекающий в несколько фаз. Главные его участники — это стенка сосуда, нервная система и тромбоциты крови. Первичный гемостаз начинается прежде всего с первичного сосудистого спазма рефлекторной природы. Затем начинается так называемая эндотелиально—тромбоцитарная реакция. На месте травмы эндотелий сосуда меняет свой заряд. Тромбоциты, занимающие в сосуде краевое положение, начинают адгезировать (прилипать) к поврежденной поверхности сосуда и агглютинировать (склеиваться) между собой. В результате через 2—3 минуты наступает третья фаза — фаза образования «тромбоцитарного гвоздя». В течение этой фазы происходит остановка кровотечения, однако свертывания крови еще не произошло; плазма крови остается жидкой. Образовавшийся тромб рыхлый, и еще в течение короткого времени процессы имеют обратимый характер. Четвертая фаза заключается в том, что в образовавшемся тромбе начинаются морфологические превращения тромбоцитов, которые приведут к их необратимым изменениям и разрушению. Это вязкий метаморфоз тромбоцитов. В результате вязкого метаморфоза из тромбоцитов выходят содержащиеся там факторы свертывания. Их взаимодействие приводит к появлению следов тромбина, который и запускает каскад химических ферментативных реакций — ферментативное свертывание.
Появление следов тромбина запускает сложный процесс так называемого ферментативного свертывания.
Первая фаза ферментативного свертывания начинается в результате многоступенчатого взаимодействия кровяных и тканевых факторов свертывания, когда в крови появляется ранее отсутствующий фактор — тромбопластин. Вторая фаза — это взаимодействие тромбопластина с протромбином — неактивным предшественником тромбина. В результате взаимодействия тромбопластина и протромбина в присутствии солей кальция в крови появляется активный тромбин в концентрации достаточной, чтобы начался этап свертывания — взаимодействие тромбина с растворимым фибриногеном и переход последнего в нерастворимый фибрин. Это и есть третья фаза. По появлению первых нитей фибрина в клинике определяют время свертывания крови.
Читайте также: Изготовление лица куклы из ткани
Таким образом, процесс ферментативного свертывания крови протекает в три стадии: 1 — образование активного тромбопластина, 2 —появление активного тромбина и 3 — выпадение в осадок нерастворимых нитей фибрина.
Затем начинается следующая ферментативная стадия, в течение которой происходит уплотнение и сжатие кровяного сгустка, отделение прозрачной, жидкой, потерявшей способность к свертыванию сыворотки. Это четвертая стадия свертывания крови — ретракция (сжатие) кровяного сгустка. И, наконец, наступает последняя пятая стадия — лизис (растворение) тромба. Это также многоступенчатый процесс, в котором протекают ферментативные взаимодействия многих веществ, в конечном итоге приводящих к появлению активного фермента — фибринолизина. Фибринолизин разрушает связи между нитями фибрина и переводит его вновь в нерастворимый фибриноген. В настоящее время принято говорить о существовании самостоятельной фибринолитической системы организма. Конечно, эти процессы в организме протекают значительно сложнее, и в них принимают участие гораздо большее число факторов. Антикоагулянты (от анти… и лат. coagulans, род. падеж coagulantis — вызывающий свёртывание) — химические вещеcтва и лекарственные средства, угнетающие активность свёртывающей системы крови и препятствующие образованию тромбов. Антикоагулянты оказывают влияние на различные звенья процесса свёртывания крови. Различают антикоагулянты прямого действия (гепарин, гирудин и др.), понижающие активность тромбина в крови, и антикоагулянты непрямого действия (дикумарин, варфарин, неодикумарин, или пелентан, фенилин, синкумар и др.), нарушающие образование протромбина в печени, участвующего в свёртывании крови. Антикоагулянты применяют при инфарктах миокарда и лёгких, тромботических и эмболических инсультах, тромбофлебитах и другие; профилактически — при атеросклерозе коронарных артерий, мозговых сосудов, ревматических митральных пороках сердца; в хирургии — для предупреждения образования тромбов в послеоперационном периоде; в гематологии — для использования с устройствами автоматического афереза для заготовки компонентов крови человека (эритроциты, тромбоциты, плазма).
29.Активная реакция крови — чрезвычайно важная гомеостатическая константа организма, обеспечивающая течение окислительно-восстановительных процессов, деятельность ферментов, направление и интенсивность всех видов обмена.
Кислотность или щелочность раствора зависит от содержания в нем свободных ионов водорода [Н+]. Количественно активная реакция крови характеризуется водородным показателем — рН (power hydrogen — «сила водорода»).
Водородный показатель — отрицательный десятичный логарифм концентрации водородных ионов, т. е. pH = -lg[H+].
Символ рН и шкалу рН (от 0 до 14) ввел в 1908 г. Сервисен. Если рН равно 7,0 (нейтральная реакция среды), то содержание ионов Н+ равно 107 моль/л. Кислая реакция раствора имеет рН от 0 до 7; щелочная — от 7 до 14.
Кислота рассматривается как донор ионов водорода, основание — как их акцептор, т. е. вещество, которое может связывать ионы водорода.
Постоянство кислотно-основного состояния (КОС) поддерживается как физико-химическими (буферные системы), так и физиологическими механизмами компенсации (легкие, почки, печень, другие органы).
Буферными системами называют растворы, обладающие свойствами достаточно стойко сохранять постоянство концентрации водородных ионов как при добавлении кислот или щелочей, так и при разведении.
Буферная система — это смесь слабой кислоты с солью этой кислоты, образованной сильным основанием.
Примером может служить сопряженная кислотно-основная пара карбонатной буферной системы: H2CO3 и NaHCO3.
В крови существует несколько буферных систем: бикарбонатная (смесь H2CO3 и HCO3-); система гемоглобин — оксигемоглобин (оксигемоглобин имеет свойства слабой кислоты, а дезоксигемоглобин — слабого основания); белковая (обусловленная способностью белков ионизироваться); фосфатная система (дифосфат — монофосфат).
Самой мощной является бикарбонатная буферная система — она включает 53% всей буферной емкости крови, остальные системы составляют соответственно 35%, 7% и 5%. Особое значение гемоглобинового буфера заключается в том, что кислотность гемоглобина зависит от его оксигенации, то есть газообмен кислорода потенцирует буферный эффект системы.
Исключительно высокую буферную емкость плазмы крови можно проиллюстрировать следующим примером. Если 1 мл децинормальной соляной кислоты добавить к 1 л нейтрального физиологического раствора, который не является буфером, то его рН упадет с 7,0 до 2,0. Если такое же количество соляной кислоты добавить к 1 л плазмы, то рН снизится всего с 7,4 до 7,2.
Роль почек в поддержании постоянства кислотно-основного состояния заключается в связывании или выведении ионов водорода и возвращении в кровь ионов натрия и бикарбоната. Механизмы регуляции КОС почками тесно связаны с водно-солевым обменом. Метаболическая почечная компенсация развивается гораздо медленнее дыхательной компенсации — в течение 6-12 ч.
Постоянство кислотно-основного состояния поддерживается также деятельностью печени. Большинство органических кислот в печени окисляется, а промежуточные и конечные продукты либо не имеют кислого характера, либо представляют собой летучие кислоты (углекислота), быстро удаляющиеся легкими. Молочная кислота в печени преобразуется в гликоген (животный крахмал). Большое значение имеет способность печени удалять неорганические кислоты вместе с желчью.
Выделение кислого желудочного сока и щелочных соков (панкреатического и кишечного) также имеет значение в регуляции КОС.
Огромная роль в поддержании постоянства КОС принадлежит дыханию. Через легкие в виде углекислоты выделяется 95% образующихся в организме кислых валентностей. За сутки человек выделяет около 15 000 ммоль углекислоты, следовательно, из крови исчезает примерно такое же количество ионов водорода (H2CO3 = CO2↑ + H2O). Для сравнения: почки ежедневно экскретируют 40-60 ммоль Н+ в виде нелетучих кислот.
Количество выделяемой двуокиси углерода определяется ее концентрацией в воздухе альвеол и объемом вентиляции. Недостаточная вентиляция приводит к повышению парциального давления CO2 в альвеолярном воздухе (альвеолярная гиперкапния) и соответственно увеличению напряжения углекислого газа в артериальной крови (артериальная гиперкапния). При гипервентиляции происходят обратные изменения — развивается альвеолярная и артериальная гипокапния.
Таким образом, напряжение углекислого газа в крови (РаCO2), с одной стороны, характеризует эффективность газообмена и деятельность аппарата внешнего дыхания, с другой — является важнейшим показателем кислотно-основного состояния, его дыхательным компонентом.
Респираторные сдвиги КОС самым непосредственным образом участвуют в регуляции дыхания. Легочный механизм компенсации является чрезвычайно быстрым (коррекция изменений рН осуществляется через 1-3 мин) и очень чувствительным.
При повышении РаСО2 с 40 до 60 мм рт. ст. минутный объем дыхания возрастает от 7 до 65 л/мин. Но при слишком большом повышении РаCO2 или длительном существовании гиперкапнии наступает угнетение дыхательного центра с понижением его чувствительности к СОг.
При ряде патологических состояний регуляторные механизмы КОС (буферные системы крови, дыхательная и выделительная системы) не могут поддерживать рН на постоянном уровне. Развиваются нарушения КОС, и в зависимости от того, в какую сторону происходит сдвиг рН, выделяют ацидоз и алкалоз.
В зависимости от причины, вызвавшей смещение рН, выделяют дыхательные (респираторные) и метаболические (обменные) нарушения КОС: дыхательный ацидоз, дыхательный алкалоз, метаболический ацидоз, метаболический алкалоз.
Системы регуляции КОС стремятся ликвидировать возникшие изменения, при этом респираторные нарушения нивелируются механизмами метаболической компенсации, а метаболические нарушения компенсируются изменениями вентиляции легких.
74. Регуляция мочеобразования осуществляется нейрогуморальным путем. Высшим подкор-ковым центром регуляции мочеобразования является гипоталамус. Импульсы от рецепторов почек по симпатическим нервам посту-пают в гипоталамус, где вырабатывается ан-тидиуретический гормон (АДГ) или вазо-прессин, усиливающий реабсорбцию воды из первичной мочи и являющийся основным компонентом гуморальной регуляции. Этот гормон поступает в гипофиз, там накаплива-ется и затем выделяется в кровь. Повышение секреции АДГ сопровождается увеличением проницаемости извитых канальцев и собира-тельных трубок для воды. Усиленная реаб-сорбция воды при недостаточном ее поступ-лении в организм приводит к снижению диу-роеза; моча при этом характеризуется высокой концентрацией находящихся в ней ве-ществ. При избытке воды в организме осмо-тическое давление плазмы падает. Через ос-мо- и ионорецепторы гипоталамуса и почек происходит рефлекторное снижение продук-ции АДГ и его поступления в кровь. В этом случае организм избавляется от избытка воды путем выделения большого количества мочи низкой концентрации. Существенное значе-ние в гуморальной регуляции мочеобразова-ния принадлежит гормону коры надпочечни-ков альдостерону (из группы минерало-кортикоидов), который увеличивает реаб-сорбцию ионов Nа* и секрецию ионов К+, уменьшая диурез.
Нервная регуляция мочеобразования выраже-на слабее, чем гуморальная, и осуществляется как условнорефлекторным, так и безус-ловнорефлекторным путем. В основном она происходит благодаря рефлекторным измене-ниям просвета почечных сосудов под влияни-ем различных воздействий на организм. Это ведет к сдвигам почечного кровотока и, сле-довательно, процесса мочеобразования. Ус-ловнореф-лекторное повышение диуреза на индифферентный раздражитель, подкреплен-ное повышенным потребление воды, свиде-тельствует об участии коры больших полу-шарий в регуляции мочеобразования. Следует иметь в виду, что почки обладают высокой способностью к
саморегуляции. Выключение высших корко-вых и подкорковых центров регуляции не приводит к прекращению мочеобразования.
71. Температура тела человека и высших животных поддерживается на относительно постоянном уровне, несмотря на колебания температуры окружающей среды. Это постоянство температуры тела носит название изотермии. Изотермия свойственна только так называемым гомойотермным, или теплокровным, животным. Изотермия отсутствует у пойкилотермных, или холоднокровных, животных, температура тела которых переменна и мало отличается от температуры окружающей среды. Изотермия в процессе онтогенеза развивается постепенно. У новорожденного ребенка способность поддерживать постоянство температуры тела далеко не совершенна. Вследствие этого может наступить охлаждение организма при таких темпе-ратураА окружающей среды, которые не оказывают влияния на взрослого человека. Равным образом даже/небольшая мышечная работа, например, связанная с длительным криком ребенка, может повысить температуру тела. Организм недоношенных детей еще менее способен поддерживать П0етоянство температуры тела, которая у них в значительной мере зависит от температуры среды /обитания. Температура органов и тканей, как и всего организма в целом, зависит от интенсивности образования тепла и от величины теплопотерь. Теплообразование происходит вследствие непрерывно совершающихся экзотермических реакций. Эти реакции протекают во всех органах и тканях, но неодинаково интенсивно. В тканях и органах, производящих активную работу — в мышечной ткани, печени, почках, выделяется большее количество тепла, чем в менее активных —- соединительной ткани, костях, хрящах. Потеря тепла органами и тканями зависит в большой степени от их месторасположения: поверхностно расположенные органы, например кожа, скелетные мышцы, отдают больше тепла и охлаждаются сильнее, чем внутренние органы, более защищенные от охлаждения. Отсюда ясно, что температура разных органов различна. Так, печень, расположенная глубоко внутри тела и дающая большую теплопродукцию, имеет у человека более высокую и постоянную температуру (37,8—38 °С) по сравнению с кожей, температура которой значительно ниже (на покрытых одеждой участках 29,5—33,9 °С) и в большей мере зависит от окружающей среды. Поэтому справедливо говорить о том, что изотермия присуща главным образом внутренним органам и головному мозгу. Поверхность же тела и конечности, температура которых может несколько изменяться в зависимости от температуры окружающей среды, являются в некоторой мере пойкилотермными. При этом различные участки кожной поверхности имеют неодинаковую температуру. Обычно относительно выше температура кожи туловища и головы (33—34 °С). Температура конечностей ниже, причем она наиболее низкая в дистальных отделах. Из сказанного следует, что понятие «постоянная температура тела» является условным. Лучше всего среднюю температуру организма как целого характеризует температура крови в наиболее крупных сосудах, так как циркулирующая в них кровь нагревается в активных тканях (тем самым охлаждая их) и охлаждается в коже (одновременно согревая ее). О температуре тела человека судят обычно на основании ее измерения в подмышечной впадине. Здесь температура у здорового человека равна 36,5—36,9 °С. В клинике часто (особенно у грудных детей) измеряют температуру в прямой кишке, где она выше, чем в подмышечной впадине, и равна у здорового человека в среднем 37,2—37,5 °С. Температура тела не остается постоянной, а колеблется в течение суток в пределах 0,5—0,7 °С. Покой и сон понижают температуру, мышечная деятельность повышает ее. Максимальная температура тела наблюдается в 4—6 ч вечера, минимальная — в 3—4 ч утра. У рабочих, длительно работающих в ночных сменах, колебания температуры могут быть обратными указанным выше. Постоянство температуры тела у человека может сохраняться лишь при условии равенства теплообразования и теплопотери всего организма. Это достигается с помощью физиологических механизмов терморегуляции. Терморегуляция проявляется в форме взаимосочетания процессов теплообразования и теплоотдачи, регулируемых нервно-эндокринным путем. Терморегуляцию принято разделять на химическую и физическую. Химическая терморегуляция осуществляется путем изменения уровня теплообразования, т. е. усиления или ослабления интенсивности обмена веществ в клетках организма. Физическая терморегуляция осуществляется путем изменения интенсивности отдачи тепла.
ХИМИЧЕСКАЯ ТЕРМОРЕГУЛЯЦИЯ
Химическая терморегуляция имеет важное значение для поддержания постоянства температуры тела, как в нормальных условиях, так и при изменении температуры окружающей среды. У человека усиление теплообразования вследствие увеличения интенсивности обмена веществ отмечается, в частности, тогда, когда температура окружающей среды становится ниже оптимальной температуры, или зоны комфорта. При обычной легкой одежде эта зона находится в пределах 18—20 °С, а для обнаженного человека 28 °С. Оптимальная температура во время пребывания в воде выше, чем на воздухе. Это обусловлено тем, что вода, обладающая высокой теплоемкостью и теплопроводностью, охлаждает тело в 14 раз сильнее, чем воздух. Поэтому в прохладной ванне обмен веществ повышается значительно больше, чем во время пребывания на воздухе при той же температуре. Наиболее интенсивное теплообразование в организме происходит в мышцах. Даже если человек лежит неподвижно, но с напряженной мускулатурой, окислительные нродес-сы, а вместе с тем и теплообразование повышаются на 10%. Небольшая двигательная активность ведет к увеличению теплообразования на 50—80%, а тяжелая мышечная работа — на 400—500%. В условиях холода теплообразование в мышцах увеличивается, даже если человек находится в неподвижном состоянии. Это обусловлено тем, что охлаждение поверхности тела, действуя на рецепторы, воспринимающие холодовое раздражение, рефлекторно возбуждает беспорядочные непроизвольные сокращения мышц, проявляющиеся в виде дрожи (озноб). При этом обменные процессы организма значительно усиливаются, увеличивается потребление кислорода и углеводов мышечной тканью, что и влечет за собой повышение теплообразования. Даже произвольная имитация дрожи увеличивает теплообразование на 200%. Если в организм введены миорелаксанты — вещества, нарушающие передачу нервных импульсов с нерва на мышцу и тем самым устраняющие рефлекторную мышечную дрожь, при понижении температуры окружающей среды гораздо быстрее наступает понижение температуры тела. В химической терморегуляции, кроме мышц, значительную роль играют печень и почки. Температура крови печеночной вены выше температуры крови печеночной артерии, что указывает на интенсивное теплообразование в этом органе. При охлаждении тела теплопродукция в печени возрастает. Освобождение энергии в организме совершается за счет окислительного распада белков, жиров и углеводов. Поэтому все механизмы, которые регулируют окислительные процессы, регулируют и теплообразование.
ФИЗИЧЕСКАЯ ТЕРМОРЕГУЛЯЦИЯ
Физическая терморегуляция осуществляется путем изменений отдачи тепла организмом. Особо важное значение она приобретает в поддержании постоянства температуры тела во время пребывания организма в условиях повышенной температуры окружающей среды. Теплоотдача осуществляется путем теплоизлучения (радиационная теплоотдача), конвекции, т. е. движения и перемешивания нагреваемого телом воздуха, теплопроведе-ния, т. е. отдачи тепла веществам, непосредственно соприкасающимся с поверхностью тела, /И испарения воды с поверхности кожи и легких. У человека в обычных условиях потеря тепла путем теплопроведения имеет небольшое/значение, так как воздух и одежда являются плохими проводниками тепла. Радиация, испарение и конвекция протекают с различной интенсивностью в зависимости от температуры окружающей среды. У человека в состоянии покоя при температуре воздуха ‘ около 20 °С и суммарной теплоотдаче, равной 419 кДж (100 ккал) в час, радиация составляет 66%, испарение воды —19%, конвекция -15% общей потери тепла организмом. При повышении температуры окружающей среды до 35 °С теплоотдача посредством радиации и конвекции становится невозможной и температура тела поддерживается на постоянном уровне исключительно посредством испарения воды с поверхности кожи и альвеол легких. Для того чтобы было ясно значение испарения в теплоотдаче, напомним, что для испарения 1 мл воды необходимо 2,4 кДж (0,58 ккал). Следовательно, если в условиях основного обмена телом человека отдается посредством испарения около 1675—2093 кДж (400—500 ккал), то с поверхности тела должно испаряться примерно 700—850 мл воды. Из этого количества 300—350 мл испаряются в легких и 400—500 мл — с поверхности кожи. Характер отдачи тепла телом изменяется в зависимости от интенсивности обмена веществ. При увеличении теплообразования в результате мышечной работы возрастает значение теплоотдачи, осуществляемой посредством испарения воды. Так, после тяжелого спортивного соревнования, когда суммарная теплоотдача достигала почти 2512 кДж (600 ккал) в час, было найдено, что 75% тепла было отдано путем испарения, 12% — путем радиации и 13 % —посредством конвекции. Одежда уменьшает теплоотдачу. Потере тепла препятствует тот слой неподвижного воздуха, который находится между одеждой и кожей, так как воздух — плохой проводник тепла. Теплоизолирующие свойства одежды тем выше, чем более мелкоячеиста ее структура, содержащая воздух. Этим объясняются хорошие теплоизолирующие свойства шерстяной и меховой одежды. Температура воздуха под одеждой достигает 30 °С. Наоборот, обнаженное тело теряет тепло, потому что воздух на его поверхности все время сменяется. Поэтому температура кожи обнаженных частей тела намного ниже, чем одетых. В значительной степени препятствует теплоотдаче слой подкожной жировой клетчатки в связи с малой теплопроводностью жира. Температура кожи, а следовательно, интенсивность теплоизлучения и теплопрове-дения могут изменяться в результате перераспределения крови в сосудах и при изменении объема циркулирующей крови. На холоде кровеносные сосуды кожи, главным образом артериолы, сужаются; большее количество крови поступает в сосуды брюшной полости и тем самым ограничивается теплоотдача. Поверхностные слои кожи, получая меньше теплой крови, излучают меньше тепла —теплоотдача уменьшается. При сильном охлаждении кожи, кроме того, происходит открытие артериовенозных анастомозов, что уменьшает количество крови, поступающей в капилляры, и тем самым препятствует теплоотдаче. Перераспределение крови, происходящее на холоду—уменьшение количества крови, циркулирующей через поверхностные сосуды, и увеличение количества крови, проходящей через сосуды внутренних органов, способствует сохранению тепла во внутренних органах. Эти факты служат основанием для утверждения, что регулируемым параметром является именно температура внутренних органов, которая поддерживается на постоянном уровне. При повышении температуры окружающей среды сосуды кожи расширяются, количество циркулирующей в них крови увеличивается. Возрастает также объем циркулирующей крови во всем организме вследствие перехода воды из тканей в сосуды, а также потому, что селезенка и другие кровяные депо выбрасывают в общий кровоток дополнительные количества крови. Увеличение количества крови, циркулирующей через сосуды поверхности тела, способствует теплоотдаче посредством радиации и конвекции. Для сохранения постоянства температуры тела человека при высокой температуре окружающей среды основное значение имеет испарение пота с поверхности кожи. Значение потоотделения для поддержания постоянства температуры тела видно из следующего подсчета: в летние месяцы температура окружающего воздуха в средних широтах нередко равна температуре тела человека. Это означает, что организм человека, живущего в этих условиях, не может отдавать образующееся в нем самом тепло путем радиации и конвекции. Единственным путем для отдачи тепла остается испарение воды. Приняв, что среднее теплообразование в сутки равно 10 048— 11 723 кДж (2400—2800 ккал), и зная. что на испарение 1 г воды с поверхности тела расходуется 2,43 кДж (0,58 ккал), получаем, что для поддержания температуры тела человека на постоянном уровне в таких условиях необходимо испарение 4,5 л воды. Особенно интенсивно потоотделение происходит при высокой окружающей температуре во время мышечной работы, когда возрастает теплообразование в самом организме. При очень тяжелой работе выделение пота у рабочих горячих цехов может составить 12 л за день. Испарение воды зависит от относительной влажности воздуха. В насыщенном водяными парами воздухе вода испаряться не может. Поэтому при высокой влажности атмосферы высокая температура переносится тяжелее, чем при низкой влажности. В насыщенном водяными парами воздухе (например, в бане) пот выделяется в большом количестве, но не испаряется и стекает с кожи. Такое потоотделение не способствует отдаче тепла; только эта часть пота, которая испаряется с поверхности кожи, имеет значение для теплоотдачи (эта часть пота составляет эффективное потоотделение). Плохо переносится также непроницаемая для воздуха одежда (резиновая и т. п.), препятствующая испарению пота: слой воздуха между одеждой и телом быстро насыщается парами и дальнейшее испарение пота прекращается.