
БОТАНИКА ТОМ 1 — КЛЕТОЧНАЯ БИОЛОГИЯ. АНАТОМИЯ. МОРФОЛОГИЯ — 2007
2. СТРОЕНИЕ И УЛЬТРАСТРУКТУРА КЛЕТКИ
2.2. Растительная клетка
На рис. 2.2, 2.6—2.8 показаны типичные растительные клетки, видимые с помощью светового и электронного микроскопа. Здесь одновременно представлены важнейшие структурные компоненты растительных клеток. Прежде всего дадим четкие определения клеточных компонентов, а строение, функции и генезис отдельных органелл детально будут рассмотрены в последующих разделах. Под органеллами мы понимаем внутриклеточные функциональные единицы (от лат organellum — маленький аппарат, поэтому в единственном числе корректнее было бы говорить «органелл»; однако это слово употребляется в женском роде) 1 .
1 Исходное слово в латыни — среднего рода. При заимствованиях грамматический род может не сохраняться — Примеч. ред.
Рис. 2.6. Ультраструктура растительной клетки.
А—зародышевая клетка клеточная стенка со срединной пластинкой и плазмодесмами, в цитоплазме две диктиосомы, гладкий и шероховатый ЭР, рибосомы и полисомы, различные пузырьки (в том числе и coated vesicles) и липидные тельца (олеосомы) Под клеточной мембраной местами микротрубочки, в продольном и поперечном сечении, вакуоли; в расположенном в центре клеточном ядре ядрышко и плотный хроматин; две пропластиды (с пластоглобулами и крахмалом) и митохондрия (с кристами). Органеллы содержат собственную ДНК; неплазматические компоненты показаны белым (см. бокс 2 3) В—фрагмент среза клетки ткани (например, клетки листа) с сильно увеличенной вакуолью Разросшаяся первичная стенка (саккодерма), возле углов клеток межклеточные пространства — межклетники (показаны точками), в цитоплазме рядом с митохондрией, шероховатым ЭР и олеосомами — пероксисома с кристаллом каталазы, а также хлоропласт с тилакоидами, пластоглобулами и крахмальным зерном. CV—везикулы (coated vesicles); D—диктиосомы, ER—эндоплазматический ретикулум (ЭР), S—крахмал, V—вакуоль

Клеточная стенка (англ, cell wall). Окружает тело живой клетки (+ протопласт) как формообразующий внешний скелет (экзоскелет), содержит прочные фибриллы из целлюлозы или хитина, пронизана тонкими каналами (плазмодесмами) — плазматическими соединениями между соседними клетками (греч. désmos — узы, связь).
Клеточная мембрана (плазматическая мембрана, плазмалемма; англ, cell membrane). Биомембрана (от лат. membrana — кожа), окружающая весь протопласт, как и большинство биомембран, обладает избирательной проницаемостью: пропускает воду и незаряженные молекулы, напротив же, ионы и более крупные полярные частицы проходят через мембрану только тогда, когда для них имеются специфические переносчики.
Биомембраны представляют собой вязкую жидкость (толщина 6 — 11 нм). Главный элемент всех биомембран — двойной липидный слой, пронизанным поперек интегральными мембранными белками; периферические мембранные белки прикреплены к поверхности. Биомембраны ограничивают структурные элементы, или компартменты (англ, compartments), окружая их сплошным слоем, — таким образом, у биомембран нет боковых краев. Они отделяют «внутреннее» от «внешнего» биомембрана — это не лист, а «воздушный шар».
Цитоплазма (англ, cytoplasm) — основная масса протопласта, имеющая консистенцию от вязкой до слизистой. В ней расположены различные органеллы; место многих реакций обмена веществ; выпадает при фракционировании клетки как цитозоль («растворимая фракция»).
Цитоскелет (англ, cytoskeleton) — внутренний скелет (эндоскелет), может в отдельных местах укреплять цитоплазму (золь → гель); с другой стороны, обеспечивает с помощью моторных молекул процессы движения внутри клетки (например, ток цитоплазмы, перемещение пузырьков, движение хромосом при ядерном делении); у растений это прежде всего микротрубочки и актиповые микрофиламенты (лат. túbulus — трубочка, filum — нить).
Рибосомы — мелкие (30 нм) плотные частицы в цитоплазме и на мембранах эндоплазматической сети (от греч. soma — тельце, частица), чаще всего объединены в полисомы.Органеллы биосинтеза белка (трансляции).
Эндоплазматическая сеть (эндоплазматический ретикулум — ЭС, ЭР) (от лат. reticulum — сеть) пронизывает цитоплазму в виде разветвленной мембранной системы. Существует в клетке в двух формах: шероховатый ЭР (rER) — на мембранах внешней стороны располагаются полисомы, и гладкий ЭР (sER, от англ, smooth — гладкий), без полисом. Внутренние пространства шероховатого ЭР чаще всего плоские, и мембраны ЭР проходят параллельно через цитоплазму («двойные мембраны»); в этом случае говорят о цистернах. Типичный пример: ядерная оболочка = перинуклеарная цистерна (англ, nuclear envelope).
Диктиосомы (от греч. diktyon — сеть) — мелкие стопки свободных от рибосом цистерн (цистерны Гольджи), которые получают материал от шероховатого ЭР путем притока мелких пузырьков, перерабатывают его в секреты (например, материал клеточной стенки) и через пузырьки Гольджи доставляют к клеточной мембране; там происходит выделение секретов наружу (экзоцитоз)
Диктиосомы являются элементами аппарата Гольджи, названного в честь К. Гольджи, открывшего органеллы биосинтеза белка.
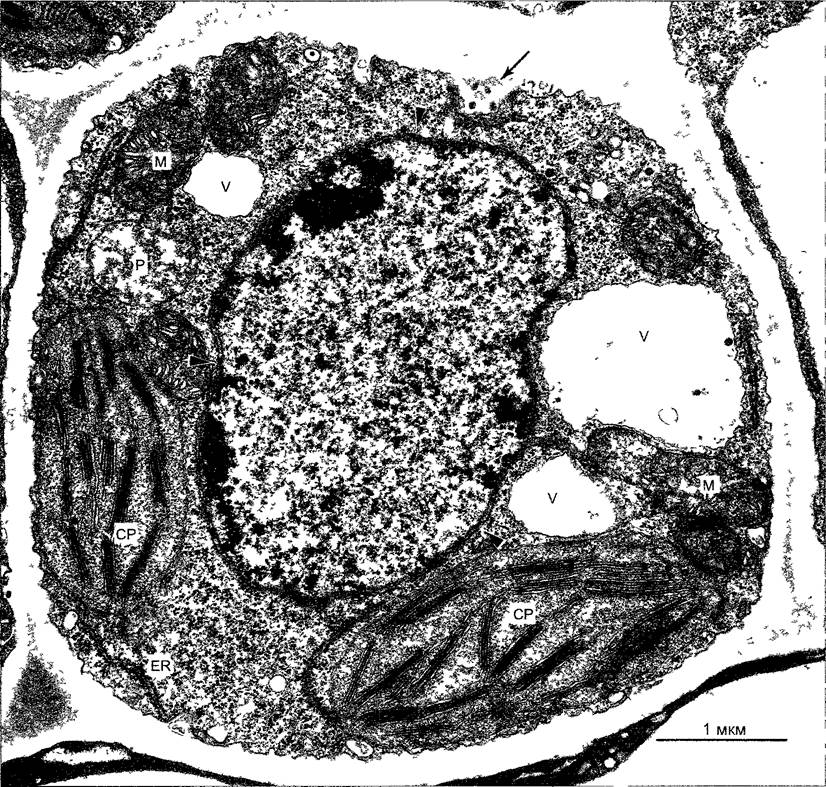
Рис. 2.7. Электронная микрофотография растительной клетки (ультратонкий срез, клетка паренхимы флоэмы фасоли Phaseolus vulgaris) (препарат и электронная микрофотосъемка Н. Falk)
Видны особенности молодой клетки, где идет активный обмен веществ (многочисленные мелкие вакуоли, в цитоплазме много рибосом/полисом), однако, с другой стороны, здесь имеются хлоро- пласты, митохондрии и пероксисомы Ядрышко находится вне плоскости среза, стрелками в виде треугольников показаны ядерные поры Стрелка—плазмодесмы в поперечном сечении По соседству с диктиосомами четыре мембранных пузырька В центре ядра преимущественно рыхлый эухроматин, у ядерной оболочки местами плотный гетерохроматин, СР—хлоропласты, М—митохондрии; Р—пероксисома, остальные обозначения, как на рис 2 6
С помощью ультрацентрифуги можно получить однородные фракции внутриклеточных частиц для биохимических или аналитических исследований (рис А). От сохранения клеточных структур при этом конечно приходится отказаться. Более крупные массы однородных клеток с максимальными предосторожностями помещают в соответствующие среды для выделения и измельчают, например, с помощью миксера путем растирания или ультразвуком. Возникающий при этом гомогенат в идеальном случае больше не содержит целых клеток, однако еще могут сохраняться неповрежденные ядра, пластиды, митохондрии и т. д. Отдельные компоненты клеток можно теперь разчичным образом отделить от гомогената.
Рис. 2.8. Электронная микрофотография эмбриональной растительной клетки из апекса вегетативного побега цветной капусты (препарат получен путем замораживания скалывания) (препарат и электронная микрофотосъемка К. A. Platt-Aloia и W. W. Thomson с любезного разрешения J. Electron Мicг Techn John Wiley & Sons N. Y.)
Разлом замороженных клеток происходит местами вдоль мембран, которые проходят примерно параллельно плоскости скалывания такие мембраны оказываются здесь в рассматриваемой плоскости. Здесь, это имеет место в случае оболочки обоих ядер N с многочисленными ядерными порами. Митохондрии М и пропластиды РР частично расколоты частично видны как пластичный рельеф. Также, клеточные мембраны (плазматическая мембрана РМ) и тонопластные мембраны вакуолей местами представлены на поперечном сломе (срезе) в других местах по ним проходит слом Кроме того видны цистерны эндоплазматического ретикулума ER, а также диктиосома D. В клеточной стенке W местами различимы целлюлозные фибриллы (стрелки)

При дифференциальном центрифугировании гомогенат последовательно центрифугируют при разном числе оборотов (100 — 50 000 об/мин при большом числе оборотов ускорение может более чем в 100 000 раз превышать g — ускорение свободного падения на Земле). Фракционирование в этом случае осуществляется в основном по массе или размеру частиц. Сначала при малых оборотах (соответственно примерно 10 3 g в течение10 мин) из гомогената осаждаются ядра и пластиды Осадок посте сливания супернатанта ресуспедируют и считают более или менее «чистой» фракцией. Супернатант за тем вновь центрифугируют при более высоких оборотах, при этом осаждается следующая фракция — митохондрии (10 4 g в течение 30 мин) — и т д.
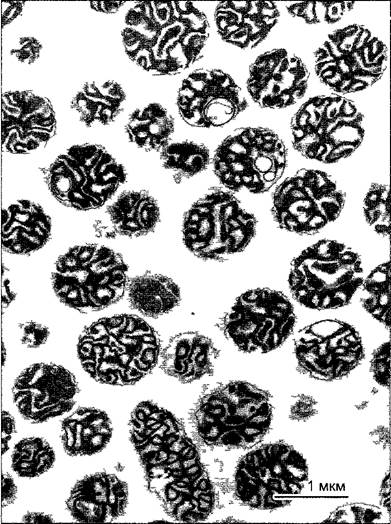
Рис. А. Фракция митохондрий, полученная путем изопикнического центрифугирования гомогената ткани шпината Матрикс митохондрий сморщен (ср рис 2 79), однако внутренняя дифференциация в значительной мере сохраняется Примесь других органелл клетки крайне незначительна (препарат В. Liedvogel, электронная микрофотография Н. Falk)
При центрифугировании в градиенте плотности плотность среды в центрифужных пробирках возрастает сверху вниз вследствие повышающихся концентраций сахарозы, С5СI 1 и т. п. В этом случае частицы гомогената разделяются по их плавучей плотности. Каждая частица располагается там независимо от размера и веса в градиентах, где плотность окружающей среды соответствует ее собственной (изопикническое, или равновесное, центрифугирование). Ультрацентр и фуга позволяет не только осуществлять сортировку внутриклеточных частиц, но и получать их характеристику по числу S (S — в честь Т. Сведберга (T. Svedberg), изобретателя ультрацентрифуги). Эти числа указывают дтя определенных частиц скорость осаждения (седиментации) при соответствующем ускорении центрифуги в единицах Сведбергa, 1S = 10 -13 с. У сферических частиц число Sпропорционально М 2,3 (М — масса частицы). Число S обычно наиболее широко используется для характеристики рибосом и их субъединиц (особенно частиц рибонуклеопротеидов) и белковых комплексов.
1 Хлористый цезий обладает высокой ионной силой, из-за которой нарушаются связи в белковых и нуклеопротеиновых комплексах. Поэтому его используют только для выделения индивидуальных веществ, но не органелл. — Примеч. ред.
Пузырьки, или везикулы (от лат. vesica — пузырь; уменьшительная форма vesícula) — мелкие, округлые элементы клетки; часто служат для накопления запасных веществ внутри клетки; возникают из более крупных элементов клетки путем отшнуровывания; особая форма: пузырьки диаметром только 0,1 мкм, с плотной оболочкой (coated vesicles). Транспорт путем перемещения потока пузырьков трактуется как совокупность процессов мембранных потоков (цитозы; англ, membrane flow).
Вакуоли (от лат. vácuus — пустой) — более крупные округлые компоненты, покрытые мембраной. Во взрослых растительных клетках часто образуется центральная вакуоль, которая часто занимает свыше 90 % объема клетки; содержит клеточный сок, чаще всего имеющий кислую реакцию; он отграничен от цитоплазмы мембраной вакуоли (топопластом; от греч. tonos — напряжение, давление, plástis — создание). Вакуоли часто содержат запасные вещества и вещества, подтежащие выделению, а также пигменты и токсины.
Пероксисомы — относительно крупные, плотные пузырьки (около 1 мкм в диаметре), которые наряду с другими веществами содержат фермент каталазу для разложения клеточного яда — перекиси водорода (Н2О2), возникающего в ходе процессов обмена веществ.
Олеосомы — капельки масла в цитоплазме (лат. oleum — масло); раньше их из-за округлой формы называли сферосомами.
Пластиды (англ, plastids) — во всех зеленых клетках водорослей, мхов и сосудистых растений это содержащие хлорофилл хлоропласты(chloroplasts; греч. chloros — желто-зеленый) — органеллы фотосинтеза. Преобразование энергии света (греч. phos — солнечный свет) в химическую энергию осуществляется на комплексных мембранных системах, которые образованы из содержащих хлорофилл мембранных цистерн (тилакоидов; греч. thylakos — мешок). Здесь возникает наряду с прочим аденозинтрифосфат (АТФ, англ. АТР — энергетическая «разменная монета» клетки; химическая энергия высвобождается за счет отщепления конечных фосфатных остатков и может использоваться для энергоемких реакций: синтезов, движения, активного транспорта на мембранах и т.д.; см. 6.1.5). В клетках тканей растений не зеленого цвета содержатся другие формы пластид, например, в образовательных тканях (меристемах) — мелкие, непигментированные пропластиды. В качестве хромопластов (от греч. chroma — цвет, окраска), окрашенных в цвета от желтого до красного, они служат в цветках и плодах для привлечения животных. Пластиды
всегда окружены оболочкой из двух мембран 1 и содержат собственную ДНК (плДНК = ctDNA) и рибосомы, которые отличаются от рибосом цитоплазмы (пластидные рибосомы). Размножение пластид осуществляется путем их деления — подобные возникают из подобных. Все формы пластид способны к образованию зерен крахмала и капелек масла (пластоглобул).
1 В дополнение к двум мембранам хлоропласта, пластиды могут иметь дополнительные мембраны: так, у Оchrophyta (Неterokontophyta) они окружены еще складкой эндоплазматической сети, так что фактически получается оболочка из четырех мембран — Примеч. пер.
Митохондрии (англ, mitochondria; от греч. mitos — нить, chóndros — зерно — из-за нитевидной или же короткоовальной формы), как и пластиды, имеют собственную ДНК (мтДНК — mtDNA) и рибосомы. Митохондрии возникают только из себе подобных путем деления. Окружены двойной мембраной. Это органеллы клеточного дыхания, при котором образуется АТФ. Образование АТФ и этапы клеточного дыхания осуществляются на внутренней мембране, площадь которой увеличена за счет впячиваний (крист, лат. crista — гребень) внутрь тела органеллы.
Клеточное ядро (Nucleus, Karyon; англ, cell nucleus; лат. nucleus; греч. káryon — ядро) — у большинства эуцитов (= эукариотных клеток) это самая крупная плазматическая органелла (около 10% объема клетки); чаще всего клетка имеет одно ядро. Ядро окружено двойной мембраной с характерными ядерными порами (англ, nuclear pores), а внутри него мембраны отсутствуют. Оно содержит большую часть наследственного материала клетки: генетическую информацию, закодированную в виде последовательностей оснований на длинных двойных спиралях ДНК. Молекулы ДНК представляют собой центральные структурные и функциональные элементы хромосом. В хроматине (веществе хромосом), наряду с ДНК в комплексе с основными белками — гистонами, находятся также в разных пропорциях негистонные белки. Ядро содержит одно или несколько ядрышек, где образуются предшественники цитоплазматических рибосом. Размножение ядер происходит путем деления (митоза). В нормальном случае исчезают ядерная оболочка и ядрышки, а хроматин из физиологически активной, деконденсированной «рабочей формы» путем конденсации отдельных хромосом переходит в «транспортную форму». (Для сильно окрашивающихся структур, имеющих форму от палочковидной до нитевидной, которые представляют собой конденсированный хроматин во время митоза, был исходно предложен термин «хромосомы».) Хромосомы благодаря аппарату веретена деления (англ, mitotic spindle: цитоскелетная структура, состоящая прежде всего из микротрубочек) поровну распределяются по дочерним клеткам, в которых затем заново образуются ядерные оболочки и ядрышки и происходит деконденсация хроматина по меньшей мере «эухроматических» компонентов, тогда как гетерохроматин остается конденсированным и неактивным. В эухроматине синтезируется РНК на определенных последовательностях ДНК (транскрипция активных генов). Закономерно следующие друг за другом, повторяющиеся от деления к делению, стадии составляют клеточный цикл (англ, cell cycle). В S-фазе клеточного цикла происходит репликация ДНК и за счет нее удваивается число хромосом.
Читайте также: Декор своими руками для дома из ткани своими руками
При процессах дифференцировки клетки изменяются, однако многие органеллы сохраняют свои облики функции. Только пластиды, вакуоли и клеточные стенки в целом изменяются сильнее,
В общем, ультраструктура клетки у высших растений варьирует значительно меньше, чем у водорослей и грибов, которые филогенетически древнее и родство между ними еще меньше, чем у различных по облику мохообразных, папоротникообразных и семенных растений.
Термин «протоплазма» 1 означает вязкую или студенистую массу, в которой расположены рибосомы и элементы цитоскелета и в которую погружены пластиды и митохондрии, клеточное ядро и часто также скопления запасных веществ (олеосомы, у грибов гранулы гликогена). Она богата ферментами. Общая концентрация белков составляет от 10 до 30 %. В цитоплазме значительная часть воды находится в связанном с белками состоянии. Белки протоплазмы преимущественно глобулярные. За счет активных, т. е использующих АТФ, ионных насосов на граничащих мембранах в цитоплазме поддерживается определенная ионная среда. По сравнению с внешней средой цитоплазма богата К + , бедна Na + и содержит незначительное количество Са 2+ ; pH составляет немногим более 7, что для ферментов цитоплазмы оптимально
1 Термины «протоплазма» и «цитоплазма» часто употребляются как синонимы. — Примеч. ред.
В протоплазме протекают многие важные реакции и осуществляются обменные процессы (гликолиз, образование запасных липидов, синтез аминокислот и — на рибосомах — биосинтез белка, а кроме того, синтез нуклеотидов и сахарозы, см 6.10 — 6.17). В протоплазме клеток многих растений и грибов образуются вещества, используемые в фармакологии (алкалоиды, гликозиды); они откладываются в вакуолях или клеточных стенках и там накапливаются. У грибов (и животных) в цитоплазме синтезируются жирные кислоты; у растений этот синтез локализован в пластидах.
Протоплазма может существовать в форме золя и геля. Сгущающее действие оказывают элементы цитоскелета. Растворы глобулярных макромолекул (таковыми являются коллоиды из сферических частиц — ферментные белки протоплазмы) даже при высоких концентрациях сохраняют низкую вязкость 1 . Напротив, вытянутые в длину частицы (линейные коллоиды) уже при низких концентрациях образуют студенистую массу 2 . Из-за большой поверхности они склонны к уплотнению. Актиновые нити и микротрубочки образуют явные линейные коллоиды, однако одновременно они являются агрегатами глобулярных белков В живой клетке эти агрегаты могут быстро собираться и разбираться, так что вязкость цитоплазмы может в короткий срок изменяться в соответствии с потребностями. Поскольку растительные и грибные клетки имеют жесткий внешний скелет в виде клеточной стенки, жидкая цитоплазма встречается у них значительно чаще, чем в клетках животных или голых клетках многих жгутиковых и низших грибов. Во всех клетках части цитоплазмы, преимущественно расположенные по периферии, — эктоплазма — кортикальная цитоплазма (лат cortex — кора) — представлены в виде геля, тогда как расположенная в середине клетки эндоплазма часто жидкая. Заметный ток характерен только для эндоплазмы
1 Именно такие коллоидные растворы называют золями — Примеч. ред.
2 Те образуют гели — Причеч. ред.
Интенсивный ток цитоплазмы наблюдается прежде всего в особенно крупных клетках. Он служит для быстрого внутриклеточного транспорта веществ, для которого недостаточно одной диффузии. У имеющих стенку клеток различают ротационный и циркуляционный токи. В случае ротации цитоплазмы эндоплазма, окружающая центральную вакуоль, постоянно однообразно перемещается вокруг нее, просто циркулируя или двигаясь восьмерками. Этот вид тока цитоплазмы наблюдается в чрезвычайно крупных клетках междоузлий водорослей Chara и Nimlla (см рис 5 9), а также в клетках листьев широко известных аквариумных растений Elodea и Vallisneria. В клетках с верхушечным ростом (гифы грибов, корневые волоски, пыльцевые трубки), в клетках волосков (например, в жгучих волосках крапивы) и многих эпидерматьных клетках ток цитоплазмы осуществляется в виде многочисленных, иногда встречных течений, преимущественно также в плазматических тяжах, которые пронизывают центральную вакуоль (см рис 2 59, С) (о физиологии внутриклеточных движений см в разделе 8 2 2)
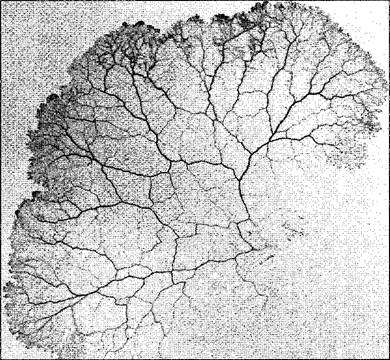
Амебоидное движение клеток, не имеющих стенки или же плазмодиев, также основывается на плазматических токах Рекорд скорости — 1 мм/с — достигается при так называемом маятникообразном токе в сетевидном плазмодии слизевиков (рис 2.9) Он происходит оттого, что сократительный эктоплазматический слой в некоторых местах жилок, составляющих сеть плазмодия, сжимается, а в других расширяется Направление тока меняется на противоположное каждые 2,5 мин. Не только здесь, но и при токах, на порядок более медленных циркуляционных и ротационных, движущая сила обусловлена актомиозиновой системой (см. 2.2.2.2). В противоположность маятникообразному току у миксомицетов, здесь возникают не гидравлические, а силы деформации сдвига, которые смещают эндоплазму относительно неподвижной эктоплазмы.
Рис. 2.9. Плазмодий слизевика Рhysarum confertum (1,25х) (фотография R. Stiemerling)
Лишенные стенки клетки (протопласты) 1 под влиянием сил поверхностного натяжения, действующих у плазматической мембраны, имеют тенденцию к приобретению круглой формы, к сокращению своей поверхности до минимума. Искусственно лишенные стенки растительные, грибные и бактериальные клетки соответственно принимают круглую форму (ср. рис. 2.48). Отклонения от круглой формы возможны за счет выступающих жестких структур вне клеточной мембраны (дерматобластов 2 , с клеточной стенкой) и/илицитоскелета в самой цитоплазме. Цитоскелет, разумеемся, развит особенно сильно у клеток, лишенных стенки: яйцеклеток и клеток тканей животных и человека. В амебоидных, многоядерных плазмодиях слизевиков (рис. 2.9) цитоскелетный белок актин составляет примерно 1 /5 всего белка. Если клеточные стенки в нормальном случае могут лишь медленно изменяться в ходе необратимого роста и только в исключительных случаях разрушаться, цитоскелет может быстро воссоздаваться и деградировать. Он проявляет себя как динамичный образователь структуры, который определяет облик клеток, внутреннюю архитектуру и все происходящие в них движения, и в конечном счете, морфогенез органов.
1 В исходном тексте был использован термин, встречающийся в немецкой литературе, —- гимнобласты. В переводе он заменен на более распространенный. — Примеч. ред.
2 Русского эквивалента этому термину найти не удалось. — Примеч. ред.
Цитоскелет водорослей, высших растений и грибов составляют прежде всего актиновые микрофиламенты и микротрубочки (рис. 2.10).
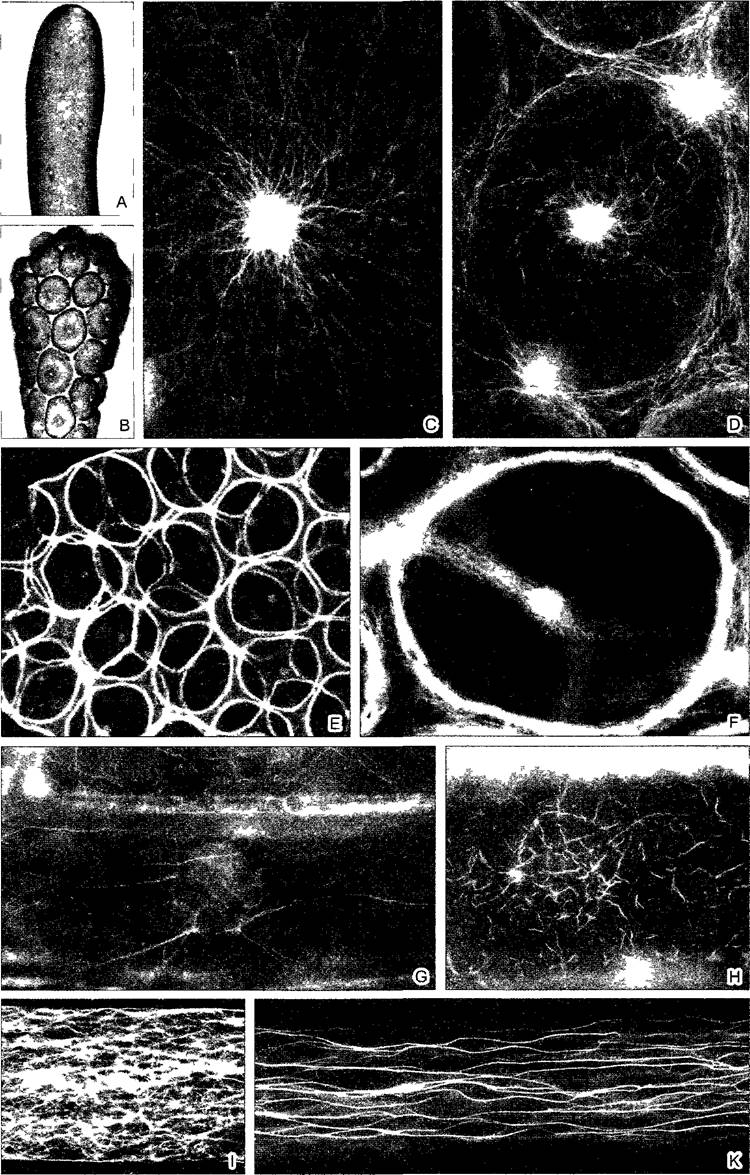
Рис. 2.10. Цитоскелет в растительных клетках (препараты и фотография: A—F — D. Menzel: G, Н — Н.Quader; I, К — М. Braun и A.Sievers): А—F — образование цист у дазикладовой водоросли Acetabularia cliftoni (ср. рис. 11.92). А, В — переход вторичных ядер в клетку «зонтика» водоросли и начинающееся формирование цист путем свободного образования клеток (см. 2.2.3.6); С, D — микротрубочки становятся видимыми благодаря применению непрямой иммунофлуоресценции; в середине картины во всех случаях клеточное ядро (С— х350; D— х235); Е, F — соответствующая локализация актиновых микрофиламентов при образовании цист; при более сильном увеличении (F) видны отдельные нити (Е— 60х; F— 235х); G—актиновые микрофиламенты в клетках эпидермиса чешуи лука (в середине клеточное ядро), видимые при применении флуоресцентной микроскопии благодаря образованию комплекса с фаллоидином — компонентом токсина гриба бледной поганки. Циклический полипептид фаллоидин с высокой специфичностью присоединяется к актину; он связывается с красителем родамином, который в ультрафиолетовом свете дает красное свечение; Н — разрушение микрофиламентов цитохалазином (G. Н— 400х); I, К — актиновые микрофиламенты в ризоидах харовой водоросли Chara globularis (ср. рис. 11.106) после флуоресцентной окраски родаминфаллоидином, позади кончика ризоида микрофиламенты сетевидные, в базальной зоне — в виде более густых пучков (1000х)
Актин был первоначально выделен из мышечных волокон. Позже было установлено его наличие во всех эукариотических клетках. Глобулярная молекула актина (G- актин) имеет диаметр 40 нм и массу 42 кДа. В области соединения более крупного С- концевого и меньшего N-концевого домена локализован центр связывания для АТФ. В растворах G-актина легко образуются актиновые нити — микрофиламенты (F-актин; рис. 2.11). При этом АТФ G-актина расщепляется, АДФ остается связанной с протомерами F-актина. Гидролиз АТФ не является обязательной предпосылкой для образования нитей, но за счет стабилизации нитей оказывает аллостерический эффект.
Рис. 2.11. Актиновый микрофиламент.
Глобулярные (точнее; эллипсоидные) актиновые мономеры группируются в спирали примерно с двумя молекулами на оборот. Это придает микрофиламенту вид растянутой двойной спирали с шагом примерно 40 нм
Микрофиламенты обладают кинетической полярностью. Встраивание дальнейших актиновых молекул осуществляется преимущественно у так называемого плюс-конца. Образование нитей начинается в живой клетке в особых местах нуклеации, предпочтительно в определенных местах клеточной мембраны, которые заняты образующими актин белками (например, α-актинином). Поразительным образом на этих центрах образования находится плюс-конец нити. Рост микрофиламентов осуществляется, таким образом, путем удлинения у фиксированного, а не у свободного конца. (Микротрубочки ведут себя в этом плане противоположным образом.)
На скорость и масштаб роста микрофиламентов, на их положение и ориентацию могут влиять также многие природные и искусственные факторы. В живой клетке микрофиламенты стабилизируются или же дестабилизируются. группируются в сети или пучки, испытывают подавление дальнейшего роста или перемежаются ассоциированными с актином белками. На действии этих белков — в наиболее хорошо изученных клетках млекопитающих их имеется свыше 100 типов — основывается высокая динамика актинового цитоскелета. В растениях при этом встречаются два белка, действующих как антагонисты; профилин и деполимеризующий актин фактор (ADF). Они всегда накапливаются в местах локальных структурных преобразований на поверхности клеток как при усиленном верхушечном росте (корневые волоски, пыльцевые трубки), так и при клеточном делении (образование срединной пластинки и первичных плазмодесм, см. 2.2.3.6 или 2.2.7.3). В эксперименте цитохалазин В — яд грибного происхождения — разрушает микрофиламенты. В результате блокируются внутриклеточные процессы движения, в которых участвуют микрофиламенты. Это относится, например, к токам цитоплазмы и перемещению хлоропластов. Такой же эффект вызывает фаллоидин — яд бледной поганки (Amanita phalloïdes). Правда, под его влиянием, наоборот, происходит агрегация всего клеточного актина в нити, которые больше не могут разделяться («Ph-актин»), и таким образом подавляет жизненно важную динамику цитоскелета. Актин является одним из наиболее консервативных белков эукариот: последовательность его аминокислот почти не изменилась на протяжении всего времени эволюции. Однако в геноме большинства эукариот имеется несколько генов актинов (например. 10 — у Агаbidopsis), продукты которых не полностью идентичны. Речь идет об изотипах, или изовариантах. В клетках, не относящихся к мышцам, обычно синтезируется γ-изотип актина.
Молекулярный компонент микротрубочек (рис. 2.12) — димерная единица из двух сходных, но не идентичных белков: α- и β-тубулина («гетеродимер»). Гетеродимеры тубулина (100 кДа) показывают при наличии ГТФ и в отсутствие ионов кальция сильно выраженную тенденцию к агрегации. Типичная структура их самосборки — микротрубочка (microtubulus; лат. tubulus — трубочка). Стенка микротрубочки состоит в целом из 13 продольных рядов (протофиламентов) из одинаково ориентированных тубулиновых гетеродимеров. Внешний диаметр третичной структуры микротрубочки составляет порядка 25 нм, тогда как диаметр микрофиламента актина — только 6 нм. Поэтому, микротрубочки — сравнительно жесткие вытянутые образования В случае чрезмерных изгибов (чего, правда, в нормальной клетке не происходит) они ломаются.
Рис. 2.12. Тубулин и микротрубочки (электронная микрофотосъемка В — I Dorr, С —N FaJk)
А—гетеродимеры из глобулярного α- и β-тубулина (показаны соответственно темным и светлым, каждый примерно 50 кДа 4 нм в диаметре) организованы в продольные ряды — протофиламенты, 13 протофиламентов образу ют полую цилиндрическую микротрубочку. Гетеродимеры соседних протофиламентов при этом слабо сдвинуты друг относительно друга так что возникает плоская спиральная структура. У так называемой каймы (на схеме спереди) единицы тубулина располагаются не так, как в остальных местах (a-единица рядом с α-единицей β-единица рядом с β-единицей), а α- единица находится рядом с β-единицей, В—микротрубочки банана Musa paradisiaca в негативном контрасте, С—поперечный разрез микротрубочки препрофазного пучка (см рис 2 23 В) в зародышевой клетке корневого чехлика лука, местами различимы 13 протофиламентов

Молекулярные структуры α- и β-тубулина очень сходны, хотя последовательности аминокислот совпадают только на 40 % К удивлению, можно установить их гомологию с белком клеточного лечения FtsZ у бактерий Каждая молекула тубу- лина обладает местом связывания с ГДФ/ ГТФ. Свободные гетеродимеры тубулина, в изобилии встречающиеся в большинстве клеток, связывают ГТФ, а после агрегации — ГДФ, что соответствует аналогичным взаимоотношениям в случае G-/F- актина относительно соединения АТФ/ АДФ.
Места образования «затравки» для микротрубочек в клетке называют центрами организации микротрубочек, или ЦОМТ (англ, microtubuli organizing center, МТОС).
В качестве таковых выступают прежде всего базальные тельца жгутиков, они соответствуют центриолям (см 2 2 2 3), а те и другие — полярным областям веретена ядерного деления (бокс 2 2) и, кроме того, определенным участкам мембран Как и микрофиламенты, каждая микротрубочка обладает плюс- и минус-концом, что выражается в одинаковой ориентации тубулиновых тетеродимеров по всей длине каждой микротрубочки Однако в противоположность микрофиламентам у микротрубочек минус-конец прикрепляется к ЦОМТ, а плюс-конец отходит оттуда в сторону Места образования новых микротрубочек на ЦОМТ, имеющие форму коротких левовращающих спиралей (места образования «затравки»), состоят из нескольких специфических белков и содержат третий изотип тубулина — у-тубулин. Последний взаимодействует с β-тубулином Минус-конец микротрубочек является их β-концом, тогда как плюс-конец соответствует α-концу. У высших растений у- тубулин ассоциирован не только с местами нуклеации, так называемыми кольцевыми комплексами у-тубулина; скорее он встречается вдоль целых микротрубочек, а также на эндомембранах.
Читайте также: Чем отмыть ткань от краски для волос
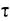
Скорость и масштаб удлинения микротрубочек зависят не только от наличия тубулиновых димеров и ГТФ, но и от ряда других факторов и могут ими регулироваться. Так, агрегация тубулина происходит только при концентрациях кальция -7 М. В живой клетке важную роль играют различные белковые факторы. Они в совокупности обозначаются как ассоциированные с микротрубочками белки (microtubuli associated proteins, МАР). (Внимание! Аббревиатура МАР часто применяется также для «активирующих митоз белков — mitosis activating proteins»; недоразумений можно избежать, только учитывая контекст.) Имеется два класса таких белков:-фактор (тау-фактор, 55 — 65 кДа), который встраивается в микротрубочки, и высокомолекулярные МАР (250 — 350 кДа), которые обычно отходят от микротрубочек как боковые «руки» длиной до 30 нм и могут функционировать как мостики между ними и, например, мембранами. Некоторые из высокомолекулярных МАР — ферменты: они могут,
например, фосфорилировать белки или являются АТФазами, из них важнейшие — динеин и кинезин (см. следующий раздел).
Как и в случае микрофиламентов, на сборку и разборку микротрубочек могут влиять в эксперименте специфические вещества. Давно известен колхицин — алкалоид безвременника осеннего (Colchicum autumnale). Он присоединяется к ß-тубулину свободных тубулиновых гетеродимеров и блокируем их встраивание в микротрубочки. В настоящее время для экспериментальной разборки микротрубочек применяются более сильно и специфично действующие гербициды — оризалин и амипрофосметил (АПМ). Противоположное действие оказывает таксол — алкалоид тиса (Taxus; см. рис. 6.123). Он стабилизирует микротрубочки и стимулирует агрегацию свободных гетеродимеров.
Стабильность микротрубочек одной и той же клетки зачастую не одинакова: различают «стабильные» и «лабильные» микротрубочки. Под воздействием колхицина дезинтеграции подвергаются лабильные (например, микротрубочки веретена ядерного деления), но не стабильные (в жгутиках) микротрубочки. Микротрубочки жгутиков сохраняются даже при низких температурах и фиксации тетроксидом (четырехокисью) осмия, тогда как лабильные микротрубочки исчезают в обоих случаях. Широкое распространение микротрубочек лабильного типа удалось установить только после внедрения в практику электронной микроскопии фиксации глутаральдегилом. В основе различной стабильности микротрубочек лежит, возможно, существование различных изотипов тубулина (у Arabidopsis, например, известны девять α-тубулинов и шесть β-тубулинов) и/или специфические сопутствующие белки.
Во многих клетках встречаются комплексные структурные образования из микротрубочек — иногда они временные, для ограниченных во времени функций, в других случаях это длительно существующие образования.
Наиболее известный пример таких функциональных структур — веретено деления ядра (см. бокс 2.2). Однако не только при митозе, но и во время других фаз клеточного цикла в клетках высших растений часто возникают характерные группировки микротрубочек — цикл микротрубочек (рис. 2.13). В интерфазе микротрубочки преимущественно локализованы непосредственно под клеточной мембраной в кортикальной цитоплазме. Они играют там важную роль при образовании клеточной стенки (ориентация целлюлозных микрофибрилл; локальное отхождение клеточной мембраны от клеточной стенки для ограниченных в пространстве вторичных образований стенки, как при дифференцировке спиральных сосудов в ксилеме; ср. рис. 2.74, С; 3.24, Е) и при морфогенетических процессах.
Рис. 2.13. Изменения расположения микротрубочек перед началом митоза в клетках корневой меристемы (по М. С. Ledbetter): А—интерфаза, В—образование препрофазного пучка перед вступлением в профазу, его положение определяют последующий экватор и плоскость деления клетки, С—поздняя профаза

Хорошо выраженные стабильные микротрубочки распространены у лишенных покровов протистов и сперматозоидов, где они связаны с выработкой характерных жестких клеточных форм и/или закреплением жгутикового аппарата.
В клетках позвоночных животных наряду с актиновыми и миозиновыми нитями и микротрубочками встречаются также другие элементы цитоскелета в виде нитей диаметром порядка 10 нм. Таким образом, по размеру они находятся между микротрубочками (25 нм) и микрофиламентами (6 нм) и поэтому получили название промежуточные филаменты (intermediate filaments — IF = 10-нм-филаменты). К тому же в цитоплазме млекопитающих 1F часто образуют вытянутые в длину, плотные волокнистые структуры. Они отличаются тем, что нерастворимы — за исключением концентрированного раствора мочевины. Промежуточные филаменты представляют собой пучки вытянутых в длину белковых молекул. К настоящему времени известно 6 подсемейств; примерно 40 различных 1F-белков гомологичны по своим последовательностям. (Сюда относятся также ламины — белки ядерной пластинки; см. 2.2.3.4.) На основании данных по последовательностям ДНК гены 1F-белков выявлены и у низших животных. Растительные клетки также содержат белки, которые взаимодействуют с антителами к 1F-белкам животных. Для некоторых из этих белков установлена сильно выраженная тенденция к агрегации со структурами типа промежуточных филаментов.
2.2.2.2. Моторные белки и процессы движения в клетке
Цитоскелет принимает решающее участие в процессах движения в клетке (сократимость, подвижность). С одной стороны, движения направлены как по рельсам поезда или трамвая. С другой стороны, по закону Ньютона (действие численно равно противодействию) — каждый действующий элемент нуждается в противодействующем (например, мускулатура и скелет). В клетке специфические АТФазы, функционируя как хемомеханические преобразователи энергии (моторные молекулы), поставляют энергию, высвобождающуюся при расщеплении АТФ, для структурных изменений и таким образом непосредственно взаимодействуют с элементами цитоскелета. У эукариот соответственно двум основным компонентам цитоскелета широко распространены две подобные системы: актомиозиновая система и система микротрубочки—дине- ин/кинезин.
Партнером актина при создании сил растяжения и сдвига является миозин — комплексная АТФаза (греч. myon — мышца), которая активируется актином. Миозин был особенно детально изучен на мышцах позвоночных животных и насекомых. Имеющийся у них в большом количестве миозин II обладает четвертичной структурой (470 кДа) из 2 параллельных длинных («тяжелых») и 4 более коротких («легких») цепей (рис. 2.14, А, В). Сильно анизометрическая элементарная часть миозинового комплекса имеет α-спиральную область («хвост») и два N-концевых глобулярных участка («головки»). Последние идентичны и функционируют при сократительных процессах независимо друг от друга. В них локализована активность АТФазы и они же являются местом связывания актина; связывающие Са 2+ легкие цепи находятся по соседству. Над хвостовыми доменами миозин II агрегируется в поперечнополосатых мышечных волокнах в заметные стабильные миозиновые нити. При достаточно высокой (примерно мил- лимолярной) концентрации Са 2+ одна из двух миозиновых головок ложится на актиновую нить, и продукты уже осуществившегося расщепления АТФ высвобождаются. Это приводит к радикальному изменению структуры миозина: головка изгибается и сдвигает микрофиламент примерно на 10 нм, за счет нового накопления АТФ связь с актином разрывается, и головка при расщеплении АТФ снова выпрямляется. При мышечном сокращении эти процессы циклически повторяются, происходит взаимное перемещение миозина и актина. Двухкомпонентная система из нитей актина и миозина действует, таким образом, не столько за счет укорочения самих нитей, сколько благодаря смещению друг относительно друга. Модель скользящих волокон (модель «slidingfilament») подходит в основных чертах также для системы микротрубочки— динеин/кинезин.
Рис. 2.14. Моторные молекулы (В—Е — по D. Menzel): А—миозин II из мышцы млекопитающих — наиболее давно известная и лучше всего изученная моторная молекула. Она расщепляется про- теиназой трипсином (Т) на головную и хвостовую части. Папаин (Р) разлагает головную часть на N-концевые глобулярные домены тяжелых миозиновых цепей с местом соединения с актином и АТФазной активности и шейковый участок, с которым ассоциированы легкие цепи (здесь не изображены). Шейковый и хвостовой участки представляют собой вытянутые α-спиральные домены. Места воздействия Т и Р находятся в местах ослабления вторичной структуры, у которых миозиновые субъединицы могут надламываться, как в суставах. На изгибах «сустава» возле Р основывается взаимное смещение актиновых и миозиновых филаментов; В—структура домена миозина II. Глобулярные единицы на шейковых участках соответствуют (как и на С—Е) легким цепям; С—Е —миозины из растительных клеток: С, D — миозины соответственно классов XI и VIII подсолнечника и ре- зуховидки (Arabidopsis thaliana)’, Е—миозин класса XII из сифональной зеленой водоросли Acetabularia; F—кинезин; обе α-цепи образуют у своих N-концов (справа) глобулярные домены с АТФазной активностью и местом присоединения тубулина; две легкие цепи расположены у С-концов. Миозины класса II имеют длину 160 нм, тетрамеры кинезина — только примерно половину этой длины
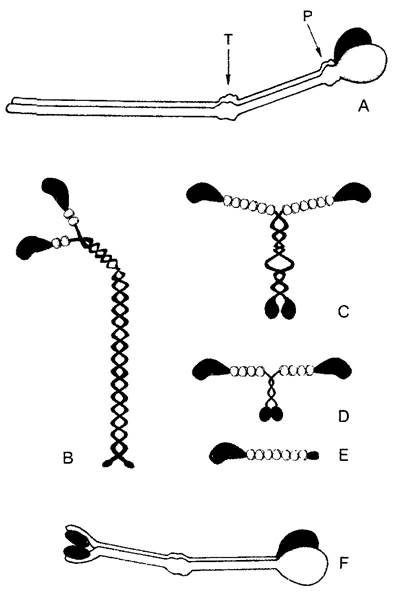
Путем сравнения ДНК-последовательностей генов и исследования подвижности (белковые фракции тестируют на способность вызывать движения у изолированных нитей актина) в последние годы были обнаружены многочисленные миозины, причем некоторые из них существенно отличаются от миозина II из мышечных клеток. Это касается прежде всего растительных миозинов, которые были отнесены к классам VIII, XI и XII надсемейства миозинов (см рис 2 14 С —Е). По характер) действия они сходны с миозином II однако их хвостовые области не образуют миозиновых нитеи (они имеются только в мышечных клетках), а прикрепляются непосредственно к мембранам, пузырькам или другим клеточным структурам за исключением микротрубочек). Токи цитоптазмы и перемещения хлоропластов вызываются в подавляющем большинстве случаев актомиозиновои системой.
В противоположность актомиозиновои системе при движениях, обусловленных микротрубочками, задействованы два разных класса моторных белков динеины и кинезины (греч dynamis — сила, kinesis — движение). Динеин — высокомолекулярный комплекс — выявляется прежде всего в жгутиках и ресничках (см следующий раздел), где он и был открыт. Вместе с тем, широко распространен «цитоплазматический динеин» более простого строения — он имеется и в клетках, лишенных жгутиков. Динеин действует совместно с динактином — другим крупным белком динаминового комплекса, а также с другими, от случая к случаю меняющимися сопутствующими белками Сообщаемые динеином движения происходят всегда в направлении к минус-концу микротрубочек, функционирующих как упор/рельсовый путь динеины представляют собой минус-моторы Кинезины, напротив, — в большинстве плюс-моторы (Лишь немногие представители надсемейства кинезинов, отличающиеся по своему строению — у них моторные домены локализованы на карбоксильном, а не на N-конце, — действуют как минус-моторы ). Они были открыты на осевых придатках нервных клеток. Между тем, достоверно подтверждено их наличие и у растений. По молекулярному строению (см рис 2 14, F) они сходны с миозинами, которые также (только именно на актиновых микрофиламентах) являются плюс-моторами. Хотя кинезины и не гомологичны миозинам по последовательности аминокислот, трехмерная структура моторных доменов в обоих случаях сходна.
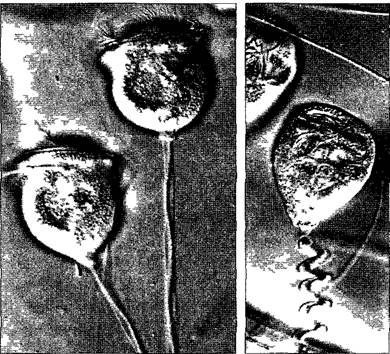
Клеточные движения осуществляются в некоторых случаях без участия описанных систем. Так, простое удлинение или же укорачивание микрофиламентов или микротрубочек может вызывать перемещение органелл или изменение их формы. Механизм движения основан на совершенно иных молекулярных взаимодействиях. Например, ножка одноклеточных сувоек (сидячие инфузории) сокращается при прикосновении к клетке (телу животного) При этом центральный тяж в ножке — спазмонема — скручивается в спираль с малым шагом (рис 2 15). Спазмонема состоит в основном из низкомолекузярного фосфопротеина (≈20 кДа), который относится к семейству центринов (- спазминов = кальтрактинов) Он резко изменяет свою пространственную структуру (и тем самым облик спазмонемы) под воздействием ионов кальция АТФ при этом не расщепляется. Однако необходим сравнительно длительный процесс, чтобы посте сокращения удалить Са 2+ и снова вытянуть спазмонему. Центрины по постедоватетьности аминокислот близко родственны белку кальмодулину, связывающему кальций. С помощью специфических антитез центрин удалось выявить у очень многих эукариот, даже у высших растений. Он ассоциирован прежде всего с базальными тельцами и центриолями, а также со структурами прикрепления жгутиков. Центрин встречается при клеточных делениях также в центральной части цитоптазмы и во фрагмопластах (см. 2 2 3 6), но его функции здесь пока неизвестны.
Рис. 2.15. Сократительная спазмонема сувойки Vorticella (слева — в вытянутой ножке справа — после раздражения) спазмонема и ножка спиралевидно укорочены (420х, интерференционный контраст)
Постоянные жгутики эукариот (= flagellа; лат. flagellum — бич, жгут) по структуре в основных чертах сходны. Вообще, здесь мы имеем дело с чрезвычайно консервативными клеточными структурами.
Даже широко распространенные у животных и человека реснички показывают, в основном, сходную ультраструктуру. Реснички короче, чем жгутики, и всегда многочисленны на несущих их клетках (клетки ресничного эпителия; одноклеточные организмы — инфузории). Для жгутиков и ресничек эукариот стал популярным (как обобщающее название) термин ундулиподии (лат. undulare — извиваться, podium — придаток в виде ножки) Аналогичные органеллы передвижения бактерий построены по иным принципам и функционируют совершенно другим образом (см. 2.3.2).
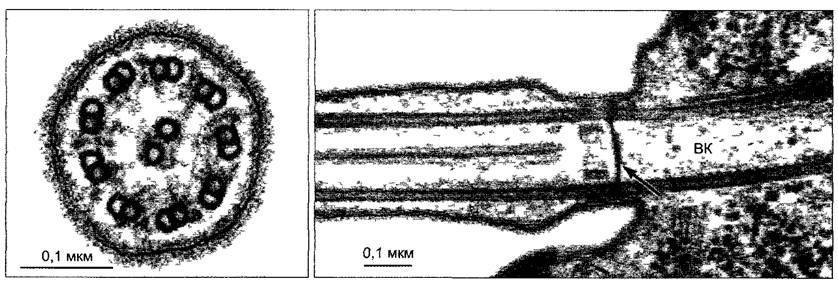
На поперечном срезе жгутика можно различить характерное расположение 20 микротрубочек (рис. 2.16), известное как модель 9 + 2. Две центральные одиночные микротрубочки (синглеты) симметрично окружены венцом из 9 двойных жгутиков (дуплетов). Дуплеты ориентированы не строго тангентально: так называемая А-
Читайте также: Игрушка сердечко своими руками из ткани
трубочка находится несколько ближе к центру, чем В-трубочка. Только А-трубочка построена из 13 протофиламентов. В-трубочка, имеющая больший диаметр и включающая 11 протофиламентов, расположена сбоку от А-трубочки и использует 4 из ее протофиламентов. За счет этого она имеет вид также замкнутой (правда, не совсем округлой) трубки. Синглеты и дуплеты вместе с многочисленными другими белками образуют сложный цитоскелет жгутиков (рис. 2.17). Вместе с динеином в качестве моторной молекулы он осуществляет биение жгутика. Подвижная общая структура, которая проходит по длине жгутика и имеет диаметр 200 нм, носит название аксонема (от греч. axon — ось, nema — нить).
Рис. 2.16. Жгутик Scourfieldia caeca — зеленой жгутиковой водоросли (электронная микрофотосъемка М.Меlkoniai).
Слева — поперечный разрез, справа — основание жгутика с базальным телом на продольном разрезе. Центральные синглеты, которые отсутствуют в базальном теле, начинаются только на расстоянии 100 нм кнаружи от этой пластинки. На поперечном срезе не очень четко видны динеиновые рукоятки и радиальные спицы; ВК— базальное тело, стрелка—базальная пластинка у перехода базального тела в жгутик
Динеин жгутиков представляет собой очень сложную АТФазу, активируемую тубулином. Динеин так называемых «рукояток» имеет, например, массу, едва ли достигающую 2 МДа и состоит из примерно 12 различных протомеров. В ЭМ он выглядит как многочленная структура. Динеиновые рукоятки, которые отходят от А-трубочек и направлены к соседним В-трубочкам, могут передвигать соседние дуплеты друг относительно друга — здесь также работает модель скользящих волокон (см. рис. 6.6). Радиальные «спицы» и нексиновые мостики преобразуют получающиеся в результате продольные сдвиги в характерные изгибы жгутика.
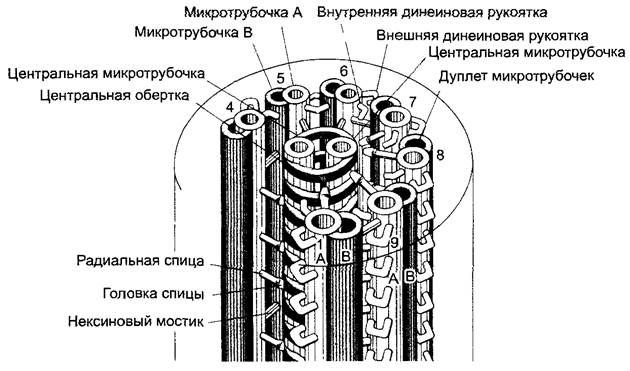
Рис. 2.17. Схема ультраструктуры эукариотического жгутика (по Р.Эайг)
Обе центральные микротрубочки (синглеты) окружены спиральным влагалищем, с которым периферические дуплеты связаны через эластичные радиальные спицы. В каждом дуплете А-микротрубочка (показана светлым) непрочно соединена с В-микротрубочкой (показана темным) соседнего дуплета эластичной белковой «рукой» (нексин). Каждая А-микротрубочка, кроме того, несет внутренние и внешние динеиновые рукоятки. Отсчет дуплетов начинается от плоскости симметрии синглетов с 1 и продолжается в направлении динеиновых рукояток (при рассмотрении в направлении от основания к свободному концу жгутика — почасовой стрелке). Для большей наглядности показано только 7 дуплетов; отсутствие дуплетов 2 и 3 отражено в прерывании круга, который отмечает положение плазматической мембраны
Поверхность жгутиков у разных организмов может различаться. Перистые жгутики снабжены расположенными по их бокам нитевидными мастигонемами (от греч. mastix — жгутик), за счет чего сильно увеличивается трение о воду (см. рис. 11.20, А—С; 11.72, Г). Мастигонемы образуются в аппарате Гольджи как оформленный секрет и доставляются путем направленного экзопитоза на поверхность жгутика. Гладкие жгутики отличаются удлиненной тонкой верхушечной зоной, в которую заходят только синглетные микротрубочки.
Каждый жгутик прикрепляется в периферической цитоплазме базальным телом — коротким цилиндром из 9 триплетов микротрубочек (трубочки А, В, С); центральные синглеты отсутствуют (см. рис. 2.16; 2.18). Базальное тело ориентировано перпендикулярно к поверхности клетки. При возникновении жгутиков оно функционирует как центр образования, от которого вырастает жгутик. В переходной зоне между базальным телом и телом жгутика С-трубочки заканчиваются и берут начало обе синглетные трубочки. Трубочки А и В базального тела продолжаются в 9 дуплетах аксонемы. Базальные тела имеют также функцию ЦОМТ, а плюс-концы микротрубочек жгутиков находятся на свободном конце жгутика.
Структура базальных тел идентична структуре центриолей. Они встречаются в основном попарно. В клетках, имеющих базальные тела, чаще всего они функционируют как пары центриолей, но, что примечательно, центриоли не всегда служат полюсами веретена деления.
В свое время ошибочно предполагали, что центриоли являются центрами образования аппарата веретена. Однако имеются организмы и без центриолей, например, все покрытосеменные 1 , которые тем не менее вполне способны образовывать веретено деления при митозе, но не образуют жгутиков (и поэтому также не имеют сперматозоидов, так что сближение ядер гамет у этих растений осуществляется за счет быстро растущей пыльцевой трубки — сифоногамия). Таким образом, центриоли — это потенциальные базальные тела, которые при необходимости могут перемещаться к поверхности клетки и там разрастаться в жгутики. Во время митоза они в первую очередь занимают полюса веретена, за счет чего обеспечивается их равномерное распределение по дочерним клеткам.
1 Кроме того, без центриолей обходятся клетки красных водорослей (Rhodophyta), часть зеленых водорослей (Соnjugatae) и часть голосеменных. Клетки многих грибов также лишены центриолей. — Примеч. ред.
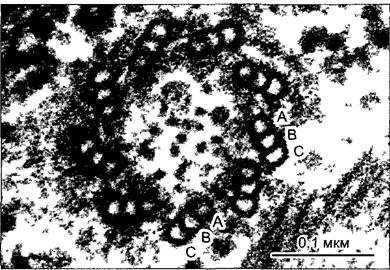
Рис. 2.18. Базальное тельце Scourfieldia, поперечный разрез (электронная микрофотосъемка М.Меlkonian).
В триплетах микротрубочек местами различимы протофиламенты на поперечном срезе. Только самые внутренние микротрубочки триплета (А) комплектны; обе косо направленные наружу микротрубочки В и С желобчатые, они имеют несколько протофиламентов, общих с соседней микротрубочкой, расположенной ближе к середине. С-микротрубочки заканчиваются у базальной пластинки, А и В продолжаются в дуплетах аксонемы жгутика
Базальные тела (и соответственно центриоли) возникают не путем деления из подобных себе, а каждый раз образуются заново. Правда, часто это происходит в непосредственной близости от базальных тел/центриолей, которые, очевидно, оказывают индуцирующее действие. Базальные тела высокоразвитых папоротникообразных и голосеменных 1 , которые еще образуют сперматозоиды (в некоторых случаях со жгутиками в числе свыше 1 000 на клетку), возникают в сферической обла сти уплотненной цитоплазмы — так называемых блефаропластах (греч. blepharon — ресница). Блефаропласты (рис. 2.19) представляют собой четко структурированную форму центроплазмы = центросомы — обычно структурно не означенную область плазмы, которая функционирует как ПОМТ и, например, у лишенных центриолей цветковых растений организует полюса веретена ядерного деления.
1 Имеются в виду прежде всего саговниковые (Сусаdales) и гинкговые (Сinkgoales). — Примеч. ред.
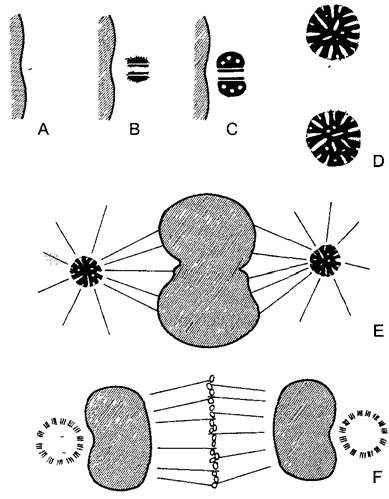
Рис. 2.19. Возникновение заново центриолей базальных тел при микроспорогенезе водного папоротника Marsilea (по P.K.Hepler): А—С — в уплотненной части цитоплазмы вблизи ядерной оболочки возникает симметричная структура из 2 блефаропластов. Они разделяются перед ближайшим ядерным делением (D) и образуют два полюса веретена (Е, F). Из каждого блефаропласта получается в конечном счете примерно 150 базальных телец имеющих жгутики сперматозоидов. Весь процесс в целом показывает, что такие комплексные характерные структуры, как центриоли или же базальные тельца, могут возникать de novo
Базальные тела чаще всего закрепляются сложной системой микротрубочек в цитоплазме, часто будучи еще связанными с клеточным ядром. Микротрубочки в
свою очередь часто ассоциированы с характерными волокнистыми структурами двух видов. Поперечнополосатые волокна системы I построены из нитей толщиной 2 нм. В качестве главного компонента они содержат белок ассемблин массой 34 кДа и не способны к сокращению. Сократимые волокна системы II построены из нитей толщиной 4 — 8 нм и содержат цен- трин.
Генетическая информация всех клеток, как прокариот, так и эукариот, заключена в последовательности нуклеотидов молекул ДНК. У эукариот ДНК содержится прежде всего в ядре, но также в митохондриях и пластидах. Ядро — это основная структура для хранения и размножения (репликации) ДНК, а также для синтеза (транскрипции) и созревания (процессинга) РНК. Все эти процессы происходят в нуклеоплазме (= кариоплазме), которая отграничена от окружающей цитоплазмы двухслойной ядерной оболочкой (англ, nuclear envelope). Она соответствует сферической цистерне ЭР. Отличительная особенность ядерной оболочки — многочисленные поровые комплексы для обмена макромолекулами между ядром и цитоплазмой. Молекулы тРНК, иРНК и образуемые в ядрышке прерибосомы покидают пространство ядра через эти поровые комплексы, а специфичные для ядра белки проникают через них в ядро (см. рис. 2.20; 7.9; 7.17).
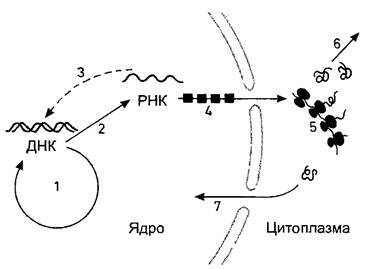
Рис. 2.20. «Центральная догма» молекулярной биологии гласит, что поток информации в клетке идет от ДНК через РНК к белкам: «ДНК создает РНК, РНК создает белок».
Однако ДНК служит не только матрицей для синтеза РНК (транскрипция 2), но еще и осуществляет собственное размножение (репликация 1). От РНК возможен однозначный обратный перевод в последовательности ДНК (обратная транскрипция 3, происходящая, в частности, у РНК-вирусов, которые встраивают свой геном в ДНК клетки хозяина). У эукариот эти процессы и процессинг вновь образованной РНК (4) происходят внутри пронизанной порами ядерной оболочки. Образованные в ядре и процессированные молекулы РНК становятся в цитоплазме активными при синтезе белка на рибосомах (трансляция 5). Многие белки в качестве ферментов управляют обменом веществ и энергии в клетке (6); другие же мигрируют в ядро (7), где они, например, принимают участие в репликации и транскрипции или берут на себя важные функции в хроматине как белки, сопутствующие ДНК
Внутри ядра мембран нет, но имеются молекулы ДНК длиной в сантиметры и дециметры (1 — 100 млрд Да). Необходимый функциональный и структурный порядок обеспечивается своего рода ядерным скелетом — ядерным матриксом. В этом геле из структурных белков распределен дезоксирибонуклеиновобелковый комплекс — хроматин. В нем главенствующую роль играют основные гистоны как белки, непосредственно взаимодействующие с ДНК. Имеются различные степени уплотнения (конденсации) хроматина. Хроматин, участвующий в репликации или транскрипции, находится в ядре в деконденсированной форме (эухроматин). Особенно плотные «хромоцентры», напротив, очень плотно конденсированы и генетически неактивны (гетерохроматин). То же самое относится и к компактным хромосомам во время делений ядер. Активация генов как предпосылка для транскрипции достигается различными негистонными белками, прежде всего факторами транскрипции (ТР), связывающимися с определенными локусами последовательностей ДНК. Различные варианты подавления и активации генов обусловливают дифференцировку клеток тканей многоклеточных организмов (дифференциальная экспрессия генов).
Большинство молекулярных и надмолекулярных структурных компонентов клеточного ядра — функциональные и недолговечные структуры. Ядерная оболочка и ядрышки распадаются, например, на начальных стадиях деления и только в его конечной фазе образуются заново. Ядерный скелет проявляет себя так же как динамическая структура. Его молекулярные компоненты меняются в клеточном цикле — характерной последовательности событий между ядерными делениями и во время них. Единственный компонент ядра, который, один раз возникнув путем репликации, в нормальной ситуации не подвергается никакому преобразованию или разложению, — это ДНК, несущая наследственную информацию (гены).
Между объемом клеточного ядра как обширного архива генетической информации в эукариотической клетке и общим объемом цитоплазмы в нормальном случае имеется соотношение объемов примерно 1:10. Оно сохраняется постоянным при изменении объема клетки. Крупные, богатые цитоплазмой клетки чаще всего имеют также крупные ядра, которые почти всегда отличаются увеличением в несколько раз содержащегося в них набора хромосом (см. 2.2.3.5).
Большая часть ядерной ДНК находится в комплексе с гистонами. Гистоны имеются у подавляющего большинства эукариот. (Исключение составляют только динофлагелляты 1 — см. рис. 11.70, — хроматин которых организован иначе. Однако во всем остальном эти одноклеточные организмы — типичные эукариоты.) Соотношение по массе гистон/ДНК составляет примерно 1:1. Гистоны встречаются в живой клетке только в комплексе с ДНК. Они синтезируются синхронно с ДНК в фазе репликации клеточного цикла (S-фазе) в цитоплазме и сразу же перемещаются в ядро. ДНК, имеющая сильно кислую реакцию, притягивает как полианион молекулы гистонов, которые, в свою очередь, за счет наличия многочисленных остатков лизина и аргинина имеют щелочную реакцию (pI ≈ 12) и представляют собой поликатионы (табл. 2.1) 1 . Расположение в ряд от Н1 до Н4 отражает уменьшение доли лизина и увеличение доли аргинина. Гистоны, особенно НЗ и Н4, лишь незначительно изменились в ходе эволюции. Однако имеются специфичные для определенных тканей вариации, которые могут быть обусловлены дифференциальной активностью несколько отличающихся друг от друга кодирующих гистоны генов (изотипы) или же обратимыми модификациями молекул гистонов после трансляции (ацетилирование или фосфорилирование отдельных аминокислот; см. 7.2.2.2).
1 В последнее время гистоны в очень ограниченном количестве обнаружены и у динофлагеллят. — Примеч. пер.
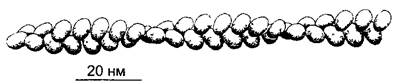
Четыре гистона — от H2A до Н4, которые имеют сходные размеры и конфигурации молекул, самостоятельно, даже без ДНК, образуют плоско-эллипсоидные четвертичные структуры. В этих частицах диаметром 10 нм и толщиной 5 нм имеются 2 молекулы каждого из составляющих гистонов. Поэтому их называют гистонными октамерами, а их компоненты — гистоновыми ядрами (histone cores). Каждый октамер по диаметру, вдоль которого находятся N-концы молекул гистонов с особенно щелочной реакцией, охвачен петлей ДНК, представляющей собой отрезок длиной в 145 пар нуклеотидов (п. н.) (рис. 2.21). Двойная спираль ДНК делает при этом не более 2 оборотов вокруг октамера гистона, а затем идет к следующему. Промежуточный участок длиной примерно 60 п. н., линкер (англ, linker — соединитель), является предпочтительным местом атаки эндонуклеаз. В соответствующих опытах с расщеплением эндонуклеазами сюда присоединялись нуклеогистонные комплексы, имеющие единую массу частиц, — нуклеосомы. В ЭМ очень рыхлый хроматин, свободный от Н1, образует типичную структуру «нитки жемчуга» (рис. 2.22, А).
Таблица 2.1. Обзор основных пяти типов гистонов
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом