
Большинство генов белков промежуточных филаментов у человека кодируют кератины. Как показано в таблицах ниже, известно 28 кератинов, относящихся к типу I, и 26 белков, относящихся ко II типу. В ткани эпителия кератины, которые иногда называются цитокератинами, экспрессируются совместно, т. е. как тип I/тип II. Экспрессия кератинов является настолько характерной особенностью клеток эпителия, что их присутствие может служить признаком эпителиальной ткани. Ткани, которые не экспрессируют кератины (например, клетки эндотелия кровеносных сосудов) не относятся к эпителиальным.
Кератиновые филаменты связаны с межклеточными контактами (десмосомами) и с местами контактов клетки с матриксом (полудесмосомами).
Все вместе образует структурную сеть, которая особенно характерна для многослойного эпителия, например эпидермиса. Основные кератины эпителиального слоя подразделяются на группы в зависисмости от заряда и молекулярной массы. В состав группы К1 (тип II) входят наиболее крупные и основные белки, а в состав группы К19 — более мелкие и кислые белки группы I.
Каждый кератин группы I экспрессируется совместно с кератином группы II, и каждая пара характерна для степени дифференцировки и специализации определенного типа клеток эпителия. Хотя in vitro любой из кератинов, относящихся к I или II типу, будет образовывать филаменты с различными комплементарными кератинами другого типа, in vivo они проявляют гораздо большую селективность, образуя специфические пары.
Экспрессия таких пар кератинов тесно связана со специфическими направлениями дифференцировки эпителия или даже с различными стадиями этого процесса, и присутствие одного представителя пары почти всегда служит признаком наличия другого. С функциональной точки зрения, пары кератинов можно подразделить по крайней мере на три группы: простые, барьерные и структурные кератины.

Экспрессия кератинов характеризуется тканевой специфичностью.
Они экспрессируется попарно: тип I/тип II.
Каждая пара характерна для специфического типа дифференцировки эпителия. Выделены первичные кератины.
Кератины экспрессируются в клетках эпителия. Эпителиальная ткань представляет собой тип тканевой организации, который характеризуется плотной упаковкой клеток в один или несколько слоев. Эпителий образует границу органов, а также секреторных и всасывающих протоков. В этих случаях обычно говорят об однослойном эпителии, клетки которого непосредственно контактируют с подлежащим слоем внеклеточного матрикса (базальной ламины), а свободная поверхность открыта в просвет протока или кишки.
Крайний случай представляет собой многослойный или стратифицированный эпителий, образующий основную барьерную ткань оболочек тела и органов. К этому типу относятся эпидермис, покрывающий внешнюю поверхность тела, а также специализированный эпителий, выстилающий отверстия и полости прилегающих к ним протоков. Клетки стратифицированного эпителия обычно называют кератиноцитами. Ткань стратифицированного эпителия также формирует дополнительные структуры эмбрионального происхождения, состоящие из специализированного сложного эпителия, образующего, например, железы, волосы и ногти, которые могут включать участки, состоящие из простого эпителия.
Наружные клетки полностью сформировавшегося стратифицированного барьерного эпителия плоской или чешуйчатой формы. Этот эпителий обычно состоит из 6-10 слоев клеток Чтобы защитить тело от физических, химических и канцерогенных воздействий, клетки постоянно обновляются.
Клетки эпителия скрепляются вместе посредством десмосом, а также полудесмосом с субстратами. Все контакты в виде плотной сети, состоящей из связок кератиновых филаментов, проходят через цитоплазму каждой клетки. По сравнению с клетками других типов, кератиноциты многослойного эпителия экспрессируют большее количество и более широкий набор белков промежуточных филаментов, что связано с тем, что эти клетки должны противостоять массивным внешним физическим воздействиям.
Состав специфических белков кератиновых филаментов, которые экспрессируются в эпителиальных клетках, зависит от локализации и физиологического состояния последних. Особенно отчетливо это прослеживается в изменении экспрессии кератина, которое происходит при дифференцировке клеток стратифицированного эпителия. К числу наименее дифференцированных клеток стратифицированного эпителия принадлежат базальные клетки, которые еще сохраняют контакт с базальной мембраной. Эти клетки еще способны к делению, некоторые из них являются стволовыми клетками ткани, и делиться крайне редко, однако большая часть клеток делится, обеспечивая тем самым рост ткани.
После того как базальная клетка разделилась, одна из дочерних клеток покидает базальный компартмент и перемещается в первый супрабазальный слой. В результате этого важного перемещения клетка выходит из-под прямого влияния базальной мембраны и сигналов роста и становится коммитированной к терминальной дифференцировке. Затем клетка начинает свой путь к поверхности эпителия. Этот путь представляет собой процесс терминальной дифференцировки, который в конце концов завершается гибелью клетки и ее потерей из ткани. Рисунок ниже иллюстрирует прогрессирующие изменения экспрессии кератинов, связанные с выходом некоторых клеток из пролиферативного компартмента и их коммитированием к дифференцировке.
В процессе развития первыми экспрессируются простые кератины. В этой группе первичные или универсальные кератины представлены белками К8 (тип II) и К18 (тип I). Они присутствуют в ранних эмбриональных клетках и, по-видимому, представляют собой эволюционно наиболее древние кератины. Их экспрессия минимальна, что соответствует наименее дифференцированному состоянию кератинов, характерному для функционального эпителия, пласт которого состоит из поляризованных и плотно, без промежутков, скрепленных между собой клеток.
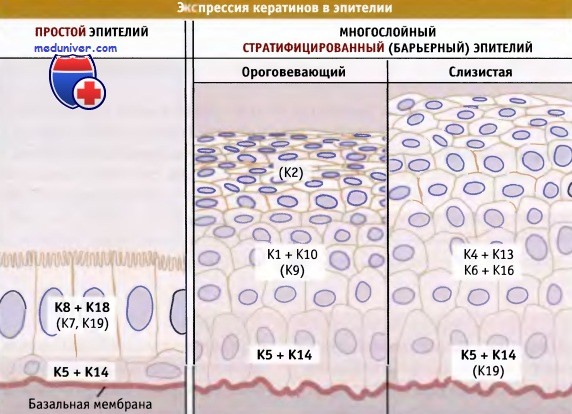
Последовательность экспрессии основных кератинов в однослойном (например, в железистом) и барьерном (например, в ороговевающем или слизистом) эпителии.
Первичные кератины выделены жирным шрифтом. Вариабельные или минорные кератины не показаны.
Экспрессия кератинов зависит от положения клеток в ткани и, таким образом, от ее пролифертивного статуса:
утрата контакта клеток с базальным слоем приводит к их выходу из цикла и к вступлению в дифференцировку.
У позвоночных К8 и К18 представляют собой белки с наиболее консервативной первичной структурой и присутствуют в клетках на всех ступенях развития, от яйцеклетки до зрелой ткани. Все эмбриональные клетки экспрессируют К8 и К18 до стадии гаструляции, когда некоторые эктодермальные клетки начинают дифференцироваться, образуя слой мезодермы, прекращают синтез К8/К18, и начинают экспрессировать виментин, белок, относящийся к типу III.
Белки К8 и К18 продолжают экспрессироваться в клетках эмбрионального эпителия до тех пор, пока эти клетки не станут коммитированными к специфическому направлению морфогенеза, т. е. когда они начинают экспрессировать тканеспецифические белки промежуточных филаментов. Во взрослом организме К8 и К18 характерны для клеток простого эпителия, обладающих секреторными и/или всасывающими функциями, например клеток желез, печени, дыхательного эпителия и желудочно-кишечного тракта. Эти белки экспрессируются в клетках различных карцином, и антитела к ним широко используются для диагностики различных патологических состояний. Наряду с первичными кератинами, известны, по меньшей мере, еще два простых кератина, К7 (близкий к К8 в основном содержится в клетках протоков желез) и К20 (близкий к К18, который содержится в отделах желудочно-кишечного тракта).
Читайте также: Ширина ткани для пододеяльника
Барьерные кератины характерны для стратифицированного эпителия. К первичным, или основным, кератинам этой группы относятся К14 (тип I) и К5 (тип II), присутствующие в кератиноцитах базального слоя стратифицированного сквамозного эпителия, например эпителия кожи. Этот слой, состоящий из одного ряда базальных клеток, экспрессирующих К5/К14, представляет собой наименее дифференцированный тканевой компартмент, сохраняющий способность к пролиферации. В сложном железистом эпителии, наряду с простыми клетками, экспрессирующими К8/К18, находятся базальные клетки, экспрессирующие К5/К14. В некоторых тканях базальные пролиферирующие клетки экспрессируют другие кератины, например К19, К15 и К6/К17.
Когда клетки выходят из базального слоя, они прекращают продуцировать К5/К14 и начинают экспрессировать вторую пару кератинов, специфичную для дифференцирующихся клеток. В эпидермисе это кератины К1 типа II и К10 типа I. Экспресия вторичных кератинов в супрабазальных слоях клеток зависит от типа ткани. Один набор кератинов, представляющий собой изоформы К6, а также К16 и К17, проявляет свойства стрессорных белков эпидермиса, и их синтез индуцируется вскоре после раневых повреждений и воспалительного процесса. В других тканях эти «стрессорные» кератины экспрессируются постоянно, и этим тканям свойственна роль характерных индукторов воспаления.
Таким образом, пролиферативный компартмент (базальный слой клеток) многослойного барьерного эпителия характеризуется спектром синтеза кератинов отличным от дифференцирующегося компартмента (супрабазальных слоев). Для этих барьерных тканей выход клеток из цикла тесно связан с остановкой синтеза первичных кератинов и с началом синтеза вторичных, специфичных для процесса дифференцировки. Кератины клеток супрабазального слоя обеспечивают большую тканевую упругость, и экспрессия нескольких вторичных кератинов может препятствовать делению клеток в митозе и, таким образом, несовместима с пролиферацией.
Третья группа «структурных» кератинов включает большое количество белков промежуточных филаментов, которые экспрессируются только в специализированных поддерживающих структурах, таких как волосы и ногти. К ним относятся кератины клеток волос или трихоцитов, образующих эти структуры, а также специализированные кератины, образующиеся в продуцирующих их эпителиальных клетках. Первичная структура этих двух типов структурных кератинов различна, и считается, что у позвоночных они представляют собой поздний продукт эволюции.
Последовательная экспрессия структурных кератинов особенно характерна для волосяных фолликулов. Первая группа структурных кератинов экспрессируется в концентрических слоях эпителиальных трубочек внутреннего корневого чехла волосяного фолликула. Эта структура становится очень прочной и формирует основную часть волоса. Вторая группа опорных кератинов (трихоцитов или клеток волоса) экспрессируется в основной части волоса, ногтях, некоторых опорных клетках поверхностных сосочков языка (что характерно для языка кошачьих), и в небольшом количестве в тимусе. Эти кератины трихоцитов отличаются высоким содержанием цистеина и пролина в головных и хвостовых доменах.
Последнее позволяет им образовывать дисульфидные связи с белками, которые находятся в цитоплазме клеток волоса по мере дифференцировки и упрочения последних. Эти белки не входят в состав филаментарных структур и называются белками, связывающимися с кератином. Наличие дисульфидных связей приводит к образованию очень прочных структур, формирующих поддерживающую ткань.
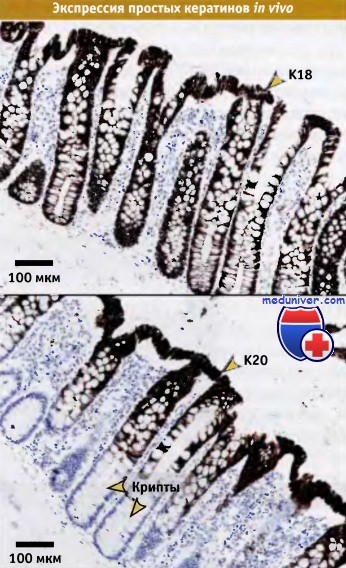 Иммуногистохимическое окрашивание простых кератинов на срезе слизистой толстого кишечника.
Иммуногистохимическое окрашивание простых кератинов на срезе слизистой толстого кишечника.
Локализация двух кератинов I типа, К18 и К20 определена с помощью двух моноклональных антител с последующим иммунопероксидазным выявлением.
Кератины окрашиваются коричневым цветом на фоне голубой окраски гематоксилином.
Первичный кератин К18 находится во всех клетках простого эпителия, в то время как К20 специфичен для определенных клеток желудочно-кишечного тракта.
Как видно на препарате, его присутствие отмечено только в более дифференцированных клетках эпителия, по мере миграции клеток вверх от основания крипты. 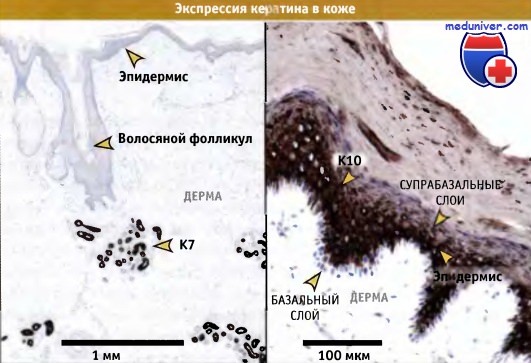 Тканевая специфичность экспрессии кератина в коже.
Тканевая специфичность экспрессии кератина в коже.
На срезах, окрашенных антителами к К7, простому кератину типа II, видны только секреторные клетки потовых желез (иммунопероксидазой окрашиваются в темно-коричневыц цвет).
Многослойный ороговевающий барьерный эпителий (эпидермис) не окрашивается антителами, и после окраски гематоксилином ядра имеют бледно-голубую окраску.
Справа: срез плотного эпидермиса, окрашенный антителами на К10,
тип I вторичный или тканеспецифичный кератин, экспрессирующийся в постмитотических супрабазальных клетках ороговевших барьерных тканей.
Базальный слой клеток не окрашивается антителами, и ядра окрашены в синий цвет. 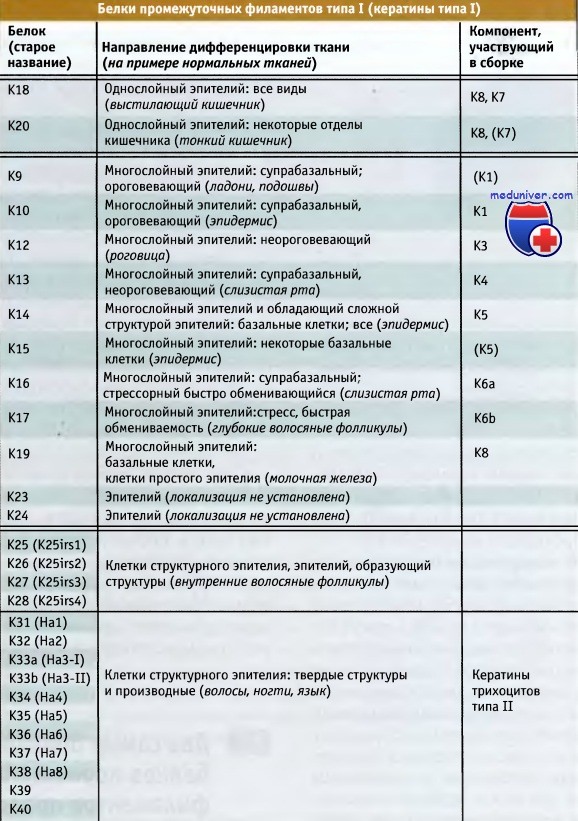 Белки промежуточных филаментов человека, относящиеся к гомологичной группе типа I (кератины типа I).
Белки промежуточных филаментов человека, относящиеся к гомологичной группе типа I (кератины типа I).
Белки сгруппированы в простые кератины (клетки простого эпителия), барьерные кератины (характерные для многослойного чешуйчатого эпителия и эпителия сложной структуры)
и в два типа структурных кератинов, связанных с эпидермальными элементами  Белки промежуточных филаментов человека, относящиеся к гомологичной группе типа II (кератины типа II).
Белки промежуточных филаментов человека, относящиеся к гомологичной группе типа II (кератины типа II).
Белки сгруппированы в простые кератины (клетки простого эпителия),
барьерные кератины (характерные для многослойного ороговевающего эпителия и эпителия сложной структуры)
и в два типа структурных кератинов, связанных с эпидермальным элементами.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
БАРЬЕРНЫЕ ФУНКЦИИ
Барьерные функции — функции, осуществляемые особыми физиологическими механизмами (барьерами) для защиты организма или отдельных его частей от изменений окружающей среды и сохранения необходимого для нормальной жизнедеятельности органов и тканей относительного постоянства состава, физико-химических и биологических, свойств внутренней среды (крови, лимфы, тканевой жидкости). Как и все другие приспособительные свойства организма, барьерные функции выработались в процессе эволюции. По мере усложнения, дифференцировки и совершенствования многоклеточных организмов совершенствовались барьерные функции, регулирующие обмен веществ между организмом и средой, а также способствующие предохранению клеток органов и тканей от соприкосновения с повреждающими их агентами, чужеродными веществами, ядами, токсинами, продуктами нарушенного обмена, вирусами и т. д.
Условно различают барьеры внешние и внутренние. К внешним барьерам относят: 1) кожу, охраняющую животный организм от физических и химических изменений в окружающей среде и принимающую участие в терморегуляции. Кожный барьер препятствует проникновению бактерий, токсинов, ядов в организм и способствует выведению из него некоторых продуктов метаболизма; 2) дыхательный аппарат, который, помимо своей основной функции газообмена, задерживает различные вредные вещества, находящиеся в атмосфере; 3) пищеварительный аппарат, через который поступают необходимые питательные вещества. В нем они претерпевают соответствующие изменения, теряют антигенные свойства, становясь пригодными для усвоения и использования живыми системами; 4) печень, обезвреживающая ряд чуждых организму ядовитых соединений, поступивших с пищей или образовавшихся в полости кишечника; 5) почки, регулирующие постоянство состава крови и освобождающие ее от конечных продуктов метаболизма. К внешним барьерам многие авторы относят также ретикулоэндотелиальную систему, участвующую в обезвреживании чуждых и болезнетворных агентов.
Внутренние барьеры регулируют поступление из крови в органы и ткани необходимых энергетических ресурсов и своевременный отток продуктов клеточного обмена веществ (очищение, клиренс), что обеспечивает постоянство состава, физико-химических и биологических свойств тканевой (внеклеточной) жидкости и сохранение их на определенном оптимальном уровне. Одновременно они препятствуют поступлению из крови в органы и ткани чужеродных и ядовитых веществ.
Основоположником учения о барьерных функциях является Л. С. Штерн, которая впервые на Международном физиологическом конгрессе в Бостоне (1929) высказала предположение, что между кровью и тканевой жидкостью находятся дифференцированные защитно-регуляторные приспособления, названные ею гисто-гематическими барьерами. Каждый орган, по представлению Л. С. Штерн, имеет свою адекватную среду (непосредственная питательная среда или микросреда), так как кровь не приходит в соприкосновение с клетками органов. Функциональная характеристика отдельных барьеров определяется физиологическими и морфологическими особенностями соответствующих органов и тканей. Особенностью каждого гисто-гематического барьера является его избирательная (селективная) проницаемость, то есть способность пропускать одни вещества и задерживать другие.
Читайте также: Что такое крепитация мягких тканей
В литературе внутренние барьеры получили различные названия: тканевых, гемато-паренхиматозных (А. А. Богомолец и Н. Д. Стражеско), гистиоцитарных, сосудисто-тканевых (А. В. Лебединский), биологических, физиологических и т. д. Однако наиболее распространен термин «гисто-гематические барьеры», хотя он не отражает их ведущей роли в осуществлении обмена между общей внутренней средой (кровью) и микросредой органов и тканей. Учение о барьерных функциях не сводится к проблеме биологических мембран. Оно значительно шире, хотя одним из механизмов, определяющих функциональное состояние барьеров, является проницаемость мембран (см. Проницаемость).
К гисто-гематическим барьерам могут быть отнесены все без исключения барьерные образования между кровью и органами. Некоторые авторы признают существование специализированных барьеров, имеющих особо важное значение для жизнедеятельности организма. К ним обычно относят более подробно изученные гемато-энцефалический барьер (между кровью и центральной нервной системой), гемато-офтальмический барьер (между кровью и водянистой влагой глаза), гемато-лабиринтный барьер (между кровью и эндолимфой лабиринта), барьер между кровью и половыми железами. К гисто-гематическим барьерам относят также барьеры между кровью и жидкими средами организма (цереброспинальной жидкостью, лимфой, плевральной, синовиальной жидкостями). Они получили название гемато-ликворного, гемато-лимфатического, гемато-плеврального, гемато-синовиального барьеров. Плацентарный барьер (между матерью и плодом), хотя и не относится к гисто-гематическим барьерам, осуществляет чрезвычайно важную функцию защиты развивающегося плода (см. Плацента).
Структура гисто-гематических барьеров определяется в значительной степени строением органа, в систему которого они входят. Она отличается некоторыми специфическими особенностями в различных органах и тканях и варьирует в зависимости от их морфологических и физиологических особенностей. Основным структурным элементом гисто-гематических барьеров являются кровеносные капилляры. Установлено, что эндотелий капилляров в разных органах обладает характерными морфологическими особенностями. По форме ядра, строению его оболочки, структуре и количеству хроматина эндотелиальные клетки различных органов значительно отличаются друг от друга. Складывающиеся в процессе онтогенеза чрезвычайно изменчивые особенности эндотелиальных клеток являются морфологической основой избирательной проницаемости гисто-гематических барьеров. Различия в механизмах осуществления барьерных функций находят свое отражение в структурных особенностях основного вещества (неклеточных образований, заполняющих пространства между клетками), способного импрегнироваться серебром. Основное вещество образует мембраны, окутывающие макромолекулы фибриллярного белка, оформленного в виде протофибрилл, составляющего опорный остов волокнистых структур.
Непосредственно под эндотелием располагается базальная мембрана капилляров, в состав которой входит большое количество нейтральных мукополисахаридов. Базальная мембрана, основное аморфное вещество и волокна составляют барьерный механизм, в котором главным реактивным и лабильным звеном, по мнению некоторых исследователей, является основное вещество. А. А. Богомолец придавал большое значение барьерной функции соединительной ткани, обладающей также свойствами депо, из которого организм черпает питательные вещества, необходимые для деятельности клеточных элементов.
Согласно современным представлениям, в систему гисто-гематических барьеров включаются также барьеры внутриклеточные. Электронная микроскопия позволила проникнуть в субмикроскопическую организацию клетки и тем самым подойти к изучению этих барьеров. Барьерные механизмы клетки состоят из однотипных трехслойных липопротеиновых мембран, являющихся основными структурными элементами митохондрий, системы каналов, эндоплазматической сети, аппарата Гольджи и клеточной оболочки. Наличие цитоплазматической мембраны позволяет в известной степени понять избирательность проницаемости гисто-гематических барьеров (перенос электронов, трансформацию энергии, энзиматическое расщепление, транспорт ионов и метаболитов, кинетику некоторых биосинтетических процессов).
Исследования показали, что химический состав, физико-химического и биологического свойства непосредственной питательной среды органов (тканевой жидкости) обусловлены: 1) поступлением веществ из крови, которое зависит от сопротивляемости гисто-гематического барьера данного органа в направлении кровь —>ткани; 2) поглощением и использованием составных частей тканевой жидкости клетками и неклеточными элементами в процессе межуточного обмена; 3) поступлением продуктов клеточного и тканевого обмена (метаболитов) в тканевую жидкость; 4) удалением метаболитов из тканевой жидкости, то есть переходом их из непосредственной питательной среды органа в кровь через гисто-гематический барьер данного органа (ткани —> кровь).
Гисто-гематический барьер органа определяет функциональное состояние последнего, его деятельность, способность противостоять вредным влияниям. Значение барьера заключается в задержке перехода того или иного чужеродного вещества из крови и ткани (защитная функция) и в регуляции состава и свойств непосредственной питательной среды органа, то есть создании оптимальных условий для жизнедеятельности его клеточных и неклеточных элементов (регуляторная функция), что особо важно для всего организма и его отдельных частей. Осуществляя регуляторную функцию, гисто-гематические барьеры способствуют сохранению органного и клеточного гомеостаза.
Функциональное состояние каждого гисто-гематического барьера характеризуется математической величиной, отражающей соотношение концентрации того или иного вещества в органе и в крови. Эта величина получила название коэффициента проницаемости. Однако фактически она соответствует распределению исследуемого вещества между тканями и кровью, поскольку содержание его в ткани зависит не только от поступления из крови в ткани или из ткани в кровь, но и от интенсивности метаболизма клеток. Функциональное состояние гисто-гематических барьеров не может быть охарактеризовано одной лишь его проницаемостью и, следовательно, коэффициент проницаемости правильнее рассматривать как коэффициент распределения. Функциональное состояние гисто-гематических барьеров обусловлено не только проницаемостью или сопротивляемостью (резистентностью) их к чужеродным или свойственным организму хим. соединениям, но в основном физиологической активностью, то есть способностью создавать и поддерживать наиболее благоприятные условия для нормальной жизнедеятельности органов, тканей и организма в целом.
В зависимости от активности гисто-гематических барьеров их сопротивляемость (или проницаемость) по отношению к тем или другим веществам может повышаться или снижаться, что ведет к увеличению или уменьшению величин коэффициентов распределения. Так, например, при значительном повышении концентрации того или другого вещества в крови содержание его в органе может не измениться или незначительно повыситься. При этом величина коэффициента распределения уменьшается, что является показателем высокой активности соответствующего гисто-гематического барьера и в то же время снижения его проницаемости. В других случаях содержание вещества в органе нарастает при постоянной или низкой его концентрации в крови. Повышенный в этом случае коэффициент распределения указывает на уменьшение активности барьера и одновременно на его высокую проницаемость.
Функционированием гисто-гематических барьеров объясняют все явления, предотвращающие, понижающие, замедляющие и даже облегчающие поступление веществ в органы и ткани и удаление из них продуктов межуточного обмена. Многочисленные физические, химические и морфологические концепции, предложенные для объяснения избирательной проницаемости гисто-гематических барьеров, не решают проблемы барьерных функций. В основе барьерных функций лежат механизмы диализа, ультрафильтрации, осмоса, а также изменение электрических свойств, растворимости в липидах, тканевого сродства или метаболической активности клеточных элементов. Барьеры активно отбирают из крови необходимые для жизнедеятельности органов и тканей вещества и выводят из их микросреды продукты обмена.
Одним из механизмов барьерных функций является активный транспорт некоторых электролитов через мембраны. Установлено, что переход биологически активных веществ (метаболитов, медиаторов, ферментов, гормонов) через гисто-гематические барьеры зависит не только от величины молекул, размеров пор в мембранах, электрического заряда, растворимости в липидах, но в основном от потребностей органа, нервных и гуморальных влияний, гемодинамики (скорости кровотока), микроциркуляции, площади открытых и резервных капилляров, наличия или отсутствия функциональных и морфологических нарушений. Важное значение для состояния барьеров имеет наличие в них метаболических структур, то есть тканевых элементов, способных нейтрализовать, разрушать или связывать содержащиеся в крови вещества. Таким образом, гисто-гематические барьеры можно рассматривать как саморегулирующуюся систему, представляющую одно из звеньев комплексного нейро-гуморально-гормонального регуляторного аппарата, обеспечивающего состояние гомеостаза (см.).
Читайте также: Загребаева л в роспись по ткани лоскутное шитье
Гисто-гематические барьеры контролируют своевременное поступление в непосредственную питательную среду органов и тканей адекватной гуморальной информации о состоянии регуляторных метаболических систем в различных частях организма. Проникая через гисто-гематический барьер в орган, биологически активные вещества оказывают свое действие на эффекторные клетки и на специфические хеморецепторы, что ведет к возникновению как местных, так и распространенных (общих) физиологических и биохимических реакций. Примером может служить действие веществ, проникающих из крови в различные по своему строению, химическому составу и функциям образования центральной нервной системы через гемато-энцефалический барьер (см.). Доказано существование активной биологической мембраны между кровью и жидкостями глаза, регулирующей состав внутриглазных жидкостей. Этой биологической мембране было дано название гемато-офтальмического барьера (см.).
В некоторых случаях механизмы регуляции функций оказываются недостаточными, и накопившиеся в крови биологически активные вещества проникают, напр., в различные нервные структуры, обычно защищенные гемато-энцефалическим барьером, вызывая эффекты, отличающиеся от обычных. При этом происходит усиление компенсаторно действующей системы (например, симпатической при накоплении в крови парасимпатомиметических веществ и наоборот), что имеет важнейшее значение для восстановления нарушенного гомеостаза.
Физиологические и биохимические процессы, протекающие как в здоровом, так и больном организме, состояние органа, его трофика, регуляция функций, взаимоотношение между отдельными органами и физиологическими системами тесно связаны с состоянием гисто-гематических барьеров. Нарушение сопротивляемости барьеров по отношению к различным чужеродным веществам и продуктам нарушенного метаболизма, циркулирующим в крови, может явиться во многих случаях причиной возникновения патологического процесса в отдельных органах и в целостном организме. Нечувствительность или иммунитет, так же как и сродство или способность органа захватывать определенные химические вещества, бактерии, токсины, зависит в той или иной мере от состояния соответствующего гисто-гематического барьера, поскольку обязательной предпосылкой непосредственного воздействия на клеточные элементы является проникновение действующего начала в микросреду органа.
Снижение сопротивляемости соответствующего гисто-гематического барьера делает орган более восприимчивым, а повышение ее — менее чувствительным к химическим соединениям, образовавшимся в процессе обмена или введенным в организм с экспериментальной или лечебной целью.
Оценка состояния отдельных гисто-гематических барьеров в эксперименте или клинике требует всестороннего исследования тканевой жидкости, что на современном уровне знаний практически неосуществимо. Поэтому предложено большое число разнообразных методов, позволяющих в известной степени как в лабораторном опыте, так и при обследовании больных в клинической практике оценить состояние того или иного гисто-гематического барьера. Наиболее распространенными в эксперименте остаются предложенные для изучения тканевой проницаемости классические методы введения в кровь красителей (коллоидных, полуколлоидных, кристаллических), туши, некоторых сложных химических соединений и радиоизотопных индикаторов с последующим определением их концентрации и распределения в органах и тканях. С этой целью используются методы световой, прижизненной (витальной), люминесцентной и электронной микроскопии, микросжигания, определения радиоактивности и т. д. Как в эксперименте, так и в клинике применяются методы сравнительного исследования состава притекающей к органу (артериальной) и оттекающей от него (венозной) крови. Для суждения о защитной и регуляторной функциях барьеров между кровью и жидкими средами организма (лимфой, цереброспинальной, плевральной, синовиальной жидкостями) производится количественное определение свойственных организму или введенных извне веществ в крови и соответствующих жидкостях.
Для оценки состояния гисто-гематических барьеров в направлении ткани кроль испытуемое вещество обычно вводится в ткани (внутрикожно, подкожно, внутримышечно) и определяется скорость его всасывания или при введении радиоизотопных индикаторов — время полуудаления.
Для оценки барьерных функций целостного организма испытуемое вещество вводится внутривенно и в течение определенного времени исследуется его выделение из крови или при введении радиоизотопных индикаторов — время полуудаления.
Большая пластичность гисто-гематических барьеров, их лабильность и приспособляемость к постоянно меняющимся условиям внешней и внутренней среды играют важную роль в жизнедеятельности организма. Барьерные функции меняются в зависимости от возраста, пола, нервных, гуморальных и гормональных взаимоотношений в организме, тонуса и реактивности вегетативной нервной системы, многочисленных внешних и внутренних воздействий. Исследования ряда авторов показали, что функциональное состояние гисто-гематических барьеров различных органов может избирательно изменяться при действии на организм разнообразных факторов (смена сна и бодрствования, голодание, утомление, травматические поражения, действие ионизирующей радиации и т. д.).
Нек-рые содержащиеся в крови и тканях или введенные извне биологически активные вещества (например, ацетилхолин, гистамин, кинины, особенно брадикинин, некоторые ферменты, в первую очередь гиалуронидаза) в физиологических концентрациях снижают сопротивляемость гисто-гематических барьеров и тем самым повышают переход веществ из крови в органы и ткани. Противоположное действие оказывают катехоламины, соли кальция, витамин Р. При патологических состояниях организма барьерные функции нередко перестраиваются, сопротивляемость гисто-гематических барьеров повышается или снижается. В одних случаях эта перестройка усиливает, в других ослабляет течение заболевания. Снижение сопротивляемости гисто-гематических барьеров делает органы более восприимчивыми к ядам и инфекциям, по некоторым данным, усиливает опухолевый рост. Напротив, повышение сопротивляемости может носить в определенных случаях защитный или компенсаторный характер. Учитывая, что в большинстве случаев гисто-гематические барьеры препятствуют поступлению в органы введенных с лечебной целью лекарственных веществ и антител важное значение для клиники имеет проблема регулирования функционального состояния барьеров. Установлено, что облучение (общее или местное) разными участками светового спектра (инфракрасным и ультрафиолетовым), воздействие ультракороткими, высокочастотными волнами, рентгеновскими лучами, ультразвуком, электромагнитным полем сверхвысокой частоты, а также введение в организм некоторых гормонов (например, кортизона), психотропных веществ, витаминов и т. д. снижает сопротивляемость гисто-гематических барьеров. Все эти методы могут быть использованы в клинической практике для целенаправленного изменения состояния барьерных функций. Искусственное снижение сопротивляемости того или иного гистогематического барьера путем различных физических или фармакологических воздействий может повысить или расширить действие лекарственных препаратов, не проникающих в микросреду органа, в то время как повышение сопротивляемости служит целям профилактики при инфекциях, интоксикациях, опухолевом росте и т. д. В определенных случаях для непосредственного воздействия на пораженный орган химического соединения, лекарственные препараты, лечебные сыворотки вводятся в обход барьера (например, в цереброспинальную жидкость, плевральную и синовиальные полости и т. д.) или в питающую орган артерию.
Библиогр.: Гисто-гематические барьеры, под ред. Л. С. Штерн, М., 1961; Кассиль Г. Н. Гемато-энцефалический барьер, М., 1963; Проблемы гисто-гематических барьеров, под ред. Л. С. Штерн, М., 1965; Развитие и регуляция гисто-гематических барьеров, под ред. Л. С. Штерн, М., 1967; Структура и функция гисто-гематических барьеров, под ред. Я. А. Росина, М., 1971; Физиология и патология гисто-гематических барьеров, под ред. Л. С. Штерн, М., 1968; Штерн Л. С. Непосредственная питательная среда органов и тканей, М., 1960; Gellhоrn E. et Regnier J. La permeabilite en physiologie et en pathologie generale, P., 1936.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом