
Белки костной ткани особенности костного коллагена i типа
• Основная функция коллагена состоит в обеспечении структурной опоры тканей
• Коллагены представляют собой семейство, состоящее более чем из 20 различных белков внеклеточного матрикса. Эти белки — наиболее распространенные в царстве животных
• Все коллагены организованы в тройные спирализованные «коллагеновые субъединицы», обладающие суперспиральной структурой и состоящие из трех отдельных полипептидов
• Коллагеновые субъединицы выходят из клеток и затем, во внеклеточном пространстве, собираются в более крупные фибриллы и волокна
• Мутации в коллагеновых генах вызывают множество патологических состояний, начиная от появления морщин до развития хрупкости костей и таких тяжелых заболеваний, как образование кожных волдырей
Семейство коллагенов включает более 20 белков, которые относятся к наиболее распространенным белкам клеток животных. У многоклеточных организмов коллагены существуют по крайней мере 500 млн лет. Почти все клетки животных синтезируют и секретируют по меньшей мере одну из форм коллагена.
Коллагены обеспечивают тканям структурную поддержку и существуют во множественных формах, организованных в различные структуры. Все белки семейства коллагенов характеризуются одним общим свойством: они собраны в тонкие (примерно 1,5 нм диаметром) тройные спиральные суперспирализованные структуры, состоящие из трех субъединиц коллагеновых белков, которые удерживаются вместе ковалентными и нековалентными связями.
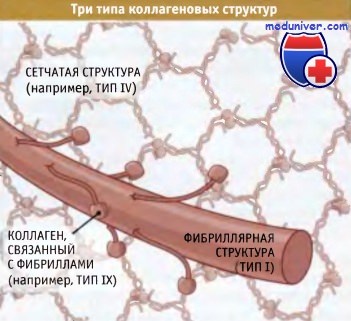
Коллагеновые субъединицы собираются в тройные спиральные структуры, которые организуются в фибриллы или в сеть,
где они связаны между собой другими белками внеклеточного матрикса, включая коллагены, связанные с фибриллами.
Суперспирализованные структуры бывают трех типов — фибриллярные, слоистые и связанные с фибринами:
• В фибриллярных коллагенах суперспирализованные спирали организованы в фибриллы или «канаты», которые обеспечивают прочность структуры вдоль единственнной оси (такая структура напоминает прочный стальной трос, образованный проволочными пучками). Когда эти фибриллы собраны в параллельно расположенные пучки, как в сухожилиях, они обеспечивают невероятную прочность структуры, которая способна противостоять усилиям, развиваемым мышцами, закрепленными на костях.
• Слоистые коллагены представляют собой сеть, состоящую из суперспирализованных спиральных структур. Они в меньшей степени устойчивы к мышечным усилиям, но гораздо лучше противостоят растяжению в нескольких направлениях. Сеть таких структур, например, характерна для кожи.
• Третий тип коллагена, известный как «фибриллярные связки», образует суперспирализованные спиральные структуры, связывающие коллагеновые фибриллы вместе.
Независимо от организации, коллагены образуют основную каркасную структуру внеклеточного матрикса. Такие входящие во внеклеточный матрикс белки, как фибронектин и витронектин, связываются с коллагенами и вплетены в структуры, образованные коллагеновым каркасом. Один из представителей семейства коллагенов представляет собой трансмембранный белок, который участвует в формировании межклеточных контактов.
Существует примерно 20 различных типов коллагена, большая часть которых может быть сгруппирована в четыре класса. Каждая из тройных спиралеобразных структур обозначается римской цифрой (I, II, III и т. д.). Каждая коллагеновая субъединица имеет обозначение как субъединица а, а ее типу присвоен номер (а1, а2, а3 и т. д.), после которого римской цифрой обозначен тип, в котором она находится. Например, основной фибриллярный коллаген хвоста (и других тканей) крыс относится к типу I и состоит из двух копий субъединиц а1(1) и одной копии субъединицы а2 (I).
На рисунке ниже представлена структура коллагеновых волокон. Три полипептидные субъединицы параллельно обернуты вокруг друг друга и образуют суперспирализованную спиральную структуру длиной 300 нм. Для коллагенов характерна повторяющаяся последовательность аминокислот, содержащая элемент глицин-X-Y, где X и Y могут представлять собой любую аминокислоту, но обычно это пролин и гидроксипролин соответственно.
Такая последовательность способствует плотной упаковке трех субъединиц и облегчает образование суперспиральной структуры. Субъединицы длиной 300 нм скрепляются вместе посредством ковалентных связей, которые образуются между N-концевым участком одной субъединицы и С-концевым участком примыкающей к ней. Суперспирализованные спиральные структуры располагаются параллельно, образуя между собой небольшие зазоры (64-67 нм). Эти зазоры обеспечивают характерный вид (исчерченность) фибрилл, видимый в электронном микроскопе.

Коллагеновые белки подразделяются на четыре основные группы, которые отличаются по молекулярной формуле,
характеру полимерных форм и по распределению в тканях. Некоторые группы включают коллагены нескольких типов.
Полностью собранные коллагеновые структуры (фибриллярные или сетчатые) по размерам оказываются гораздо больше, чем сами клетки; некоторые фибриллы могут достигать нескольких миллиметров длины. Таким образом, субъединицы коллагена синтезируются и секретируются в виде суперспирализованных спиральных структур, и окончательные этапы их сборки происходят вне клетки. Как показано на рисунке ниже, синтез коллагена и дальнейший его процессинг происходят на протяжении всего секреторного пути. При синтезе коллагеновые белки направляются в гранулярный эндоплазматический ретикулум (ЭПР) при участии частиц, распознающих сигнал и связанного с ними белкового аппарата.
Коллагеновые субъединицы синтезируются в виде крайне длинных полипептидов, которые называются проколлагены и содержат пропептиды, представляющие собой «хвосты», расположенные на амино- и карбоксильном концах.
После того как проколлагены попали в просвет ЭР, по мере транспорта из ЭР через аппарат Гольджи и в секреторные везикулы, они претерпевают серию модификаций. В процессе транспорта проколлагена через ЭР и аппарат Гольджи, к боковым цепям пролина и лизина, находящимся в средней части молекул проколлагена, добавляются гидроксильные группы (-ОН). При этом образуются гидроксипролин и гидроксилизин.
Эти модификации обеспечивают правильное образование водородных связей, которые скрепляют вместе три субъединицы в суперспирализованной спиральной структуре. Между амино- и карбоксиконцевыми частями пропептидов образуются дисульфидные связи, которые затем обеспечивают правильное расположение трех проколлагеновых субъединиц с образованием тройной спиральной суперспирализованной структуры. Затем спираль образуется спонтанно, в направлении от С-конца к N-концу.
Читайте также: Краска для ткани темно бордовая
Пропептиды препятствуют взаимодействию суперспирализованных спиралей друг с другом, тем самым предотвращая полимеризацию коллагена в клетке. Когда произошла секреция тройных спиралей проколлагена, ферменты, которые называются протеазы проколлагена, отщепляют пропептиды. Остающийся белок, известный под названием тропоколлагена, почти весь организован в тройную спираль и представляет собой основную структурную единицу коллагеновой фибриллы.
Фибриллы собираются просто: боковые цепи лизина в тропоколлагене модифицируются при действии фермента лизилоксидазы, образуя аллизины. Эти модифицированные лизины образуют ковалентные сшивки, которые обеспечивают полимеризацию тропоколлагенов. Лизилоксидаза представляет собой внеклеточный фермент, и эта стадия сборки фибрилл происходит только после выхода проколлагена из клетки. После сборки фибриллы могут объединяться, образуя большие пучки или волокна, характерные для фибриллярного коллагена.
Принимая во внимание всю важность коллагена в обеспечении структурной поддержки тканей, можно представить, какие тяжелые последствия для организма будет иметь нарушение процесса сборки фибрилл. Мутации в генах, кодирующих синтез коллагенов или ферментов, модифицирующих проколлаген, вызывают развитие множества генетических заболеваний, затрагивающих практически все ткани. Например, коллаген типа I представляет собой основной структурный белок костной ткани. Мутации в коллагеновых генах этого типа служат причиной незавершения остеогенеза, т. н. развития «болезни хрупких костей».
Мутации в коллагеновом гене типа IV приводят к нарушению сборки базальной ламины в большинстве эпителиальных тканей и к развитию такого кожного заболевания, как буллезный эпидермоз.
Клетки связываются с коллагеном посредством специфических рецепторов, называемых интегринами. Эти рецепторы обеспечивают возможность обратимого связывания клеток с коллагенами по мере их движения по внеклеточному матриксу. Интегриновые рецепторы также активируют пути передачи сигналов, так что связывание с коллагенами (и с другими белками внеклеточного матрикса) изменяет активность в клетке биохимических процессов и, таким образом, способствует контролю над ростом и дифференцировкой клеток.
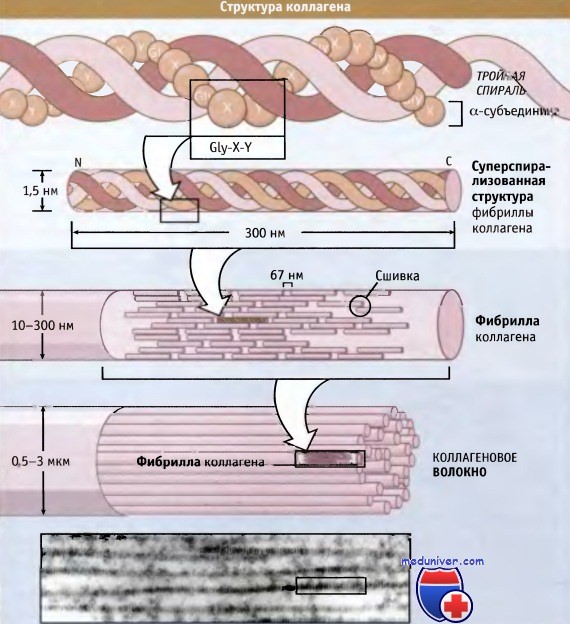 Схематическое строение тройной спиральной структуры коллагена (наверху),
Схематическое строение тройной спиральной структуры коллагена (наверху),
организация витой структуры в фибрилле (в середине) и фибрилл в коллагеновом волокне (внизу).
Наличие 67-нм зазора между соседними спиральными структурами обусловливает видимую в электронном микроскопе исчерченность фибрилл, составляющих волокно. 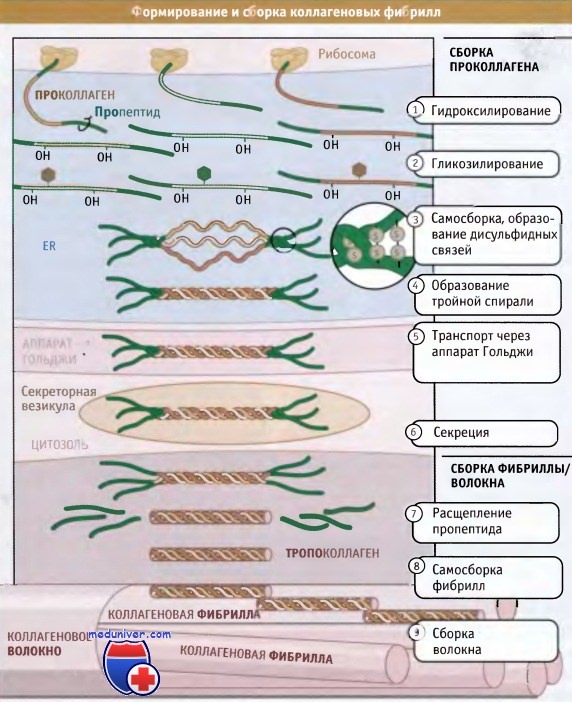 Посттрансляционная модификация и сборка субъединиц проколлагена в тройные спиральные структуры происходит при внутриклеточном перемещении по секреторному пути.
Посттрансляционная модификация и сборка субъединиц проколлагена в тройные спиральные структуры происходит при внутриклеточном перемещении по секреторному пути.
Однако сборка фибрилл происходит во внеклеточном пространстве после секреции тройных спиралей, обладающих витой структурой.
Для упрощения в тройных структурах не показаны гидроксильные группы и сахарные остатки. 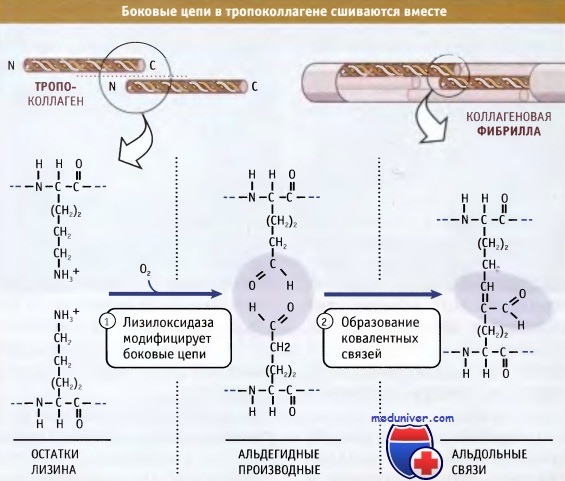 Лизилоксидаза катализирует ковалентное связывание двух боковых цепей лизина с образованием аллизинов (альдегидных производных лизина),
Лизилоксидаза катализирует ковалентное связывание двух боковых цепей лизина с образованием аллизинов (альдегидных производных лизина),
которые затем образуют между собой альдольную связь.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
8 Биохимия костной ткани.
Костная ткань — это особый вид соединительной ткани, включающий компоненты органической и неорганической природы, выполняющий функцию депо Са (99%). Костная ткань имеет особенности строения, которые не встречаются в других видах соединительной ткани. Основные особенности кости — твердость, упругость, механическая прочность.
Состоит из клеток и костного матрикса (межклеточного вещества). Костный матрикс составляет 50% сухого «веса и состоит из неорганической (50%) и органической (25%) частей и Н2О (25%).
Неорганическая часть в значительном количестве содержит Са (25%) и Р (50%), образующие кристаллы гидроксиапатита, а также другие компоненты: бикарбонаты, цитраты, соли Mg 2+ , K + , Na + и др.
Органическая часть образована коллагеном, неколлагеновыми белками, гликозаминогликанами (хондроитинсульфат, кератан-сульфат).
Собственно костные неколлагеновые белки представлены сиалопротеинами, протеогликанами, фосфопротеинами и сложным белком, содержащим углеводный компонент и ортофосфат. От правильного набора матриксных белков, особенностей строения, а также специфического аминокислотного состава зависит отложение гидроксиапатита, создавая необходимую концентрацию Са для процесса минерализации.
Сиалопротеины имеют молекулярную массу 70000. 50% — это углеводы, 12% -сиаловая кислота. Большинство углеводов — это олигосахариды (фруктоза, галактоза, глюкоза, манноза, пентоза, галактозамин). До 30% серина и другие аминокислоты: аспарагиновая и глутаминовая, ковалентно связанные с фосфатом. Присутствие этого белка обеспечивает:
Неколлагеновых белков в костной ткани около 200, они составляют 3-5% от ее массы или 15-17% от массы ее деминерализованного и высушенного внеклеточного органического матрикса. Все они участвуют в процессах обеспечения гистогенеза, самоподдержания, иммунологические свойства на протяжении всей жизни и репарации костной ткани.
Кальций-связывающие белки костной ткани.
Остеонектин — молекулярная масса 32 кДа. Он имеет кальций-связывающие участки, образованные сиаловыми кислотами и ортофосфатом, придающие возможность взаимодействия с коллагеном и избирательно с гидроксиапатитом. Он поддерживает в присутствии коллагена осаждение Са и РО4 3- .
Остеопонтин — молярная масса 41,5 кДа, богат дикарбоновыми аминокислотами и фосфосерином, 30 остатков моносахаридов, 10 остатков сиаловых кислот. Он способен фиксировать остеобласты в участках физиологического и репаративного костеобразования. Его синтез резко возрастает во время трансформации вирусов.
Остеокалъцин — это гла-содержащий протеин.
Дело в том, что костная, как и другие ткани, содержит белки, которые подвергаются посттрансляционной модификации с помощью витамин К-зависимых ферментов, в результате чего образуются остатки у-карбоксилглутаминовой кислоты (gla). Модифицированная таким образом аминокислота придает белкам способность связывать Са 2+ с помощью расположенных по соседству карбоксильных групп. Молекула этого белка состоит из 49 аминокислотных остатков (в 17-ом, 21-ом, 24-ом положениях — остатки у-карбоксилглутаминовой кислоты). Роль их — связывать кристаллы гидроксиапатита и тем самым способствовать их накоплению в ткани.
Синтез остеокальцина зависит не только от витамина К, но и D, что подчеркивает его связь с процессом минерализации.
Читайте также: Мембранная ткань для зимней одежды характеристики
gla-протеин-матрикса (молекулярная масса — 15000 ). Он сохраняется в матриксе кости после деминерализации, в отличие от остеокальцина, который легко экстрагируется в этот период. Остатков у-карбоксилглутаминовой кислоты до шести. Он связывает минеральные кристаллы и легко растворимый в воде костный морфогенетический белок, доставляя его к клеткам-мишеням.
Протеин-S — синтезируется в печени, участие в метаболизме костной ткани доказывается фактом изменения скелета у пациентов с дефицитом этого белка. Но, еще невыясно, каким типом клеток костной ткани он синтезируется.
Протеогликаны — класс сложных соединений, состоящих из различных белков, содержащих олигосахариды, связанные с гликозаминогликанами (хондроитинсульфат, дерматансульфат, кератансульфат, гепарин). Среди них различают:
Большой хондроитинсульфатсодержащий протеогликан. Предполагается, что этот протеогликан «захватывает пространство», которое должно стать костью, благодаря большому содержанию сульфата, в гидротированном состоянии способен занимать значительный объем пространства.
Декорин и бигликан очень сходны по строению, соответственно имеют один или два гликозаминогликана, белковая часть содержит 24 аминокислотных остатка, богатых лейцином. Несмотря на биохимическое сходство, эти белки отличаются по локализации. Локализация более распространенного декорина совпадает с расположением коллагена, что соответствует его функции «отделывать» молекулы коллагена и регулировать диаметр фибрилл. Бигликан сохраняется в матриксе.
К настоящему времени выделено много других типов протеогликанов, но это в основном белки клеточной поверхности, роль которых мало изучена.
На долю альбумина приходится большая часть неколлагеновых белков. По иммунологическим свойствам идентичен сывороточному.
Углеводы играют огромную роль в жизнедеятельности костной ткани, в процессах ее образования. На долю гликогена приходится 50-80мкг на 1г влажной ткани. Присутствие гликогена — необходимое условие процесса минерализации, он концентрируется главным образом на месте будущего центра минерализации. В костной ткани с большой интенсивностью протекают процессы гликолиза и пентозофосфатного пути.
Уровень нуклеиновых кислот зависит от функциональной активности. В покоящихся остеобластах количество РНК невелико, тогда как в пролиферирующих и гипертрофированных клетках повышено. Отмечено снижение содержания РНК при превращении остеобластов в остеоциты. ДНК находится в ядрах преостеобластов, остеобластов и остеокластов. Высокое содержание РНК отражает их активную и постоянную биосинтетическую функцию: образование большей массы костного матрикса.
Липиды играют важную роль в процессе минерализации и транспорта ионов через мембраны. Преобладают полярные липиды: фосфатидилхолин,
фосфатидилсерин, фосфатидилэтаноламин. Всего 0,61% липидов на сухую массу ткани.
Основная органическая кислота, находящаяся в костной ткани — цитрат. Её содержание в 230 раз превышает концентрацию в печени и достигает 90%. Активность цитратсинтетазы значительно выше активности ферментов, принимающих участие в распаде лимонной кислоты, а она, накапливаясь, принимает участие в регуляции уровня Са в сыворотке крови. Причем лимонная кислота находится в двух формах:
1. растворимой — принимает участие в цикле трикарбоновых кислот;
2. нерастворимой — неактивной, входящей в состав минерального компонента костной ткани.
В костной ткани активно преобладают метаболические процессы. Характерная особенность — аэробный гликолиз. Потребление глюкозы как в аэробных, так и в анаэробных условиях остеогенными клетками значительно больше, чем клетками печени, мышц и других органов.
Структура и функции костной ткани поддерживаются специфическими ферментами, синтезирующими и расщепляющими макромолекулярные компоненты органического матрикса кости и ферментами общих метаболических путей, снабжающих костные клетки энергией. Ферментам принадлежит важная роль в процессах минерализации и резорбции кости.
Следует отметить специфическую локализацию ферментов. В остеокластах проявляется более высокая активность дегидрогеназ, кислой фосфатазы, аминопептидазы, по сравнению с другими клетками. В то же время остеокласты не содержат щелочной фосфатазы. Высокая активность аденилатциклазы, пируваткиназы, фосфотрансфераз в зонах роста, где протекают процессы кальцификации.
Активность окислительных ферментов, таких как цитохромоксидаза, каталаза значительно ниже, чем, например, печени. Щелочная фосфатаза локализованная главным образом в остеобластах, в остеокластах вообще не обнаружена. Содержание этого фермента и его активность резко повышается в определенные сроки переломов кости, рахите и других патологий.
Кислая фосфатаза сосредоточена в остеокластах. Она непосредственно участвует в резорбции кости, осуществляя расщепление органических эфиров фосфорной кислоты с освобождением фосфатных ионов. Таким образом, кислая фосфатаза — лизосомный фермент и ее главная функция заключается в катаболизме, тогда как щелочная — принимает участие в процессах минерализации.
Основной белок костной ткани — коллаген, который содержится в количестве 15% — в компактном веществе, 24% — в губчатом веществе.
Костный коллаген — коллаген I типа — в нем больше, чем в других видах коллагена, содержится оксипролина, лизина и оксилизина, отрицательно заряженных аминокислот, с остатками серина связано много фосфата, поэтому костный коллаген — это фосфопротеин. Благодаря своим особенностям костный коллаген принимает активное участие в минерализации костной ткани.
В процессе жизнедеятельности костной ткани между ее компонентами и неорганическими ионами плазмы крови постоянно осуществляется обмен.
Костная ткань является депо минеральных компонентов, буферной системой, участвующей в поддержании концентрации ряда ионов. Она быстро поглощает из крови введенный Са, также быстро, за короткое время содержание Са в ней уменьшается на 20%.в костной ткани обнаруживаются различные соединения Са: кальцийфосфат, карбонат кальция, соединения с Cl, F.

Структура решетки неорганических кристаллов кости соответствует структуре кристаллов гидроксиапатита Ca10(PO4)6(OH)2 — это часть минеральной фазы кости, другая часть представлена аморфным фосфатом кальция. Он представляет плотную некристаллическую субстанцию в виде аморфных гранул, имеющих вид овалов или кругов, диаметром 5,0-20,0 нм. Является важным компонентом костной ткани, и его присутствие не зависит от анатомического строения кости, но подвержено значительным колебаниям в зависимости от возраста. Эта фаза преобладает в раннем возрасте, в зрелой же кости преобладающим становится кристаллический гидроксиапатит. Образование костной соли отражается общим уравнением:
Читайте также: Жаккардовая тюль что это за ткань
Растворению костной ткани способствует локальное повышение кислотности среды. При небольшом повышении содержания протонов водорода кость начинает растворяться, отдавая вначале катионы кальция:
При большей кислотности среды происходит полный ее распад:
Гидролиз аморфного кальцийфосфата обеспечивает постоянную концентрацию кальция в интерстициальной жидкости костной ткани.
В настоящее время известно более 30 микроэлементов: Си, Sr, Zn, Ba, Al, Be, Si, F и другие. Они необходимы для жизнедеятельности отеогенных клеток в процессе оссификации и декальцинации.
Обызвествление костной ткани и ее декальцинации находятся в тесной зависимости от содержания микроэлементов. Так, Sr и V способствуют обызвествлению, a Zn и Ва участвуют в регуляции процесса декальцинации. Mg активирует ряд ферментов, в частности, щелочную фосфатазу, участвующую в процессе минерализации.
Особого внимания заслуживает Sr. Его химические свойства близки к Са. Sr конкурирует с Са за место в кристаллической решетке, однако Sr удерживается в меньшей степени, чем Са, в том случае, если в рационе преобладает Са. При дефиците же Са в рационе Sr поглощается организмом в значительно больших количествах, чем в норме. Длительное поступление избыточных количеств Sr ведет к замещению им ионов Са в кристаллической решетке гидроксиапатита, в результате чего кости деминерализуются и деформируются.
В зрелом организме процессы минерализации и резорбция кости находятся в состоянии динамического равновесия. Минерализация — это формирование кристаллических структур минеральных солей костной ткани. Активное участие в минерализации принимают остеобласты. Для минерализации требуется много энергии (в форме АТФ), регулируемой многими факторами, включая ферменты, гормоны, витамины.
Решительный поворот в изучении минерализации начался с 1923г., вскоре после открытия в костной ткани фермента щелочной фосфатазы. Английский биохимик Р.Робинсон высказал предложение, что фосфорнокислый кальций откладывается там, где действует этот фермент. Однако щелочная фосфатаза содержится во многих тканях, не подвергающихся минерализации, и для того, чтобы произошло обызвествление необходимы другие факторы.
Позднее было доказано участие многих факторов: гликогена, ферментов гиколиза, АТФ, ЦТК, гликозаминогликанов.
Для приведенных всех теорий и некоторых экспериментальных данных общим является представление о ведущей роли ферментов, отщепляющий неорганический фосфат от органического субстрата. Концентрация фосфата в участках функционирования этих ферментов повышается, достигая уровня, при котором начинается его самопроизвольное осаждение, приводящее к кристаллизации. •
Дальнейшие исследования позволили предположить, что процесс кальцификации состоит в очаговом образовании центров кристаллизации гидроксиапатита из растворов Р и Са под действием коллагеновых волокон, в которых необходимо специфическое взаиморасположение реакционноспособных групп боковых аминокислотных цепей, способных служить центрами кристаллизации.
Важную роль в минерализации выполняют гликозаминогликаны, в частности хондроитинсульфат, которые обладают повышенным сродством к ионам Са и Р. подтверждением служат экспериментальные данные, демонстрирующие, что гликозаминогликаны интенсивно секретируются остеобластами в зоне минерализации, а затем подвергаются действию лизосомальных ферментов, образуя высокоактивные ионы.
Биохимическую основу нуклеации первичных зародышевых кристаллов составляет реакция образования комплекса между коллагеном, АТФ, Са и хондроитинсульфатом. К факторам, контролирующим кристаллообразование на волокнах коллагена относится также пирофосфат, который ингибирует минерализацию. Доказана также роль в этом процессе фосфолипидов, без которых органический матрикс костной ткани утрачивает способность обызвествляться.
Возрастные изменения костной ткани и основная патология.
В процессе онтогенетического развития костная ткань претерпевает выраженные изменения структурно-морфологического и биохимического характера. В ней происходит закономерное снижение содержания органических компонентов и нарастание минеральных. Эти изменения тесно связаны с обменом микроэлементов. Происходит накопление Sr, Pb, Si, A1, а концентрация Си уменьшается, интенсивность метаболизма фосфора и кальций уменьшается в десятки раз.
Один из ведущих возрастных изменений костной системы — развитие остеопороза — прогрессирующее системное заболевание скелета, характеризующееся снижением массы кости, нарушением структуры костной ткани, приводящее к увеличению хрупкости кости и риска переломов.
ПРЕДПРАСПОЛАГАЮЩИЕ К РАЗВИТИЮ ОСТЕОПОРОЗА
-принадлежность к европеоидной или монголоидной расам;
-семейная предрасположенность (нетравматические перело-
мы у родственников 1 степени родства;
— после 35-40 лет костная масса и всасывание Са в кишечнике снижается.
-низкое потребление Са и витамина D;
-низкая физическая активность;
-нарушение менструального статуса (длительная вторичная аменорея, преждевременная менопауза – до 45 лет, позднее начало менструаций);
-эндокринные болезни (первичный гиперпаратиреоз, тиреотоксикоз, сахарный диабет, синдром Кушинга);
-болезни крови (множественная миелома, системный масто-
-воспалительные ревматические заболевания (ревматоидный артрит, дерматомиозит, системная красная волчанка);
-заболевания ЖКТ (нарушения всасывания, болезнь Крона, хронические заболевания печени);
-хроническая почечная недостаточность;
-хронические неврологические заболевания.
I. Постменопаузный остеопороз (требует наблюдения в течение 15 лет с начала
менопаузы). Происходит разрежение костных балок, увеличивается частота
переломов позвонков и других костей.
П. Сенилъный остеопороз — характерен для мужчин старше 70 лет.
III. Вторичный остеопороз — на фоне глюкокортикоидной терапии синдрома
Кушинга, синдрома мальасорбции, нарушении питания, длительной
Предупредить остеопороз легче, чем лечить. Медикаментозные средства (кальцитонин, эстрогены, кальций, витамин D) способны лишь замедлить скорость потери кости, но, как правило, малоэффективны для восстановления уже утраченной костной массы.
Эстрогены — важное средство профилактики с наступлением менопаузы у женщин
Са — больным с остеопорозом показано 1000-1500 мг в день (с пищей поступает около 500 мг Са).
- Правообладателям
- Политика конфиденциальности