
Впервые А. Я. Данилевский (1881) разделил экстрагируемые из мышц белки на три класса: растворимые в воде, экстрагируемые 8 — 12% раствором хлорида аммония и белки, извлекаемые разбавленными растворами кислот и щелочей. В настоящее время белки мышечной ткани делят на три основные группы: саркоплазматические белки, миофибриллярные белки, белки стромы. На долю первых приходится около 35%, вторых —45% и третьих-20% всего мышечного белка. Эти группы белков резко отличаются друг от друга по растворимости в воде и солевых средах с различной ионной силой.
Белки, входящие в состав саркоплазмы, принадлежат к числу протеинов, растворимых в солевых средах с низкой ионной силой.
К числу саркоплазматических белков относятся также дыхательный пигмент миоглобин и разнообразные белки-ферменты, локализованные главным образом в митохондриях и катализирующие процессы тканевого дыхания, окислительного фосфорилирования, а также многие стороны азотистого и липидного обменов. Недавно была открыта группа саркоплазматических белков — парвальбумины, которые способны связывать ионы кальция.
К группе миофибриллярных белков относятся миозин, актин и актомиозин — белки, растворимые в солевых средах с высокой ионной силой, и так называемые регуляторные белки: тропомиозин, тропонин, α- и β-актинин, образующие в мышце с актомиозином единый комплекс. Перечисленные миофибриллярные белки тесно связаны с сократительной функцией мышц.

Миозин составляет 50-55% от сухой массы миофибрилл. Миозин обладает АТФазной активностью, т. е. способностью катализировать расщепление АТФ на АДФ и Н3РО4. Химическая энергия АТФ, освобождающаяся в ходе данной ферментативной реакции, превращается в механическую энергию сокращающейся мышцы. Молекула миозина имеет сильно вытянутую форму, длину 150 нм. Она может быть расщеплена без разрыва ковалентных связей на субъединицы: две тяжелые полипептидные цепи и несколько коротких легких цепей. Тяжелые цепи образуют длинную закрученную α-спираль («хвост» молекулы), конец каждой тяжелой цепи совместно с легкими цепями создает глобулу («головку» молекулы), способную соединяться с актином. Эти головки выдаются из основного стержня молекулы.

Толстые нити (толстые миофиламенты) в саркомере получены путем соединения большого ппчисла определенным образом ориентированных в ппппппппппппппппппппппппппппппппппппространстве молекул миозина.
20% от сухой массы миофибрилл. Известны две формы актина: глобулярный (Г-актин) и фибриллярный (Ф-актин) актин. Молекула Г-актина состоит из одной полипептидной цепочки, в образовании которой принимают участие 374 аминокислотных остатка. Ф-актин является продуктом полимеризации Г-актина и имеет структуру двухцепочечной спирали.
Актомиозин образуется при соединении миозина с Ф-актином. Актомиозин, как обладает АТФазной активностью. Однако АТФазная активность актомиозина отличается от АТФазной активности миозина. Фермент актомиозин активируется ионами магния и ингибируется этилен-диаминтетраацетатом (ЭДТА) и высокой концентрацией АТФ, тогда как миозиновая АТФаза ингибируется ионами Mg 2 + , активируется ЭДТА и не ингибируется высокой концентрацией АТФ. Оптимальные значения рН для обоих ферментов также различны.
Тропомиозин был открыт К. Бейли в 1946 г. Молекула тропомиозина состоит из двух α-спиралей и имеет вид стержня. На долю тропомиозина приходится около 4 — 7% всех белков миофибрилл.
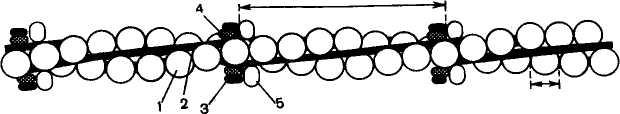
Т
Структура тонкого филамента.
1 — актин; 2 — тропомиозин; 3 — тропонин С;
ропонин — глобулярный белок, открытый С. Эбаси в 1963 г. В скелетных мышцах взрослых животных и человека тропонин (Тн) составляет лишь около 2% от всех миофибриллярных белков. В его состав входят три субъединицы (Тн-I, Тн-С, Тн-Т). Тн-I (ингибирующий) может ингибировать АТФазную активность, Тн-С (кальцийсвязывающий) обладает значительным сродством к ионам кальция, Тн-Т (тропомиозинсвязывающий) обеспечивает связь с тропомиозином. Тропонин, соединяясь с тропомиозином, образует комплекс, названный нативным тропомиозином. Этот комплекс прикрепляется к актиновым филаментам и придает актомиозину скелетных мышц позвоночных чувствительность к ионам кальция.
Белки стромы в поперечнополосатой мускулатуре представлены в основном коллагеном и эластином.
Небелковые азотистые экстрактивные вещества
В скелетных мышцах содержится ряд важных азотистых экстрактивных веществ: адениновые нуклеотиды (АТФ, АДФ и АМФ), нуклеотиды неаденинового ряда, креатин-фосфат, креатин, креатинин, карнозин, ансерин, свободные аминокислоты и др.
На долю креатина и креатинфосфата приходится до 60% небелкового азота мышц. Креатинфосфат и креатин относятся к тем азотистым экстрактивным веществам мышц, которые участвуют в химических процессах, связанных с мышечным сокращением.

К числу азотистых веществ мышечной ткани принадлежат и имидазолсодержащие дипептиды — карнозин и ансерин.
Карнозин и ансерин — специфические азотистые вещества скелетной мускулатуры позвоночных — увеличивают амплитуду мышечного сокращения, предварительно сниженную утомлением. Имидазолсодержащие дипептиды не влияют непосредственно на сократительный аппарат, но увеличивают эффективность работы ионных насосов мышечной клетки.
Среди свободных аминокислот в мышцах наиболее высокую концентрацию имеет глутаминовая кислота (до 1,2 г/кг) и ее амид — глутамин (0,8-1,0 г/кг). В состав различных клеточных мембран мышечной ткани входит ряд фосфоглицеридов: фосфатидилхолин, фосфатидилэтаноламин, фосфатидилсерин и др. Другие азотсодержащие вещества: мочевина, мочевая кислота, аденин, гуанин, ксантин и гипоксантин — встречаются в мышечной ткани в небольшом количестве.
Читайте также: Удаление крахмала с ткани
Белки входящие в мышечную ткань
Около 40% массы тела человека составляют скелегные мышцы, и, вероятно, 10% приходится на долю гладких мышц и мышцы сердца. Некоторые из основных принципов сокращения являются общими для разных типов мышц.
На рисунке показана схема организации скелетной мышцы. Все скелетные мышцы содержат множество волокон диаметром от 10 до 80 мкм. На рисунке также показано, что каждое из этих волокон состоит из последовательно соединенных более мелких элементов.
В большинстве скелетных мышц каждое волокно вытянуто во всю длину мышцы и, за исключением примерно 2%, обычно иннервируется лишь одним нервным окончанием вблизи середины волокна.
Сарколемма. Сарколеммой называют клеточную мембрану мышечного волокна. Она состоит из собственно мембраны клетки, называемой плазматической мембраной, и наружного покрытия из тонкого слоя полисахаридного материала, который содержит множество тонких коллагеновых нитей. На концах мышечного волокна этот поверхностный слой сарколеммы сливается с сухожильными волокнами. Сухожильные волокна, в свою очередь, собираются в пучки и формируют мышечные сухожилия, вплетающиеся в кости.
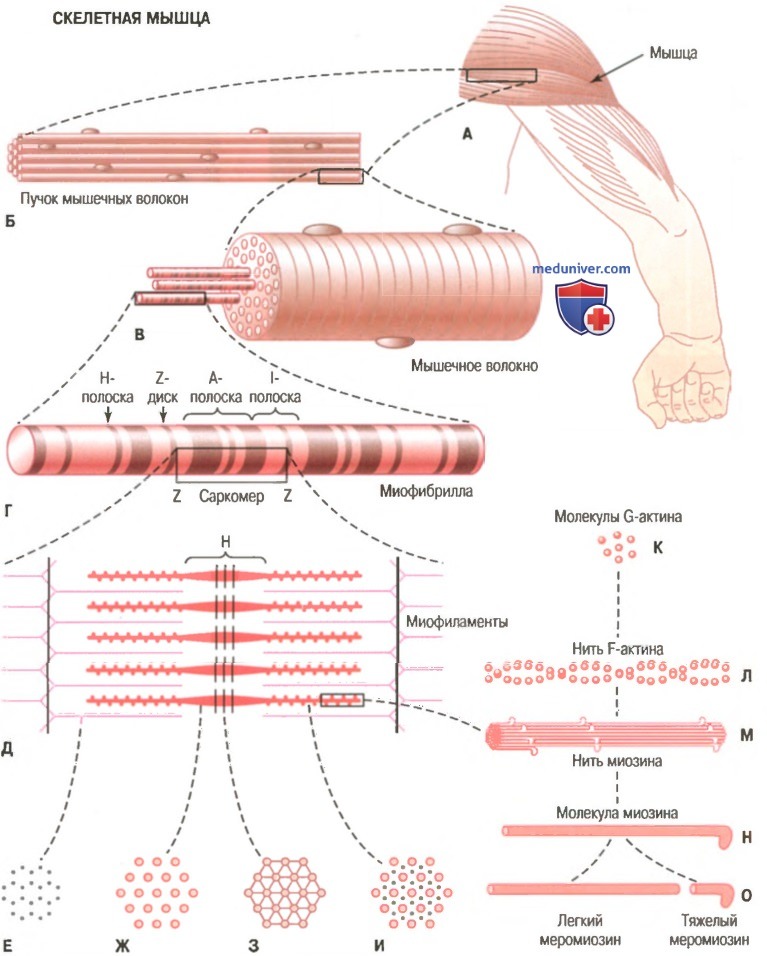
Организация скелетной мышцы от макроскопического до молекулярного уровня. Е, Ж 3, И — поперечное сечение на указанном уровне.
Миофибриллы. Нити актина и миозина. Каждое мышечное волокно содержит от нескольких сотен до нескольких тысяч миофибрилл, которые представлены на поперечном срезе в виде небольших светлых точек. Каждая миофибрилла собрана примерно из 1500 смежных нитей миозина и 3000 нитей актина, представляющих собой большие полимеризованные белковые молекулы, ответственные за мышечное сокращение. Они видны в продольном сечении на электронной микрофотографии. Толстые нити на схемах — миозиновые, тонкие — актиновые.
Обратите внимание, что на рисунке актиновые и миозиновые нити частично перекрываются, в связи с чем миофибриллы имеют чередующиеся светлые и темные полоски. Светлые полоски содержат только актиновые нити, их называют I-полосками, поскольку они изотропны в поляризованном свете. Темные полоски содержат миозиновые нити, а также концы актиновых нитей, которые частично накладываются на миозиновые нити. Эти полоски называют А-полосками, поскольку они анизотропны в поляризованном свете. На рисунке видны также маленькие выступы по сторонам миозиновых нитей. Это так называемые поперечные мостики. Их взаимодействие с актиновыми нитями лежит в основе сокращения мыщц.
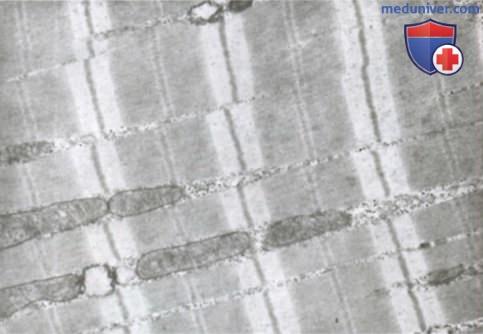
Электронная микрофотография миофибриллы, демонстрирующая детальную организацию актиновых и миозиновых нитей. Видны митохондрии, лежащие между миофибриллами.
На рисунке показано также, что концы актиновых нитей прикреплены к так называемому Z-диску (Z-мембране). Отсюда актиновые нити распространяются в обе стороны, проходя между миозиновыми нитями. Z-диск, состоящий из нитевидных белков, отличающихся от актиновых и миозиновых нитей, проходит в поперечном направлении через миофибриллу, а также через все мышечное волокно, переходя от одной миофибриллы к другой, скрепляя их между собой. В связи с этим целое мышечное волокно, как и отдельные миофибриллы, имеет светлые и темные полоски. Эти полоски придают скелетным и сердечной мышцам характерный полосатый вид.
Часть миофибриллы (или целого мышечного волокна), расположенную между двумя последовательными Z-дисками, называют саркомером. В сокращенном мышечном волокне длина саркомера равна примерно 2 мкм. При этой длине актиновые нити полностью перекрывают миозиновые, и кончики актиновых нитей начинают перекрывать друг друга. Мы увидим далее, что при этой длине мышца способна генерировать максимальную силу сокращения.
Что удерживает актиновые и миозиновые нити на месте? Удерживать актиновые и миозиновые нити рядом довольно трудно. Это обеспечивается с помощью большого числа нитевидных молекул белка, называемого тайтином. Каждая молекула тайтина имеет молекулярную массу около 3000000, что делает ее одной из самых крупных белковых молекул в организме. Кроме того, поскольку молекула тайтина волокнистая, она очень упруга. Упругие молекулы тайтина образуют каркас, удерживающий актиновые и миозиновые нити в положении, обеспечивающем нормальную работу сократительного аппарата саркомера. Это позволяет полагать, что сама молекула тайтина действует как матрица для начального формирования участков сократительных нитей саркомера, особенно миозиновых нитей.
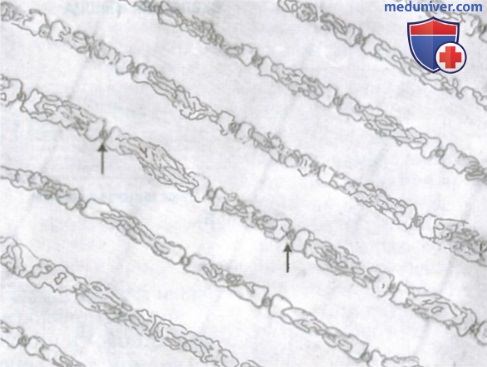
Саркоплазматический ретикулум во внутриклеточных пространствах между миофибриллами с системой продольных трубочек, параллельных миофибриллам. Показаны также в поперечном сечении Т-трубочки (стрелки), которые связаны с внеклеточным пространством и отвечают за проведение электрического сигнала в центр мышечного волокна.
Саркоплазма. В каждом мышечном волокне многочисленные миофибриллы расположены рядом, бок о бок. Пространство между ними заполнено внутриклеточной жидкостью, называемой саркоплазмой и содержащей большое количество калия, магния и фосфатов, а также множество ферментов. Кроме того, имеется огромное число митохондрий, расположенных параллельно миофибриллам. Они снабжают сокращающиеся миофибриллы большим количеством энергии в форме аденозинтрифосфата, синтезируемого митохондриями.
Саркоплазматический ретикулум. В саркоплазме, окружающей миофибриллы каждого мышечного волокна, имеется также хорошо развитый ретикулум, называемый саркоплазматическим ретикулумом. Он имеет специфическую организацию, исключительно важную для регуляции мышечного сокращения . Саркоплазматический ретикулум особенно сильно развит в очень быстро сокращающихся типах мышечных волокон.