
Миозины представляют собой белковые моторы, использующие энергию гидролиза связанного АТФ для генерации силы, обеспечивающей подвижность вдоль актиновых филаментов. Миозины хорошо известны из-за своей роли в мышечном сокращении. Однако экспрессия миозина не ограничивается только мышцами. Эта большая группа белков, которые экспрессируются практически во всех клетках эукариот.
Различные ее представители характеризуются структурными и биохимическими особенностями, которые предназначаются для специфических клеточных нужд.
Миозиновая группа молекулярных моторов на основе актина состоит по меньшей мере из 18 классов (или семейств), причем во многих классах белки имеют несколько изоформ. Классы различаются по результатам филогенетического анализа аминокислотных последовательностей. Все миозины, за исключением миозина VI, движутся по направлению к оперенному концу актиновых филаментов. Типы и количество миозинов, которые экспрессируются в данной клетке или организме, существенно различаются.
Например, в клетках дрожжей Saccharomyces cerevisiae экспрессируются пять миозинов, принадлежащих к трем разным классам, а у человека присутствуют сорок генов из двенадцати семейств.
Представители семейства миозинов участвуют в выполнении существенных и разнообразных клеточных функций. Для них характерно наличие трех общих доменов (головной или моторный домен, регуляторный и хвостовой домены), которые эволюционировали таким образом, что приобрели характеристики, позволяющие им выполнять в клетке различные специфические механические и регуляторные функции. Моторный и регуляторный домены управляют подвижностью, хвостовой домен участвует в полимеризации миозина и в связывании с другими клеточными компонентами, необходимыми для транспорта.
Достигнут большой прогресс в выяснении клеточных функций миозинов и их регуляции с участием моторных и хвостовых доменов. В таблице ниже представлены функции различных семейств миозина, другая таблица ниже суммирует свойства миозиновых доменов, а последняя таблица ниже содержит сведения об экспрессии миозинов человека. Все известные миозины можно подразделить на четыре больших функциональных группы.
Миозины, обеспечивающие мышечное и клеточное сокращение. Изоформы семейства миозин II генерируют усилия, вызывающие сокращение скелетных, сердечных и гладких мышц. Они также обеспечивают функционирование сократительного кольца при цитокинезе, миграцию клеток и прочие процессы, связанные с сократительной способностью клетки.
Миозины, обеспечивающие процессы мембранного и везикулярного транспорта. Моторы микротрубочек осуществляют цитозольный транспорт мембранных везикул на дальние расстояния. Однако некоторые миозины играют существенную роль в транспорте на короткие расстояния и в регуляции распределения везикул и органелл. Например, процессивный мотор миозин-V транспортирует пигментные органеллы, обеспечивающие окраску кожи и волос, в то время как другие миозины, например изоформы миозина-I, -VI, -IX, и -X, участвуют в формировании и транспорте везикул при эндоцитозе и фагоцитозе Миозины, играющие ключевую роль в регуляции формообразования и полярности клеток.
Миозины необходимы для образования и функционирования богатых актином специализированных поверхностных образований, таких как филоподии, стереоцилии и псевдоподии. Например, некоторые изоформы миозина-I связывают липиды мембраны с актиновым цитоскелетом и обеспечивают сокращение мембранных протрузий, богатых актином; изоформы миозина-П обеспечивают сокращение стресс-фибрилл и актиновых филаментов кортикального слоя клеток, участвующих в поддержании формы клетки; миозин-VII обеспечивает сокращаемую связь между актиновым цитоскелетом и внеклеточными матриксами.
Читайте также: Ткань для плаща название ткани
Миозины, участвующие в процессах передачи сигнала и реализации сенсорных ощущений. Миозин участвует в процессе передачи сигнала за счет ассоциации с сигнальными белками. Например, миозин-I регулирует активность ряда кальциевых каналов, миозин III взаимодействует с сигнальными молекулами фоторецепторов глаза, миозин-IX, по-видимому, регулирует активность Rho, и миозин-XVI направляет фосфатазу в специфические области клетки. Также миозины играют важную роль в реализации сенсорных ощущений. Известны мутации в генах, кодирующих синтез миозинов-VI, -VII и -XV. Они служат причиной потери слуха, которая происходит из-за нарушений в актин-содержащих структурах клеток сенсорных волосков внутреннего уха.
В последующих статьях на сайте мы рассмотрим структуру миозина, основные механизмы генерации усилий для всех охарактеризованных миозинов, и связь этих свойств с биологической ролью содержащих их клеток.
 Свойства 18 семейств миозинов.
Свойства 18 семейств миозинов.
Перечислены организмы, в которых они экспрессируются, и функции этих белков.
Для некоторых миозинов функции неизвестны. 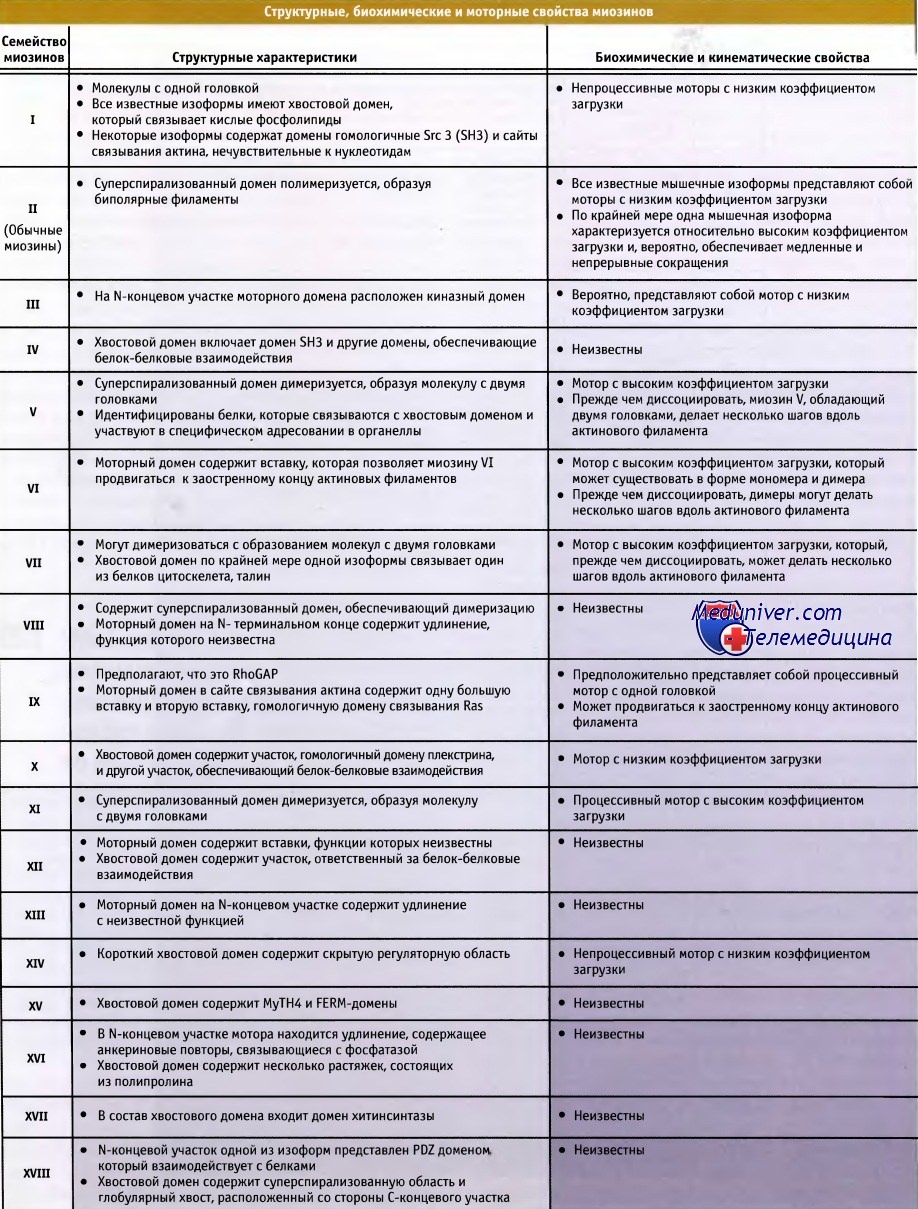 Структурные и кинематические свойства семейств миозинов
Структурные и кинематические свойства семейств миозинов 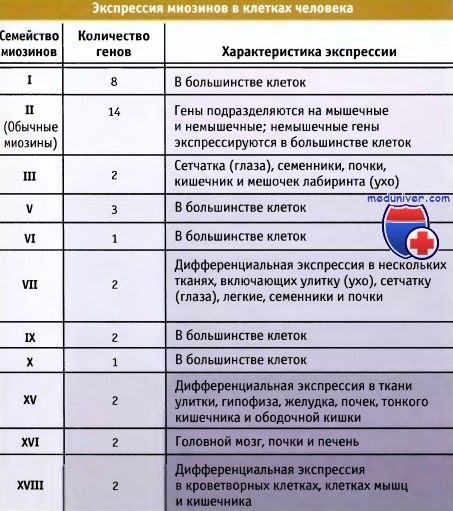 Экспрессия белков семейств миозинов в клетках человека
Экспрессия белков семейств миозинов в клетках человека
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Белок миозин в каких тканях

МИОЗИН (от греч. mys, род. падеж myos-мышца), белок сократит. волокон мышц. Его содержание в мышцах ок. 40% от массы всех белков (в др. тканях и клетках 1-2%). Молекула М. представляет собой длинный фибриллярный стержень (хвост), несущий на одном конце две глобулярные головки (рис. 1). Длина хвоста ок. 160 нм, диаметр 3 нм.
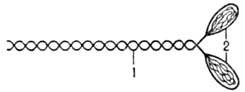
Рис. 1. Схема молекулы миозина: 1-фибриллярный стержень; 2-головки .
Головки имеют грушевидную форму: длина их составляет ок. 20 нм, а толщина-9,5 нм на конце и 5,5 нм в месте прикрепления к стержневой части. Последняя состоит из двух полипептидных цепей с мол. м. ок. 200 тыс. у каждой (т.наз. тяжелые цепи), закрученные спирально одна вокруг другой. В области головок с тяжелыми цепями ассоциированы легкие цепи-субъединицы с мол.м. ок. 20 тыс.; на каждую головку приходится две легкие цепи. Мол.м. всей молекулы составляет ок. 480 тыс.
При расщеплении М. кратковременным действием трипсина образуются два фрагмента, к-рые наз. легким и тяжелым меромиозинами (обозначаются соотв. LMM и НММ). LMM представляет собой фрагмент стержневой части молекулы М. длиной ок. 80 нм и с мол.м. 150 тыс. НММ содержит головку М. и часть хвоста, его длина 60-70 нм, мол. м. ок. 340 тыс. При действии папаином или при длит. воздействии трипсина от молекулы М. отщепляются головки (т. наз. субфрагменты 1, или S 1 с мол. м. 120 тыс. Стержневой фрагмент НММ, к-рый отщепляется при действии папаина, наз. субфрагментом 2 ( S 2 ).
N-Концевые части тяжелых цепей М. располагаются в головках. Особенность аминокислотного состава тяжелых цепей — наличие остатков метилир. аминокислот: 3-метил-гистидина, N 6 -моно- и N 6 -триметиллизина. Содержание a-спиралей в головках и хвосте молекулы составляет соотв. 33 и 94%. Тяжелые цепи имеют два гибких «шарнирных» участка: один в основании головки, другой на расстоянии 43 нм от первого.
М. обладает АТФазной активностью; катализирует гидролиз АТФ до аденозиндифосфата и Н 3 РО 4 (это св-во М. открыто В.А. Энгельгардтом и М.Н. Любимовой в 1939). Активный центр АТФазы М. находится в головках и сохраняет свои каталитич. св-ва при их протеолитич. отщеплении. Существенными для проявления активности являются e-аминогруппа лизина, гуанидиновые группировки аргинина, карбоксильные группы глутаминовой и аспарагиновой к-т.
В мышцах М. взаимод. с актином с образованием а к т о-м и о з и н о в о г о к о м п л е к с а, играющего важную роль в механизме сокращения мышц. Центр, ответственный за соединение с актином, также находится в головке. Соединение М. с актином в присут. ионов Mg 2+ сопровождается многократным увеличением АТФазной активности. При низкой ионной силе в присут. АТФ происходит р-ция суперпреципитации, в результате к-рой актомиозиновый комплекс сжимается и освобождает воду.
Читайте также: Древесина луб пробка кожица сердцевина функции ткани таблица
Одно из осн. св-в М.-его способность при низкой ионной силе агрегировать с образованием хорошо упорядоченных филаментов, подобных тем, в виде к-рых он присутствует в мышце. Нативный филамент М. представляет собой веретенообразный агрегат диаметром ок. 15 нм и длиной 1500 нм. Хвосты молекул М. упакованы в глубь филамента, а головки выходят наружу в виде выступов, регулярно расположенных на пов-сти. В центре филамента молекулы М. агрегированы хвост к хвосту, в результате чего образуется зона, не несущая головок. По обеим сторонам от центра филамента молекулы М. агрегированы полярно (хвост к голове). Один филамент содержит 300-400 молекул М.
В механизме мышечного сокращения важное значение имеют еще два белка-тропомиозин и тропонин. Молекула первого (мол. м. 67 тыс.) полностью построена из a-спиралей и состоит из идентичных по первичной структуре фрагментов, содержащих по 42 аминокислотных остатка. В бессолевой среде тропомиозин полимеризуется, образуя вязкую структуру, обладающую двойным лучепреломлением. При взаимод. с F-актином молекула тропомиозина укладывается в бороздки, образованные двойной спиралью актина. Молекула тропонина представляет собой комплекс, состоящий из трех белков,-тропонина Т (мол. м. 37 тыс.), тропонина I (мол. м. 25 тыс.) и тропонина С (мол. м. 20 тыс.). Тропонин I-ингибитор актомиозиновой Mg-АТФазы, тропонин С способен к связыванию ионов Са 2+ , тропонин I связывается с актином, тропонин Т-с тропо-миозином.
Мышечные волокна построены из продольно расположенных фибрилл (миофибрилл) диаметром ок. 1000 нм, в к-рых чередуются светлые и темные диски (соотв. I и А-диски; рис. 2). В середине диска I расположена пластинка Z, к-рая пронизывает все мышечное волокно, удерживая миофибриллы в пучке и упорядочивая их расположение. Участок между двумя соседними пластинками Z (т. наз. c a p к о м е р)- функцией, единица миофибрилл. Внутри миофибрилл находятся два вида филаментов, ориентированных параллельно продольной оси мышцы,-филаменты актина и миозина.
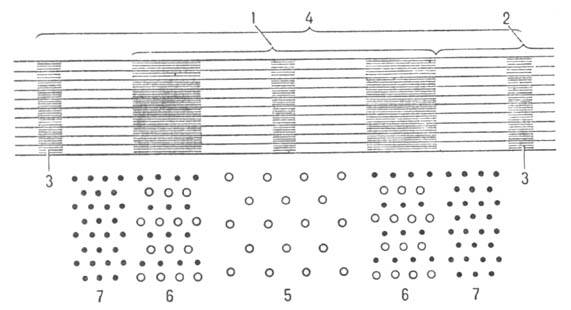
Рис. 2. Схема продольного разреза участка миофибриллы (1 -диск А, 2-диск I, 3-пластинка Z, 4-саркомер); внизу показана схема поперечного среза миофибриллы (5-только нити миозина, 6-нити актина и миозина, 7-только нити актина ).
Сокращение мышцы осуществляется в результате скольжения филаментов актина вдоль филаментов М. (рис. 3) благодаря периодич. присоединению головок М. к актино-вым филаментам и сгибания молекулы М. в шарнирных участках (происходит «проталкивание» актиновых филаментов). Расслабление происходит в результате разрыва поперечных контактов М. и актина и возвращения филаментов в исходное положение.
Такой цикл инициируется поступлением сигнала от нервных окончаний в мышечном волокне, к-рый обусловливает выброс Са 2+ из саркоплазматич. ретикулума (сложная система ограниченных мембранами пузырьков, трубочек и т. наз. цистерн, ориентированных вдоль миофибрилл и пронизывающих мышечные волокна). Ионы Са 2+ связываются с тропонином С, агрегированным с актином, и вызывают изменение конформации этого белка. В результате этого тропомиозин перемещается в сторону и не препятствует образованию контактов между головками М. и актином. Когда состояние возбуждения заканчивается, Ca 2+ -Mg 2+ -АТФаза саркоплазматич. ретикулума обусловливает обратный поток Са 2+ во внутр. пространство ретикулума. Когда концентрация Са 2+ достигает достаточно низкого уровня, контакты между головками М. и актином разрываются и мышечное волокно расслабляется. Необходимая энергия для осуществления этого процесса поступает в результате гидролиза АТФ на М.
Читайте также: Что можно сделать из остатков трикотажной ткани
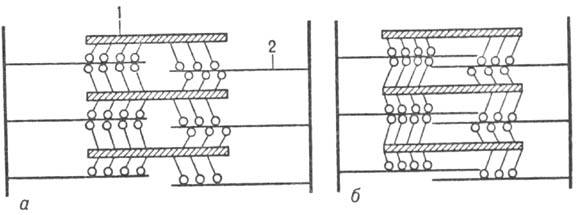
Рис. 3. Схема поперечнополосатой мышцы в покое ( а )и при ее сокращении ( б ); 1 — филаменты миозина; 2 — филаменты актина .
Лит.: Поглазов Б.Ф., Левицкий Д. И., Миозин и биологическая подвижность, М., 1982. Б. Ф. Поглазов .
Миозин в мышечных волокнах
Дана характеристика молекулы миозина: длина, толщина и молекулярная масса. Описана также структура молекулы миозина: головок и хвоста. Подробно описано формирование толстого филамента путем самосборки и факторы, отрицательно влияющие на этот процесс.
Миозин в мышечных волокнах
Основу толстого филамента саркомера (миофибриллы) составляет белок миозин. Впервые белок миозин был выделен из скелетных мышц в начале XX века.
Характеристика молекулы миозина
Молекула белка миозина (миозина II типа), который участвует в сокращении саркомера, имеет молекулярную массу 500 кДа. Длина молекулы равна 150 нм, а толщина – 2 нм. Молекула миозина образована четырьмя полипептидными цепями, две из которых (тяжелые) имеют молекулярную массу по 200 кДа, а две – легкие 20 и 17 кДа.
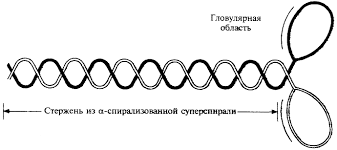
Рис. 1. Состав и структура молекулы миозина
Состав и структура молекулы миозина
Молекула миозина состоит из «хвоста» или «стержня» и двух «головок» (рис.1). Хвост миозина состоит из двух цепей, каждая из которых свернута в α-спираль. Две α-спирали дополнительно скручены в суперспираль. Эта суперспираль является стержнем (хвостом) миозиновой молекулы. Ее длина равна 135 нм.
На верхнем конце стержня две нити расходятся, сплетаются с дополнительными двумя легкими цепями и формируют две миозиновые головки. За счет головок длина миозиновой молекулы достигает 150 нм , а толщина – 2 нм. По этим характеристикам молекула миозина превосходит все молекулярные структуры, имеющиеся в природе.
Соединение между головкой и стержнем молекулы миозина подобно шарниру. Это позволяет головкам наклоняться и прикрепляться к молекулам актина.
АТФ-азная активность миозина
В 1939 году советскими учеными В.А. Энгельгардом и М.Н. Любимовой было установлено, что молекула миозина обладает АТФ-азной активностью, то есть участвует в реакции гидролиза АТФ – основной реакции энергообеспечения мышечной деятельности энергией. Позже было установлено, что на каждой миозиновой головке имеются центры. В одном из них находится фермент АТФ-аза. Другой центр предназначен для связывания головки миозина с тонким филаментом (рис.2).

Рис.2. Молекула миозина состоит из двух длинных полипептидов (тяжелых цепей), формирующих две глобулярные головки и стержнеобразный хвост, участвующий в образовании толстого филамента. Головки содержат актин-связывающие участки (А) и моторные области (М), где происходит связывание и расщепление АТФ. Вблизи головок с тяжелыми цепями ассоциированы две пары легких цепей, одна из которых содержит участок фосфорилирования (Ф).
В настоящее время на основе степени активности фермента АТФ-азы производится разделение мышечных волокон на типы.
Формирование толстого филамента
Несколько сотен миозиновых молекул способны самопроизвольно собираться в пучок. При таком объединении молекулы миозина располагаясь параллельно друг другу с некоторым сдвигом образуют своеобразный «ствол» (рис.2). Толщина «ствола» составляет 11-14 нм. В середине «ствола» переплетаются хвосты молекул миозина, а с двух концов «ствола» торчат головки. Это и есть толстый филамент (рис. 3 и рис.4).
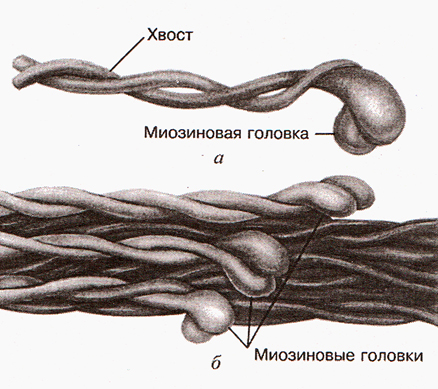

Рис.3. Молекула миозина и толстый филаментРис.4. Толстый филамент
Установлено, что агрегация (самосборка) толстого филамента очень чуствительна к рН саркоплазмы. Если рН саркоплазмы находится вне физиологических значений (рН = 7,2-7,5), самосборка толстых филаментов происходит нерегулярно и возникают очень длинные разветвляющиеся структуры (Дж. Бендолл, 1970). Из этого следует, что синтез белков, пока в мышечных волокнах много молочной кислоты (лактата) отставлен, так как могут синтезироваться «неправильные» толстые филаменты, саркомеры и миофибриллы.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом