
Микротрубочки, актиновые филаменты (микрофиламенты) и промежуточные филаменты представляют собой три основные системы белковых филаментов, составляющих цитоскелет. Промежуточные филаменты образуют в цитоплазме и ядре сеть и присутствуют во всех клетках метазоа (животных).
В отличие от микротрубочек и актиновых филаментов, которые необходимы даже для выживания изолированных клеток in vitro, основная функция промежуточных филаментов проявляется на уровне тканевой организации, где они необходимы для надлежащего функционирования тканей и органов. Некоторые типы промежуточных филаментов участвуют в скреплении клеток друг с другом, что необходимо для формирования тканей.
Белки промежуточных филаментов кодируются несколькими большими семействами генов. Эти белки образуют сложную систему филаментов, на долю которых в клетке в нормальных физиологических условиях приходится до 80% общего клеточного белка. Внутриклеточное распределение промежуточных филаментов отличается от характерного для актиновых филаментов и микротрубочек.
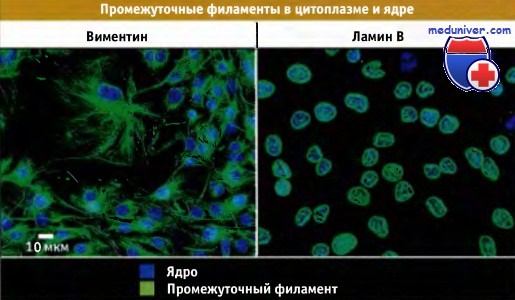
Распределение различных типов промежуточных филаментов в культивируемых фибробластах.
Иммунофлуоресцентное окрашивание на виментин и ламин В. Виментин находится в цитоплазме, а ламины в ядре.
Гистологи обнаружили их (в виде нейрофибрилл нейронов и тонофиламентов клеток эпидермиса) задолго до того, как в 1960-х гг. при электронно-микроскопическом исследовании мышечной ткани были описаны индивидуальные филаменты. В клетках мышц «промежуточные» филаменты занимали по диаметру среднее положение между «толстыми филаментами» миозина-II и «тонкими филаментами» актина. Их средний диаметр составляет около 10 нм, т. е. они толще, чем актиновые филаменты (около 8 нм), и тоньше микротрубочек (около 25 нм). Все три системы филаментов представлены рисунке ниже.
Белки промежуточных филаментов характеризуются общей молекулярной структурой и полимеризуются в филаменты, обладающие высокой механической прочностью. В электронном микроскопе они выглядят одинаково. У высших позвоночных семейство соответствующих белков организовано наиболее сложно, и этот вопрос будет рассмотрен в настоящей главе.
Похожие промежуточные филаменты также обнаружены у беспозвоночных, однако у них количество генов, кодирующих соответствующие белки, значительно меньше, чем у позвоночных. Также промежуточные филаменты беспозвоночных менее гетерогенны и обладают меньшей тканевой специфичностью, чем у млекопитающих. В геноме человека находится порядка 70 генов, кодирующих белки промежуточных филаментов. Принимая во внимание альтернативный сплайсинг для пары из них, общее количество этих белков приближается к 75.
Они представлены гораздо большим числом вариантов и более гетерогенны, чем актиновые или тубулиновые белки. Для всех белков промежуточных филаментов характерна тканеспецифическая экспрессия. Также их экспрессия изменяется в процессе дифференцировки.
Большинство сведений, касающихся экспрессии и биохимических свойств промежуточных филаментов, были получены до того, как были установлены их функции и связь с некоторыми заболеваниями. Сейчас показано, что мутации в генах белков промежуточных филаментов связаны с многими генетическими заболеваниями, которые характеризуются различными фенотипическими проявлениями. Они включают по меньшей мере 50 отдельных болезней, от фликтены до прогерии.
Читайте также: Джинсов куртки из джинсовой ткани
Почти все типы генов белков промежуточных филаментов связаны с той или иной формой проявления хрупкости тканей. Это позволяет предполагать, что для функционирования ткани in vivo необходима надлежащая механическая прочность и что в значительной степени она прямо или опосредованно связана с промежуточными филаментами. Принимая во внимание, что экспрессия генов белков промежуточных филаментов носит тканеспецифический характер, весьма возможно, что все эти белки придают клеткам тканей мельчайшие оттенки различия. Клеткам тканей необходимы различные свойства, такие как прочность, пластичность, быстрота сборки и разборки структур, обеспечивающих прочность.
Может быть, в этом кроется причина того, что в ходе эволюции возникло столь много генов, кодирующих белки промежуточных филаментов.

Основные компоненты цитоскелета в электронном микроскопе.
На ультратонком срезе клетки эпителия почки видны актиновые микрофиламенты, промежуточные филаменты К8/К18 и микротрубочки.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Белок промежуточные филаменты мышечных тканей
• Некоторые белки промежуточных филаментов, относящиеся к типу III и IV, ко-экспрессируются в разных тканях
• Многие белки типа III и IV образуют друг с другом сополимеры
• Совместная экспрессия множественных типов белков промежуточных филаментов может маскировать эффект мутаций в белке, принадлежащим к одному из типов
• Десмин представляет собой необходимый белок мышц
• Виментин часто экспрессируется в одиночных клетках
• Мутации в генах белков типа III или IV обычно связаны с мышечными или неврологическими расстройствами, носящими дегенеративный характер
В таблице ниже перечислены гомологичные некератиновые белки промежуточных филаментов, которые относятся к группам III—IV. В данной статье мы рассмотрим белки промежуточных филаментов, относящихся к типам III и IV. Эти типы различаются по принципу гомологии по определенным последовательностям. К белкам III типа относятся десмин, виментин, фибриллярный кислый белок глиального происхождения (GFAP, фибриллы которого представлены на рисунке ниже) и периферии.
Эти белки дифференциально экспрессируются в клетках соединительной ткани, мышц, в нейронах и некоторых других дифференцированных клетках. Их экспрессия индивидуально регулируется для каждого белка и тесно связана со специфическими процессами дифференцировки. Некоторые из этих белков одинаково экспрессируются в разных тканях, а другие могут образовывать друг с другом сополимеры. Однако они никогда не образуют таких комплексов с кератинами (тип I/II) или с ламинами (тип V).
Так же как и для других белков промежуточных филаментов, мы пока точно не знаем, каким образом вариации в аминокислотной последовательности этих белков связаны с их специфическими функциями в клетках различных типов, хотя мутации в них хорошо коррелируют с различными нарушениями в функционировании соответствующей ткани.
Десмин представляет собой белок, необходимый для функционирования клеток мышц всех типов (поперечнополосатых, сердечной и гладких); подобно кератину эпителиальных клеток, десмин обеспечивает упругость ткани при физических воздействиях. Десминовые филаменты проходят между точками скрепления сократительных структур (например, саркомер поперечнополосатых мышц). Эти места в клетках мышц являются сайтами приложения значительных механических усилий. Поэтому десмин можно рассматривать как белок, обеспечивающий устойчивость мышечных клеток к механическому воздействию.
Такая точка зрения возникла после того, как оказалось, что ткани, в клетках которых десмин отсутствует или находится в мутантной форме, теряют способность противостоять механическим воздействиям. Десмин образует комплексы с другими белками типа III; в некоторых клетках он сополимеризуется также с белками типа IV.
Читайте также: Как сделать ламбрекен из плотной ткани
В отличие от десмина, который экспрессируется в тканях состоящих из плотных агрегатов клеток, таких как мышечная и эпителиальная, экспрессия виментина типична для клеток, которые функционируют изолированно, или в виде рыхло-связанных клеточных ассоциатов или слоев. В процессе развития виментин экспрессируется после кератинов и остается во многих мезенхимальных и соединительнотканных клетках взрослого организма, от фибробластов до кроветворных клеток и клеток эндотелия сосудов, а также некоторых видов эпителия.
Астроциты и глиальные клетки представляют собой клетки центральной нервной системы, отличные от нейронов. Они необходимы для роста, дифференцировки и регенерации последних. Все астроглиальные клетки экспрессируют GFAP, который обычно экспрессируется вместе с виментином или белком нестином, относящимся к группе IV. Такая совместная экспрессия защищает клетки от последствий мутаций или утраты одного из генов. Эксперименты по двойному нокауту генов, проведенные на животных, показали, что эти промежуточные филаменты типа III необходимы для нормального функционирования астроцитов в центральной нервной системе.
Эти клетки необходимы для заживления ран и развития устойчивости к осмотическому шоку. Для нормального функционирования астроцитам необходимо сформировать отростки, которые при отсутствии промежуточных филаментов несут различные дефекты.
Периферии, главным образом, экспрессируется в клетках периферической нервной системы. При формировании аксонами отростков они вначале синтезируют периферии и виментин; затем этот синтез сменяется экспрессией триплета белков нейрофиламентов (NF-L, NF-M, и NF-H). Впрочем, после повреждения нервной ткани экспрессия периферина быстро возобновляется. По-видимому, для нормального выполнения белком своих функций важен его уровень. Так, в экспериментах на животных показано, что слишком высокое содержание периферина приводит к летальным последствиям, а его отсутствие — к утрате некоторых небольших сенсорных аксонов.
Филаменты астроцитов спинного мозга, состоящие из глиального фибриллярного кислого белка (GFAP).
В настоящее время гомологичный тип IV включает триплет белков нейрофиламентов низкой, средней и высокой молекулярной массы (NF-L, NF-M и NF-H), а также а-интернексин, нестин, синкоилин и синемин. В тканях большинство белков типа IV присутствуют в гетерополимерной форме, и гораздо более эффективно образуют сополимеры с белками типа III или другими белками, относящимися к типу IV. В зрелых нейронах, белки триплета нейрофиламентов почти всегда экспрессируются вместе, в то время как синкоилин и два продукта альтернативного сплайсинга синемина (а и b) преимущественно экспрессируются в клетках мышц и сополимеризуются с белками группы III.
Согласно иммуногистохимическим данным, синкоилин и синемин, так же как и десмин, локализованы в мышечных клетках, в точках приложения нагрузки. Возможно, что эти белки промежуточных филаментов тоже обеспечивают устойчивость клеток к нагрузкам.
Необычайная длина аксональных выростов в нейронах (у нейронов седалищного нерва человека она может достигать 1 м) свидетельствует о важности поддержания соответствующего уровня экспрессии белков нейрофиламентов, необходимых для укрепления цитоплазмы. В процессе развития экспрессия нейрофиламентов происходит по сложной программе, которая включает одновременный синтез многих белков. Соответствующие профили накладываются и смещаются относительно друг друга. Таким образом, нейроны постоянно экспрессируют промежуточные филаменты. Первым экспрессируются нестин и виметин. (Подобно GFAP астроглии, при раневых повреждениях нейрональный нестин может экспрессироваться повторно.)
Читайте также: Синдромы при повреждении мягких тканей
По мере удлинения выростов, доминирующим становится синтез а-интернексина, который сменяется NF-L, и наконец, происходит экспрессия крупных белков нейрофиламентов. В стабилизации сформировавшихся аксонов и нейритов важную роль играет триплет белков нейрофиламентов.
У нескольких белков, относящихся к группе IV, имеются длинные хвостовые домены, которые участвуют в организации каркасных структур цитоплазмы. В хвостовом домене NF-H присутствует ряд повторов, представляющих собой последовательность Lys-Ser-Pro, которая является сайтом фосфорилирования. Когда происходит фосфорилирование этих повторов, хвостовой домен приобретает высокий положительный заряд и ориентируется под прямым углом к оси филамента. Это способствует растяжению цитоплазмы аксона. У мышей число аксонов и их диаметр коррелирует с уровнем экспрессии белков нейрофиламентов.
Большему диаметру аксона соответствует более быстрая проводимость нервного импульса, что является особенно важным фактором в эволюции крупных по размеру тел позвоночных.
Мутации в генах белков группы III связаны с развитием многочисленных патологических состояний, механизм которых в некоторых случаях удалось выяснить в экспериментах на животных. У человека мутации в генах GFAP связаны со смертельным нейродегенеративным заболеванием, болезнью Александера. В экспериментах на животных показано, что в отсутствие GFAP наблюдается аномальная реакция астроглии на повреждения, и астроциты не образуют цитоплазматических выростов. В десмине обнаружено много патологических мутаций, которые происходят в разных участках белковой структуры.
У человека мутации в гене десмина связаны с дефектами сердечно-сосудистой системы, в особенности с развитием сердечной недостаточности на почве дилатационной кардиомиопатии, а также с некоторыми формами мышечной дистрофии. У мышей при отсутствии десмина стенки важнейших кровеносных сосудов становятся слишком мягкими и не могут поддерживать надлежащий уровень кровяного давления. Это вызывает растяжение уже ослабленного сердца, что приводит к дилатационной кардиомиопатии. При многих десминовых миопатиях в мышечных клетках обнаруживаются агрегаты этого белка, напоминающие кератиновые агрегаты, видимые в кератиноцитах больных с тяжелой формой EBS.
С мутациями в виментине не связано ни одно из известных заболеваний, что, по-видимому, отражает его способность экспрессироваться вместе с другими белками филаментов.
Почти все мутации в белках нейрофиламентов, относящихся к типу IV, локализованы в головном или хвостовом домене. Они связаны с такими нейродегенеративными заболеваниями, как амиотрофный латеральный склероз, Charcot-Marie-Tooth типа 1 и 2Е и болезнь Паркинсона. Большая длина цитоплазмы нейрона делает ее уязвимой к воздействиям различного рода, а не только к связанным с ее повышенной хрупкостью. Белки нейрофиламентов образуются в клетке и затем с помощью белковых моторов микротрубочек транспортируются по аксонам.
Сплетения нейрофибрилл служат общим показателем нейродегенеративного процесса. Они представляют собой внутриклеточные скопления нейрофиламентов, однако неизвестно, являются ли они причиной возникновения патологии. Эти сплетения могут возникать вследствие воздействий других факторов, влияющих на транспорт в аксонах, например такого, как нарушение функционирования микротрубочек По сравнению с кератинами, при развитии заболеваний, мутации в белках нейрофиламентов гораздо труднее поддаются причинно-следственному анализу.
«Кератинопатии» обычно проявляются при рождении или вскоре после него, в то время как большинство нейродегенеративных заболеваний развиваются поздно, что затрудняет проведение генетических исследований.
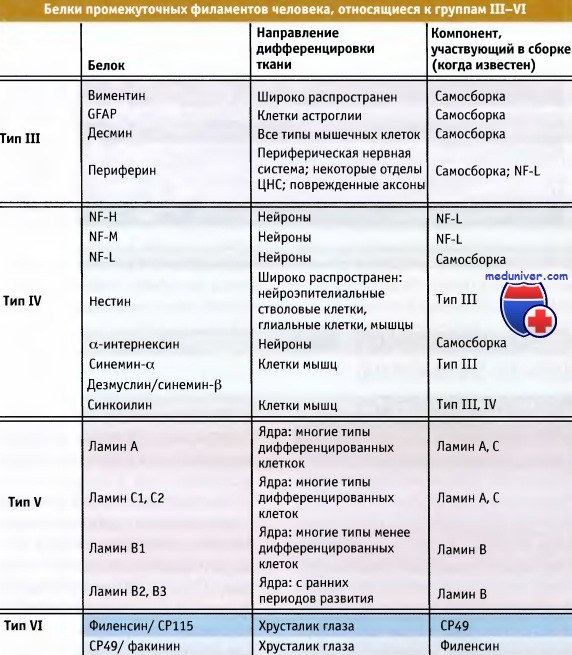
Белки промежуточных филаментов человека, объединенные в типы III—IV по принципу гомологии.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом