
Дана характеристика толстого филамента. Описан его состав и строение, а также расположение в структуре саркомера. Главной функцией толстого филамента является участие в процессе мышечного сокращения.
Состав и строение толстого филамента
После того, как мы разобрались из чего состоит саркомер, давайте рассмотрим состав и строение одного из его структурных элементов — толстого филамента. Эти знания необходимы для понимания механизма сокращения скелетных мышц.
Характеристика толстого филамента
Главная функция толстого филамента – участие в процессе сокращения саркомера. Толстый филамент занимает центральное место в структуре сота.
В структуре сота он окружен с двух сторон шестью тонкими филаментами. Расстояние между толстым и тонким филаментами составляет около 13 нм. Расстояние между толстыми филаментами в саркомере составляет приблизительно 32,5 нм. Тренировка не влияет на расстояние между толстыми филаментами.
Состав толстого филамента
Основу толстого филамента составляет белок миозин. Молекула миозина состоит из двух сплетенных протеиновых (белковых) нитей. Один конец этой молекулы называется хвостом. На другом конце эти нити заканчиваются головками, которые называются миозиновыми (рис. 1). Очень часто при описании процесса сокращения саркомера миозиновые головки называются поперечными мостиками.
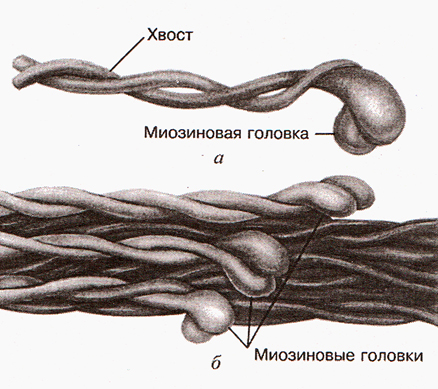
Рис. 1. Молекула миозина (а) и толстый филамент (б) (Дж.Х. Уилмор, Д.Л. Костилл, 1997)
В 1939 году советскими учеными В.А. Энгельгардом и М.Н. Любимовой было установлено, что молекула миозина обладает АТФ-азной активностью, то есть участвует в реакции гидролиза АТФ – основной реакции энергообеспечения мышечной деятельности энергией. Позже было установлено, что на каждой миозиновой головке имеются центры. В одном из них находится фермент АТФ-аза. Другой центр предназначен для связывания головки миозина с тонким филаментом.
В настоящее время на основе степени активности фермента АТФ-азы производится разделение мышечных волокон на типы.
Строение толстого филамента
Каждый толстый филамент представляет собой своеобразную структуру, похожую на ствол дерева. Его составляют приблизительно 360 молекул миозина, которые сплетены, как змеи, хвостами и имеют, соответственно, 720 миозиновых головок, расположенных в строгом порядке одна относительно другой (рис. 2).

Эта структура биполярна. В ее середине соединяются (конец в конец) два «ствола». Соединение «стволов» представляет собой М-диск. Головки толстого филамента («кроны» дерева) обращены в разные стороны. Установлено, что «кроны дерева» связаны с Z-диском, разделяющим саркомеры особым белком – титином (тайтином).
Самосборка толстого филамента
Установлено, что в растворах слабой ионной силы молекулы миозина начинают агрегировать (соединяться). В начале хвосты молекул миозина, объединяясь попарно, образуют короткую нить. Головы исходных молекул направлены в противоположные стороны. Затем этот процесс продолжается путем добавления новых молекул миозина. В результате этого формируется толстый филамент.
Белок толстых филаментов мышечных тканей
• Миозин II представляет собой мотор, обеспечивающий сокращение мышц
• Актин и миозин II являются основными компонентами саркомера, основной единицей сокращения поперечнополосатых мышц
Мышцы представляют собой ткань, способную к сокращению, которая обеспечивает движение тела и подвижность органов. В соответствии с внешним видом сократительных волокон, мышцы можно подразделить на две основных группы: поперечнополосатые и гладкие. Волокна поперечнополосатых мышц называются так потому, что при большом увеличении можно видеть их характерную исчерченность. К этой группе относятся скелетные и сердечная мышцы.
Скелетные мышцы обеспечивают подвижность скелета, а сердечная сокращение сердца. Волокна гладких мышц не исчерченные и имеют веретенообразную форму. Эти мышцы образуют стенки органов, например мочевого пузыря, кровеносных сосудов и желудочно-кишечного тракта.
Белковый мотор, который обеспечивает сокращение мышц, относится к семейству миозина II. Этот миозин принадлежит к числу наиболее распространенных белков позвоночных, он легко выделяется и поэтому является одним из хорошо изученных белков.
На рисунке ниже представлено строение молекулы миозина II, состоящей из шести полипептидных цепей: двух тяжелых и двух наборов, каждый из которых состоит из двух легких цепей.
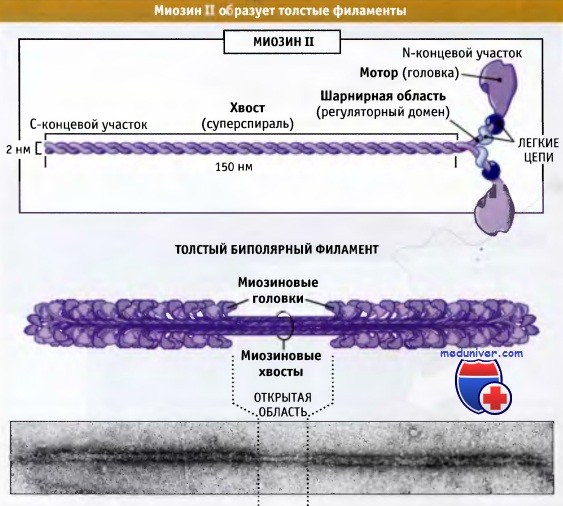
Миозин II представляет собой гексамерный комплекс,
состоящий из двух тяжелых цепей и двух пар различных легких цепей.
Эти комплексы собираются в биполярные толстые филаменты.
Дистальный участок хвоста мышечного миозина II связан с другими молекулами миозина II, образуя волокна, состоящие примерно из 300 молекул миозина. Эти волокна биполярны, и моторные домены всех миозинов направлены от центральной зоны. Эти волокна называются толстые биполярные филаменты. В настоящем разделе мы обсудим организацию и роль миозина II в сокращении поперечнополосатых мышц. Эти вопросы хорошо изучены.
Поперечнополосатые мышцы состоят из пучков мышечных волокон. Последние представляют собой крупные многоядерные клетки, длина которых варьирует от нескольких миллиметров до нескольких сантиметров, а диаметр составляет 20-100 мкм. Рисунок ниже показывает, что каждое мышечное волокно состоит более чем из 1000 миофибрилл, которые представляют собой палочковидные органеллы, способные к сокращению. Миофибриллы состоят из повторяющихся единиц, которые называются саркомеры. Саркомеры расположены друг за другом и обусловливают характерную исчерченность мышц.
Читайте также: Как правильно написать ткани или тканей
Саркомеры представляют собой основные сокращающиеся единицы поперечнополосатых мышц; при сокращении и расслаблении мышцы их длина изменяется. Как показано на рисунке ниже, саркомеры содержат толстые филаменты, состоящие, главным образом, из биполярных филаментов миозина II, и тонких филаментов, которые содержат актиновые нити и регуляторные белки. Оперенные концы актиновых филаментов с одной стороны прикреплены к структуре саркомера, которая носит название Z-диск. При этом все актиновые филаменты с одной стороны Z-диска имеют одинаковую полярность.
Актиновые филаменты прикрепляются к Z-диску и копируются за счет связывания с копирующим белком (CapZ), что предотвращает деполимеризацию актиновых филаментов. Заостренные концы актиновых филаментов ориентированы к центру саркомера и копированы тропомодулином. С актиновыми филаментами также взаимодействует белок небулин; он может регулировать сборку волокон и длину тонких филаментов.
Толстые филаменты находятся в центре, на М-линии, между Z-дисками. М-линия представляет собой структуру, состоящую из гибко связанных биполярных толстых филаментов; эти связи скрепляют толстые филаменты, гексагонально расположенные внутри. Наряду с М-линией, белок филаментов, который называется титаном, образует эластичные связи между Z-дисками и миозиновыми волокнами. Титин обеспечивает центровку толстых филаментов в саркомере и действует как пружина, которая не допускает растягиваться саркомеру.
Тонкие и толстые филаменты переплетаются, образуя трехмерную решетчатую структуру. Поскольку саркомер биполярен, в обеих половинах миозиновые моторы по отношению к актину ориентированы одинаково. При сокращении моторные домены миозина толстых филаментов взаимодействуют с актином тонких филаментов. При сокращении саркомер укорачивается за счет скольжения тонких и толстых филаментов относительно друг друга, что сближает соседние Z-диски к центру саркомера. По мере продвижения головок миозина к зазубренным концам актиновых филаментов, длина толстых и тонких филаментов остается постянной.
Ширина саркомера в расслабленной мышце позвоночных составляет около 3 мкм, а при сокращении она становится около 2,4 мкм.
В мышечном волокне укорачиваются группы, состоящие из тысяч саркомеров, что приводит к укорачиванию всей мышцы. Общая длина укороченного мышечного волокна определяется двумя факторами: длиной, на которую укорачивается каждый саркомер, и количеством саркомеров в группе. Процентная величина, на которую происходит укорачивание мышечных волокон, одна и та же, независимо от их длины.
Величина усилия, генерируемого саркомером, пропорционально числу акто-миозиновых взаимодействий в половине саркомера, а величина усилия, которое развивает мышчное волокно, пропорционально числу параллельно расположенных саркомеров. Таким образом, штангисты увеличивают свою силу за счет увеличения площади поперечного сечения мышц, а не за счет их длины.
Как показано на рисунке ниже, сокращение поперечнополосатых мышц регулируется с участием тропонин-тропомиозинового комплекса, который связан с актином в тонких филаментах. Молекулы тропомиозина представляют собой суперспирализованные полипептиды длиной 40 нм. Она располагаются друг за другом вдоль актиновых спиралей. Тропонин представляете собой комплекс из трех различных белков: тропонина С, тропонина I и тропонина Т. Один комплекс связывается с тропомиозином так, что они располагаются вдоль тонких филаментов через 40 нм интервалы.
При низких концентрациях ионов кальция тропомиозин находится в таком состоянии, что пространственно блокирует сайт связывания миозина на актине, так что мышца расслабляется и гидролиз АТФ под действием миозина происходит очень медленно. За счет взаимодействия актина с миозином релаксированные саркомеры способны к пассивному растяжению, оказывая небольшое сопротивление.
Нервные импульсы вызывают выход кальция в цитозоль из саркоплазматического ретикулума. Последний представляет собой органеллу, которая депонирует кальций в мышце. Повышение уровня кальция в цитозоле приводит к его связыванию с тропонином-С и к конформационным изменениям в молекуле белка. В результате этих изменений тропомиозин отходит от миозин-связывающего сайта в актине, и миозин получает возможность взаимодействовать с актином и генернировать усилия за счет механохимического цикла.
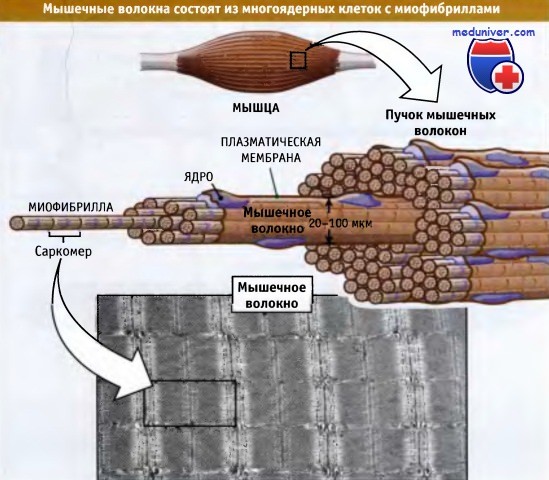 Скелетные мышцы состоят из мышечных волокон, содержащих миофибриллы и представляющих собой удлиненные клетки.
Скелетные мышцы состоят из мышечных волокон, содержащих миофибриллы и представляющих собой удлиненные клетки.
Миофибриллы способны к сокращению и содержат повторяющиеся структурные единицы, называемые саркомерами. 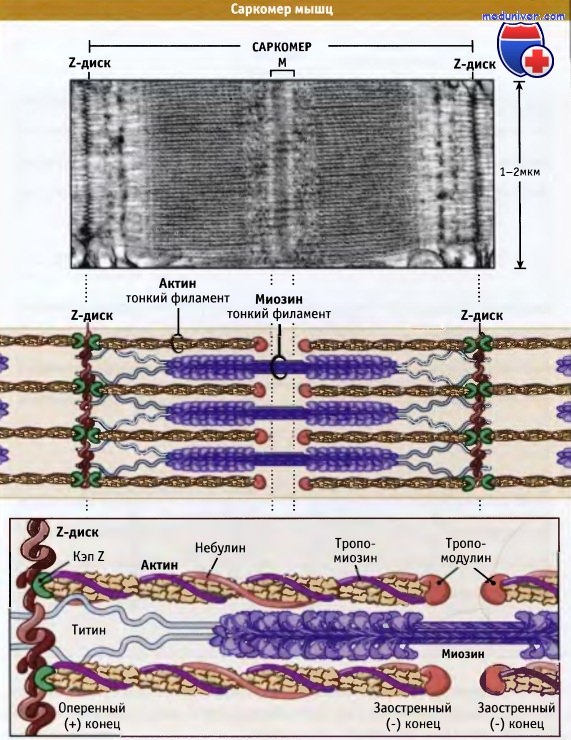 На конце каждого саркомера находится Z-диск, к которому через CapZ (кэпирующий белок) присоединяются актиновые филаменты.
На конце каждого саркомера находится Z-диск, к которому через CapZ (кэпирующий белок) присоединяются актиновые филаменты.
Толстые миозиновые филаменты соединяются с Z-диском через белок титин и вплетаются между актиновыми филментами.
Небулин проходит от Z-диска к тропомодулину. Однако как он связывает актин, пока неизвестно. 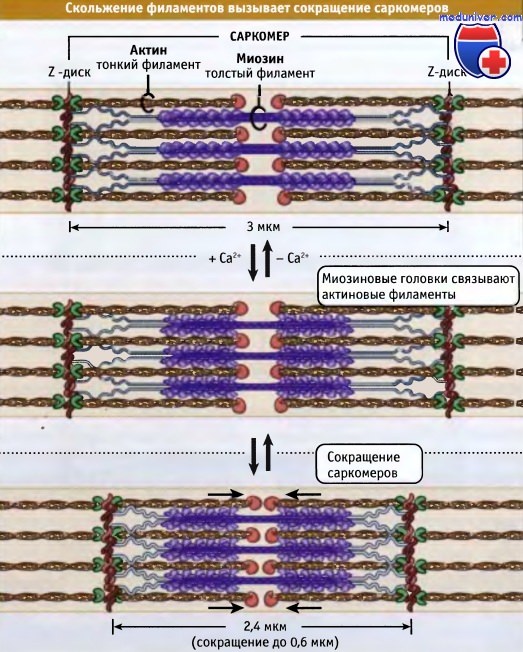 Сокращение мышцы происходит, когда толстые миозиновые филаменты присоединяются к актиновым и перемещают их таким образом,
Сокращение мышцы происходит, когда толстые миозиновые филаменты присоединяются к актиновым и перемещают их таким образом,
что Z-диски сдвигаются относительно друг друга. При этом длина саркомера уменьшается. 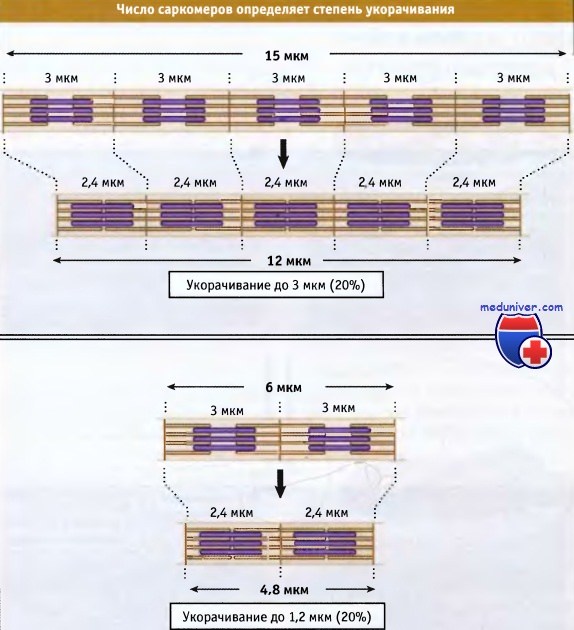 Чем длиннее миофибрилла (т. е. чем больше саркомеров), тем более она укорачивается при сокращении.
Чем длиннее миофибрилла (т. е. чем больше саркомеров), тем более она укорачивается при сокращении.
Однако в процентном отношении величина укорачивания не зависит от числа саркомеров. 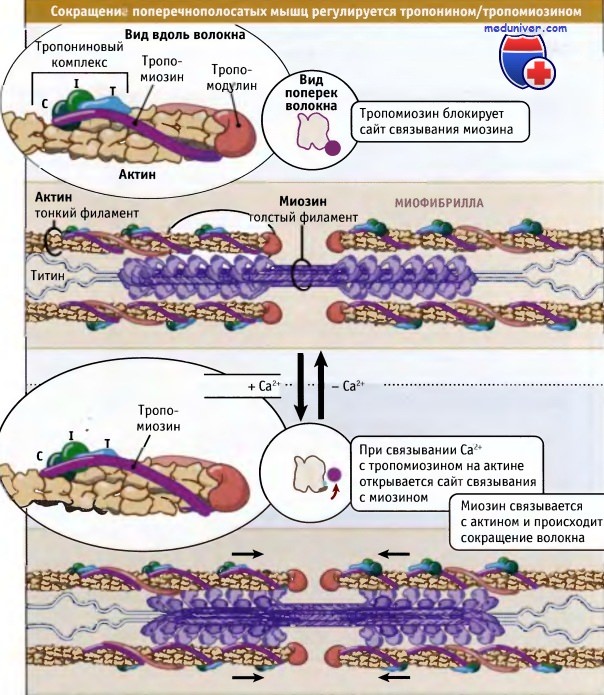 Сокращение поперечнополосатых мышц регулируется уровнем Са2+ в цитозоле,
Сокращение поперечнополосатых мышц регулируется уровнем Са2+ в цитозоле,
который определяет положение комплекса тропонин/тропомиозин относительно миозина и актина.
Читайте также: Виды основной ткани паренхимы растений
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Белок толстых филаментов мышечных тканей
• Некоторые белки промежуточных филаментов, относящиеся к типу III и IV, ко-экспрессируются в разных тканях
• Многие белки типа III и IV образуют друг с другом сополимеры
• Совместная экспрессия множественных типов белков промежуточных филаментов может маскировать эффект мутаций в белке, принадлежащим к одному из типов
• Десмин представляет собой необходимый белок мышц
• Виментин часто экспрессируется в одиночных клетках
• Мутации в генах белков типа III или IV обычно связаны с мышечными или неврологическими расстройствами, носящими дегенеративный характер
В таблице ниже перечислены гомологичные некератиновые белки промежуточных филаментов, которые относятся к группам III—IV. В данной статье мы рассмотрим белки промежуточных филаментов, относящихся к типам III и IV. Эти типы различаются по принципу гомологии по определенным последовательностям. К белкам III типа относятся десмин, виментин, фибриллярный кислый белок глиального происхождения (GFAP, фибриллы которого представлены на рисунке ниже) и периферии.
Эти белки дифференциально экспрессируются в клетках соединительной ткани, мышц, в нейронах и некоторых других дифференцированных клетках. Их экспрессия индивидуально регулируется для каждого белка и тесно связана со специфическими процессами дифференцировки. Некоторые из этих белков одинаково экспрессируются в разных тканях, а другие могут образовывать друг с другом сополимеры. Однако они никогда не образуют таких комплексов с кератинами (тип I/II) или с ламинами (тип V).
Так же как и для других белков промежуточных филаментов, мы пока точно не знаем, каким образом вариации в аминокислотной последовательности этих белков связаны с их специфическими функциями в клетках различных типов, хотя мутации в них хорошо коррелируют с различными нарушениями в функционировании соответствующей ткани.
Десмин представляет собой белок, необходимый для функционирования клеток мышц всех типов (поперечнополосатых, сердечной и гладких); подобно кератину эпителиальных клеток, десмин обеспечивает упругость ткани при физических воздействиях. Десминовые филаменты проходят между точками скрепления сократительных структур (например, саркомер поперечнополосатых мышц). Эти места в клетках мышц являются сайтами приложения значительных механических усилий. Поэтому десмин можно рассматривать как белок, обеспечивающий устойчивость мышечных клеток к механическому воздействию.
Такая точка зрения возникла после того, как оказалось, что ткани, в клетках которых десмин отсутствует или находится в мутантной форме, теряют способность противостоять механическим воздействиям. Десмин образует комплексы с другими белками типа III; в некоторых клетках он сополимеризуется также с белками типа IV.
В отличие от десмина, который экспрессируется в тканях состоящих из плотных агрегатов клеток, таких как мышечная и эпителиальная, экспрессия виментина типична для клеток, которые функционируют изолированно, или в виде рыхло-связанных клеточных ассоциатов или слоев. В процессе развития виментин экспрессируется после кератинов и остается во многих мезенхимальных и соединительнотканных клетках взрослого организма, от фибробластов до кроветворных клеток и клеток эндотелия сосудов, а также некоторых видов эпителия.
Астроциты и глиальные клетки представляют собой клетки центральной нервной системы, отличные от нейронов. Они необходимы для роста, дифференцировки и регенерации последних. Все астроглиальные клетки экспрессируют GFAP, который обычно экспрессируется вместе с виментином или белком нестином, относящимся к группе IV. Такая совместная экспрессия защищает клетки от последствий мутаций или утраты одного из генов. Эксперименты по двойному нокауту генов, проведенные на животных, показали, что эти промежуточные филаменты типа III необходимы для нормального функционирования астроцитов в центральной нервной системе.
Эти клетки необходимы для заживления ран и развития устойчивости к осмотическому шоку. Для нормального функционирования астроцитам необходимо сформировать отростки, которые при отсутствии промежуточных филаментов несут различные дефекты.
Периферии, главным образом, экспрессируется в клетках периферической нервной системы. При формировании аксонами отростков они вначале синтезируют периферии и виментин; затем этот синтез сменяется экспрессией триплета белков нейрофиламентов (NF-L, NF-M, и NF-H). Впрочем, после повреждения нервной ткани экспрессия периферина быстро возобновляется. По-видимому, для нормального выполнения белком своих функций важен его уровень. Так, в экспериментах на животных показано, что слишком высокое содержание периферина приводит к летальным последствиям, а его отсутствие — к утрате некоторых небольших сенсорных аксонов.
Филаменты астроцитов спинного мозга, состоящие из глиального фибриллярного кислого белка (GFAP).
Читайте также: Механические ткани строение функции расположение
В настоящее время гомологичный тип IV включает триплет белков нейрофиламентов низкой, средней и высокой молекулярной массы (NF-L, NF-M и NF-H), а также а-интернексин, нестин, синкоилин и синемин. В тканях большинство белков типа IV присутствуют в гетерополимерной форме, и гораздо более эффективно образуют сополимеры с белками типа III или другими белками, относящимися к типу IV. В зрелых нейронах, белки триплета нейрофиламентов почти всегда экспрессируются вместе, в то время как синкоилин и два продукта альтернативного сплайсинга синемина (а и b) преимущественно экспрессируются в клетках мышц и сополимеризуются с белками группы III.
Согласно иммуногистохимическим данным, синкоилин и синемин, так же как и десмин, локализованы в мышечных клетках, в точках приложения нагрузки. Возможно, что эти белки промежуточных филаментов тоже обеспечивают устойчивость клеток к нагрузкам.
Необычайная длина аксональных выростов в нейронах (у нейронов седалищного нерва человека она может достигать 1 м) свидетельствует о важности поддержания соответствующего уровня экспрессии белков нейрофиламентов, необходимых для укрепления цитоплазмы. В процессе развития экспрессия нейрофиламентов происходит по сложной программе, которая включает одновременный синтез многих белков. Соответствующие профили накладываются и смещаются относительно друг друга. Таким образом, нейроны постоянно экспрессируют промежуточные филаменты. Первым экспрессируются нестин и виметин. (Подобно GFAP астроглии, при раневых повреждениях нейрональный нестин может экспрессироваться повторно.)
По мере удлинения выростов, доминирующим становится синтез а-интернексина, который сменяется NF-L, и наконец, происходит экспрессия крупных белков нейрофиламентов. В стабилизации сформировавшихся аксонов и нейритов важную роль играет триплет белков нейрофиламентов.
У нескольких белков, относящихся к группе IV, имеются длинные хвостовые домены, которые участвуют в организации каркасных структур цитоплазмы. В хвостовом домене NF-H присутствует ряд повторов, представляющих собой последовательность Lys-Ser-Pro, которая является сайтом фосфорилирования. Когда происходит фосфорилирование этих повторов, хвостовой домен приобретает высокий положительный заряд и ориентируется под прямым углом к оси филамента. Это способствует растяжению цитоплазмы аксона. У мышей число аксонов и их диаметр коррелирует с уровнем экспрессии белков нейрофиламентов.
Большему диаметру аксона соответствует более быстрая проводимость нервного импульса, что является особенно важным фактором в эволюции крупных по размеру тел позвоночных.
Мутации в генах белков группы III связаны с развитием многочисленных патологических состояний, механизм которых в некоторых случаях удалось выяснить в экспериментах на животных. У человека мутации в генах GFAP связаны со смертельным нейродегенеративным заболеванием, болезнью Александера. В экспериментах на животных показано, что в отсутствие GFAP наблюдается аномальная реакция астроглии на повреждения, и астроциты не образуют цитоплазматических выростов. В десмине обнаружено много патологических мутаций, которые происходят в разных участках белковой структуры.
У человека мутации в гене десмина связаны с дефектами сердечно-сосудистой системы, в особенности с развитием сердечной недостаточности на почве дилатационной кардиомиопатии, а также с некоторыми формами мышечной дистрофии. У мышей при отсутствии десмина стенки важнейших кровеносных сосудов становятся слишком мягкими и не могут поддерживать надлежащий уровень кровяного давления. Это вызывает растяжение уже ослабленного сердца, что приводит к дилатационной кардиомиопатии. При многих десминовых миопатиях в мышечных клетках обнаруживаются агрегаты этого белка, напоминающие кератиновые агрегаты, видимые в кератиноцитах больных с тяжелой формой EBS.
С мутациями в виментине не связано ни одно из известных заболеваний, что, по-видимому, отражает его способность экспрессироваться вместе с другими белками филаментов.
Почти все мутации в белках нейрофиламентов, относящихся к типу IV, локализованы в головном или хвостовом домене. Они связаны с такими нейродегенеративными заболеваниями, как амиотрофный латеральный склероз, Charcot-Marie-Tooth типа 1 и 2Е и болезнь Паркинсона. Большая длина цитоплазмы нейрона делает ее уязвимой к воздействиям различного рода, а не только к связанным с ее повышенной хрупкостью. Белки нейрофиламентов образуются в клетке и затем с помощью белковых моторов микротрубочек транспортируются по аксонам.
Сплетения нейрофибрилл служат общим показателем нейродегенеративного процесса. Они представляют собой внутриклеточные скопления нейрофиламентов, однако неизвестно, являются ли они причиной возникновения патологии. Эти сплетения могут возникать вследствие воздействий других факторов, влияющих на транспорт в аксонах, например такого, как нарушение функционирования микротрубочек По сравнению с кератинами, при развитии заболеваний, мутации в белках нейрофиламентов гораздо труднее поддаются причинно-следственному анализу.
«Кератинопатии» обычно проявляются при рождении или вскоре после него, в то время как большинство нейродегенеративных заболеваний развиваются поздно, что затрудняет проведение генетических исследований.
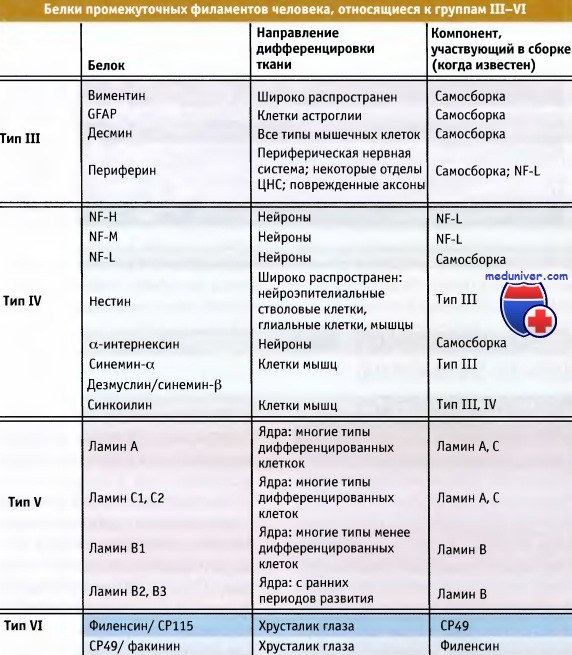
Белки промежуточных филаментов человека, объединенные в типы III—IV по принципу гомологии.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом