

Коллагены составляют основу структуры кожи, костей, хрящей, сухожилий, кровеносных сосудов и играют важную роль в сохранении нормальной структуры и функции соединительной ткани всех животных (Слуцкий, 1969, 1985; Риггз, Мелтон, 2000; Руденская, 2003).
Коллаген (К) – нерастворимый фибриллярный белок, первичная структура которого складывается из повторяющихся последовательностей триплетов аминокислот глицин-X-Y, где X и Y позиции чаще заняты, соответственно, пролином и гидроксипролином. Эти повторяющиеся последовательности позволяют трем коллагеновым полипептидам (называемым α-цепями) формировать полужесткие, очень стабильные трехспиральные молекулы (рис. 3). Они могут быть гомополимерными (три идентичные α-цепи) и гетерополимерными (две или три разные α-цепи). Под влиянием регулярно располагающихся остатков пролина и оксипролина цепь принимает форму ломаной спирали; это обусловливается жесткостью боковых групп пролина, а также тем обстоятельством, что пептидные связи, в образовании которых участвуют пролин и оксипролин, не могут образовать водородных связей. Остатки глицина образуют межцепочечные водородные связи, способствующие сохранению прочности структуры коллагена.



Рис. 3. Структура коллагена: а – электронная микрофотография коллагена соединительной ткани. Расстояние между повторяющимися структурными единицами равно 700 Å; б – конформация полипептидных цепей в трёхцепочечной молекуле тропоколлагена; в – ступенчатое расположение молекул тропоколлагена обусловливает появление в гидратированных волокнах коллагена повторяющихся структурных единиц, расстояние между которыми равно 700 Å (по Мусил и др., 1984)
Аминокислотные цепи коллагена обернуты друг вокруг друга и образуют «трехволоконный канат», где отдельные волокна связаны между собой водородными связями. Такая пространственная структура возможна, только если аминокислотная последовательность строго соблюдается.
В результате образуется трехволоконная фибриллярная молекула – тропоколлаген, обладающая большой прочностью на растяжение. Это название происходит от слова тропос – обращенный внутрь – из-за того, что коллагеновые волокна всех соединительных тканей, выстланы тропоколлагеновыми молекулами, соединенными «конец в конец» и «бок о бок» – в шахматном порядке. Гидроксильные группы некоторых остатков лизина и оксилизина участвуют в образовании связи между соседними молекулами тропоколлагена. Таким образом, формируется жесткое нерастяжимое волокно. Фибробласты синтезируют молекулы тропоколлагена и выбрасывают их в матрикс, и только здесь происходит сборка коллагеновых волокон (рис. 4).
Коллагены кожи содержат в больших концентрациях пролин и оксипролин (около 20 % от всех остальных аминокислот), глицин и аланин (свыше 50 % от содержания других аминокислотных остатков), ароматические и серосодержащие аминокислоты практически отсутствуют или содержатся в весьма малых количествах. Кроме того, коллаген является одним из немногих белков, содержащих оксипролин и оксилизин. Оксипролин и оксилизин образуются в молекуле проколлагена не в результате биосинтеза, а при гидроксилировании пролина и лизина, которое начинается в период трансляции коллагеновой мРНК на рибосомах (Неклюдов, 2003).

Рис. 4. Строение фибробласта и основные этапы синтеза коллагена. Г – аппарат Гольджи; Я – ядро; ШЭР – шероховатый эндоплазматический ретикулум; СП –секреторный пузырек; ТК – тропоколлаген; КВ – коллагеновое волокно
В настоящее время идентифицировано более 25 различных α-цепей, из которых формируется до 14 разных типов коллагеновых молекул, некоторые из них являются тканеспецифичными (Ленинджер, 1974; Риггз, Мелтон, 2000).
Таким образом, коллагены – это белки, которые:
a) содержат несколько повторов аминокислотной последовательности -Gly-X-Y-, в X-положении которой чаще всего расположен пролин, а в Y-положении – 4-гидрокси-пролин;
б) могут состоять из трех цепей с повторяющимися последовательностями, обладающими способностью к сворачиванию в характерную тройную спираль. По меньшей мере, 19 белков определены в настоящее время как принадлежащие к коллагенам. 10 родственных им белков содержат коллагеноподобные домены.
Коллагеновые белки составляют около половины массы сухого вещества суставного хряща; вблизи суставной поверхности их концентрация приближается к 90 %. В других видах хрящевой ткани коллагены количественно преобладают над другими белками, обеспечивая прочность на растяжение и разрыв. (Слуцкий, 1985). Количественно преобладающим белком протеогликанового комплекса хрящевой ткани является коллаген второго типа (КII). Он в незначительных количествах обнаружен в других специализированных тканях, например, в стекловидном теле глаз некоторых видов животных. Хрящевой ткани присущ необычный полиморфизм коллагеновых компонентов, проявляющийся присутствием большого числа минорных коллагенов. Молекулярная формула КII хрящевой ткани имеет следующий вид: [α1(ΙΙ)]3, что означает наличие трех идентичных α1(II)-цепей, которые отличаются от α1(I)-цепей более высоким содержанием оксилизина. Столь высокая концентрация оксилизина способствует увеличению количества связанных с ним углеводов. По-видимому, такая структура обеспечивает большую сольватную оболочку коллагена типа II по сравнению с типами I или III. В процессе биосинтеза коллагена С-пропептиды соединяются между собой бисульфидными мостиками и после отщепления от молекулы образуют белок, который называется хондрокальцин, с м.м. около 100 000 Да (Слуцкий, 1985).
Читайте также: Как получают хлопчатобумажные ткани
Коллаген одиннадцатого типа (КXI) составляет приблизительно 3 % хрящевого коллагена и образован двумя разными типами цепей (формула молекулы α1(XI)2 α2(XI)). Со старением организма его количество в хряще уменьшается в пользу коллагена пятого типа (КV) (Канунго, 1982; Риггз и Мелтон, 2000).
Для хрящевой ткани характерно наличие наибольшего количества минорных коллагеновых компонентов. Такой полиморфизм коллагенов позволяет считать минорные компоненты регуляторами адаптационной пластичности хряща, метаболизма хондроцитов и морфогенетических процессов (Слуцкий, 1985).
Содержание коллагена в различных тканях и органах сельскохозяйственных животных (КРС, свиньи) характеризуют следующие данные (% от массы сухого вещества): дерма – 80–90; ахиллово сухожилие – 86; костная ткань – 17,5–25; хрящи – 46–67; роговица и склера глаз – 70; мышцы – 10. При этом коллаген составляет от 25 до 33 % от общего количества белка (Титов, Апраксина, 1995). У костистых рыб коллагеновые белки преобладают в коже, сухожилиях, плавательном пузыре; у кольчатых червей и иглокожих в кожно-мускульном мешке; у моллюсков в кожных покровах и опорных хрящевых элементах.
Наиболее изучены свойства коллагенов в коже и мышечной ткани рыб. Коллаген кожи костистых рыб характеризуется частой встречаемостью молекул, состоящих из трех генетически различающихся α-цепочек, гетеротримера α1 α2 α3; среди изученных позвоночных животных цепочка α3 отмечена лишь у костистых рыб. В целом коллагены мышечной ткани костистых рыб, полученные методом электрофореза, идентичны соответствующим коллагенам типа I из кожи и имеют сходный аминокислотный состав. Однако коллагены мышечной ткани рыб более устойчивы к тепловой денатурации (более 100 °С), чем коллагены кожи, что объясняется более высокой степенью гидроксилирования пролина в мышечном коллагене. Термальная устойчивость мышечного коллагена определяется видом рыбы и по мере ее возрастания рыб можно расположить в следующем порядке: карп, угорь, скумбрия, сайра, кета.
В табл. 1 приведены данные о содержании коллагена и гексозаминов в соединительных тканях различных видов животных.
Составы субъячеек коллагена I типа из мышечной ткани и кожи являются идентичными у угря, скумбрии, сайры и карпа и отличаются у кеты. У последней коллаген кожи состоит из α1 α2 α3 гетеротримеров, а основная часть мышечного коллагена – из (α1)2α2 гетеротримеров (Богданов, Сафронова, 1993).
Содержание гексозаминов и коллагена в соединительной ткани животных
Белок в соединительных тканях человека

На долю соединительной ткани приходится более 50 % массы тела животного. Существенной особенностью, отличающей ее от других и присущей в большей или меньшей степени всем ее разновидностям, является количественное преобладание межклеточного вещества над клеточными элементами. Межклеточный матрикс состоит из волокнистых компонентов, а пространство между ними заполнено основным веществом, содержащим гликопротеины. Волокнистые компоненты состоят из коллагеновых и эластиновых волокон.
Белковый состав соединительной ткани представлен в основном коллагеном и эластином, на их долю приходится практически 90 % всех белков. Это фибриллярные белки с характерной удлиненной формой молекулы, обусловленной параллельным расположением полипептидных цепей. Аминокислотный состав коллагена и эластина отличается от мышечных. В коллагене на долю остатков глицина и аланина приходится около 45 %, также содержатся такие аминокислоты: гидроксилизин и гидроксипролин. На долю пролина и гидроксипролина приходится около 21 %. В эластине суммарное содержание глицина, аланина, валина и пролина составляет почти 70 %. Изоэлектрическая точка смещена в слабокислую область [4]. Еще одной характерной чертой является регулярность в последовательности аминокислот, чего нельзя сказать о белках мышечной ткани. Для данных белков характерно присутствие повторяющихся участков аминокислот. В коллагене — в виде триплетов четырех видов (Х – иминокислота, Y – аминокислота):
Читайте также: Расчетов ткани для ламбрекена
— Gly — Х — Х -; — Gly — Х — Y -; — Gly — Y — Х -; — Gly — Y — Y -.
В эластине — тетрамер: — Gly — Gly- Val — Pro -; пентамер: — Pro — Gly — Val — Gly — Val -; гесамер: — Pro — Gly — Val — Gly — Val — Ala.
Для соединительнотканных белков характерно присутствие внутри- и межмолекулярных поперечных сшивок: альдольная, гистидин-альдольная, лейцин-норлейциновая, основание шиффа. В результате появляется возможность ковалентного связывания четырех боковых цепей друг с другом, причем они могут принадлежать разным молекулам белка.
В эластине значительно меньше содержание лизина это обусловлено тем, что боковые цепи четырех остатков аминокислоты ферментативным путем превращаются в десмозин, в результате формируется поперечная сшивка между соседними полипептидными цепями в молекуле эластина. Именно присутствие десмозина отличает эластин от всех других белков, включая и коллаген.
Благодаря своей уникальной структуре белки соединительной ткани в нативном состоянии не подвергаются действию протеолитических ферментов. Ткани, содержащие коллаген и эластин, отличаются особой прочностью, способностью выдерживать значительные нагрузки.
При всем сходстве данных белков, каждый из них является индивидуальным белком, со свойствами присущими только ему одному.
Молекула коллагена имеет относительную молекулярную массу 300000, длину 280 нм и толщину 1,4 нм. Каждая a-цепь содержит в среднем около 1000 аминокислотных остатков. Еще одной особенностью коллагена является его гетерогенность. В настоящее время описано 28 типов коллагена, которые кодируются более чем 40 генами. Они отличаются друг от друга по аминокислотной последовательности, а также по степени модификации – интенсивности гидроксилирования или гликозилирования. Общим для всех коллагенов является существование одного или более доменов, содержащих тройную спираль и присутствие их во внеклеточном матриксе [5].
Эластин изучен меньше, чем коллаген. Наличие гибкой случайной конформации молекул эластина и большое количество поперечных сшивок обеспечивает исключительное свойство эластической ткани — обратимую деформацию под влиянием механического воздействия.
Практическое применение имеет в основном коллагенсодержащее сырье, это связано со способностью коллагена образовывать гели при взаимодействии с водой. Коллаген находит широкое применение в медицине для пластики сосудов, дефектов кожи, кости, мозговой оболочки, тазового дна, трахеи, печени и селезенки. Применяется в виде гемостатических средств и тампонов, мембран для диализа, шовного материала, а также для доставки лекарственных средств в организм человека, в пищевой промышленности – в качестве гелеобразователя.
Цель работы исследование возможности использования соединительнотканных белков в качестве носителей биологически активных веществ.
Материалы и методы исследований
Объектами исследования были сухожилия и выйная связка крупного рогатого скота, желатин, растворы гидролизатов коллагена и эластина, соли йода и железа, а также растительные настои, содержащие фенольные соединения, в частности антоцианы.
Основные этапы получения гидролизата коллагена: измельчение сухожилий, последовательная солевая и кислотная экстракция, тепловая обработка и охлаждение. Раствор гидролизата коллагена имел светло-матовый цвет, без видимых границ раздела фаз.
Читайте также: Нервная ткань физиология методичка
Эластин выделяли из выйной связки при последовательной экстракции: солевой, щелочной, водной. Растворимую форму белка получали методом биотрансформации.
Гидролизат представляет собой сложную смесь пептидов, жидкость светло-желтого цвета, без вкуса и запаха. Содержание белка в 1 мл гидролизата коллагена и эластина составило 30 мг и 40 мг соответственно.
При проведении экспериментов использовали аналитические методы качественного и количественного анализа.
Результаты исследований и их обсуждение
В результате проведенных исследований получены растворимые формы соединительнотканных белков – гидролизаты коллагена и эластина, которые исследовали в качестве матрицы микроэлементов, в частности железа и йода.
Источниками микроэлементов служили сульфат железа (II) и йодид калия. Одним из основных условий при проведении реакции являлся показатель рН среды. Выбор рН среды связан со свойствами микроэлементов и фермента. Так, в кислой среде йод восстанавливается до молекулярного состояния и улетучивается, в щелочной среде рН > 8,0 образуется гипойодид. Фермент при рН выше 6,0 инактивируется. Учитывая данные факторы, в растворах устанавливали рН 7,0 – 7,2. Расчет вводимого количества йодида калия производили по содержанию йода (76,5%).
При взаимодействии железа и гидролизатов коллагена и эластина были исследованы значения рН (9,0; 5,5; 3,5)±0,2. При рН 9,0±0,2 наблюдалось изменение окраски от светло-желтого (гидролизат эластина) и светло-матового (гидролизат коллагена) до темно-коричневого цвета с выпадением осадка в виде хлопьев, что снижало органолептические свойства и указывало на окисление железа.
При рН среды 3,5 и 5,5 наблюдалось одинаковое связывание железа без визуальных изменений растворов. Однако, учитывая значения рН максимальной устойчивости комплексов и большинства пищевых систем, выбрано значение 5,5±0,2.
Оптимальная температура процессов связывания йода и железа соединительнотканными белками выбрана 0-4 °С, так как при положительных температурах возможно микробное обсеменение исследуемых растворов.
Известно, что химические процессы протекают во времени. Для установления максимальной степени связывания в растворы гидролизатов эластина и коллагена вносили разные концентрации солей железа и йода и контролировали содержание микроэлементов через каждые 2 часа. Концентрации микроэлементов были подобраны исходя из суточной потребности организма человека в них [3]. Экспериментально установлено, что максимальное взаимодействие железа с гидролизатами наблюдалось по истечении 24 часов, в случае йода – 20 часов.
По результатам исследований учитывая совокупность данных по ИК — спектроскопии (о наличии полиглициновых структур и отсутствие связи C-I), содержанию глицина в гидролизатах, диссоциацию йодида калия в растворах на ионы, избирательное расщепление пептидной цепи пепсином (Х- глицин) можно предположить о возможности возникновения ионной связи между протонированными группами NH3+ гидролизатов и I-. На основании приведенных доводов, предложена гипотетическая модель связывания йода гидролизатами соединительнотканных белков:
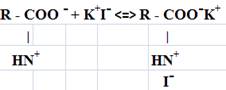
Для выяснения взаимодействия катиона железа с пептидами эластина и коллагена проведены исследования с использованием методов ИК и ЯМР-спектроскопии.
При ИК-спектроскопии в области 1400-1390 см-1 у образцов гидролизата эластина присутствует пик, который отвечает за группировку COO- . Данная область не содержит аналогичный пик в спектре в образцах эластин-железо. Следовательно, можно предположить, что свободная карбоксильная группа, присутствующая в пептидах гидролизата эластина, присоединила к себе железо с образованием комплекса. В диапазоне спектра 1130-1080 см-1 в образцах «эластин-железо» и «коллаген — железо» наблюдалось появление пиков, отвечающих за сульфатную группировку и свидетельствующих о возникновении ионной связи с SO42-.
Для более полной информации о взаимодействии между соедительнотканными белками и железом исследовали ядерно-магнитно-резонансные (ЯМР) спектры опытных образцов.
Анализ спектров показал, что в гидролизатах содержится большее количество СОО- групп (области 160-180 м.д., наличие большего количества пиков), в отличие от той же области в гидролизатах с железом. В области 50-65 м.д., которая отвечает за >СН-N
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом