
ТКАНЕВОЙ КАМБИЙ (лат. cambium смена, обмен) — малодифференцированные клетки, входящие в состав тканей и обладающие способностью к размножению и дифференцировке (специализации).
Термины «камбий», «камбиальные клетки» введены А. А. Заварзиным в 30-х гг. 20 в. С наличием Т. к. связывали физиол. регенерацию тканей. Но, как выяснилось позднее, регенерация (см.) может происходить и без размножения клеток. Наличие камбия не определяет способности к регенерации, а указывает лишь на механизм ее реализации в конкретной ткани. Камбиальные элементы, по А. А. Заварзину, являются не простыми остатками зародышевых листков, а их малодифференцированнымп потомками. Они располагаются либо в виде одиночных клеток среди дифференцированных клеток, напр, в поперечнополосатой мышечной ткани, либо группами — в многослойном плоском эпителии. В нек-рых случаях камбиальные клетки топографически отделены от дифференцированных форм (напр., бластные клетки красного костного мозга от клеток крови).
В настоящее время термины «камбий», «камбиальные клетки» употребляются редко, уступая место более точным определениям — «стволовые клетки» (полииотентные), «полустволовые клетки» (частично детерминированные, или коммутированные). В начале 20 в. А. А. Максимов высказал предположение о существовании в организме стволовой кроветворной клетки. В 00-х гт. 20 в. эта гипотеза подкрепилась экспериментальными данными Тил-ла и Мак-Каллока (J. Е. Till, E. A. McCulloch) о наличии в крови и кроветворных органах полипотентной клетки, по морфол. признакам похожей на лимфоцит. Идентифицированы стволовые клетки и для ряда других тканей. Все они характеризуются наличием диффузного хроматина в ядре, содержанием одного или большего количества ядрышек, слабым развитием органелл цитоплазмы, за исключением свободных рибосом (см. Клетка). Форма стволовых клеток различных систем клеточных дифференцировок неодинакова. В противоположность остальным пролиферирующим клеткам при делении стволовых клеток образуются потомки, к-рые необязательно подвергаются дальнейшей дифференцировки гг поэтому могут сохранять свой пролиферативный потенциал. Существенную роль в дифференцировке потомков стволовых клеток играет микроокружение. Если стволовые клетки — это длительно, медленно самоподдерживающаяся популяция клеток, то коммутированные клетки, берущие начало от стволовых клеток, интенсивно, многократно размножаются и подвергаются медленной дифференцировке, в ходе к-рой на уровне синтеза информационной РНК осуществляется программа их детерминации (см. Гистогенез). Детерминированные клетки перестают реагировать на микроокружение, но не становятся отличными от того тина клеток, к к-рому они принадлежали прежде (напр., класс морфологически распознаваемых пролиферирующих клеток красного костного мозга). В основе этих процессов лежит репрессия определенных генов на уровне трансляции генетического кода молекулы РНК на синтезируемые клеткой тканево-специфические молекулы белка (см. Ген, Трансляция). Для нек-рых частично детерминированных клеток доказана невозможность обратного перехода в стволовые клетки. В отношении других клеток однозначных данных нет, что, возможно, обусловлено неодинаковой степенью жесткости детерминации (программирования) развивающихся клеток и тканей.
Развитие учения о стволовых клетках уточняет прежние представления о Т. к. Разработка методов идентификации стволовых клеток в разных тканях имеет не только теоретический интерес, но и служит решению практически важных проблем, таких как дифференцировка, регенерация и др.
Библиогр.: Данин и Е. С. Понятие о тканевой камбиальности в современной гистологии, Усп. совр. биол., т. 29, № 3, с. 379, 1950; он же, Самообновление тканей и теория камбиальности академика А. А. Заварзина, Арх. анат., гистол., и эмбриол., т. 34, № 1, с. 16, 1957; 3 а-в а р з и н А. А. Курс гистологии и микроскопической анатомии, JI.,1939; он же, Избранные труды, т. 2, с. 217, М.— Л., 1953; Р а й ц и н а С. С. Идентификация стволовых клеток в некоторых системах клеточных дифференцировок у млекопитающих, Усп. совр. биол., т. 90, в. 1(4), с. 123, 1980; Фриденштейн А. Я. и Л у р и я Е. А. Клеточные основы кроветворного микроокружения, М., 1980; Чертков И. Л. и Фриденштейн А. Я. Клеточные основы кроветворения, М., 1977.
Без камбиальная ткань гистология
Эпителиальные ткани — древнейшие гистологические структуры, которые в фило- и онтогенезе возникают первыми. Основное свойство эпителиев — пограничность. Эпителиальные ткани (от греч. epi — над и thele — кожица) располагаются на границах двух сред, отделяя организм или органы от окружающей среды.
Читайте также: Оформленная эластическая соединительная ткань выйной связки теленка
Эпителии, как правило, имеют вид клеточных пластов и образуют наружный покров тела, выстилку серозных оболочек, просветов органов, сообщающихся с внешней средой во взрослом состоянии или в эмбриогенезе. Через эпителии осуществляется обмен веществ между организмом и окружающей средой. Важной функцией эпителиальных тканей является защита подлежащих тканей организма от механических, физических, химических и других повреждающих воздействий. Некоторые эпителии специализированы на выработке специфических веществ — регуляторов деятельности других тканей организма.
Производными покровных эпителиев являются железистые эпителии. Особый вид эпителия — эпителий органов чувств.
Эпителии развиваются с 3-4-й недели эмбриогенеза человека из материала всех зародышевых листков. Некоторые эпителии, например эпидермис, формируются как полидифферонные ткани, так как в их состав включаются клеточные диффероны, развивающиеся из разных эмбриональных источников (клетки Лангерганса, меланоциты и др.).
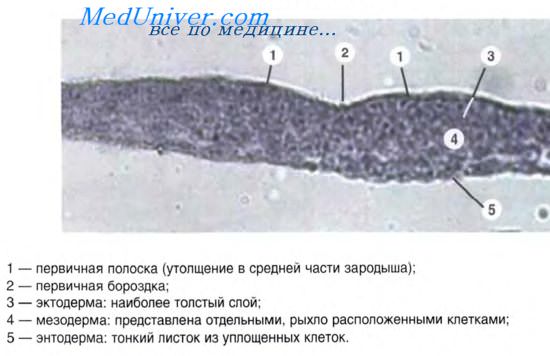
В классификациях эпителия по происхождению за основу, как правило, берется источник развития ведущего клеточного дифферона — дифферона эпителиоцитов. Цитохимическим маркером эпителиоцитов являются белки — цитокератины, образующие тонофиламенты. Цитокератины характеризуются большим разнообразием и служат диагностическим маркером конкретного вида эпителия.
Различают эктодермальные, энтодермальные и мезодермальные эпителии. В зависимости от эмбрионального зачатка, служащего источником развития ведущего клеточного дифферона, эпителии подразделяются на типы: эпидермальный, энтеродермальный, целонефродермальный, эпендимоглиальный и ангиодермальный (Хлопин Н.Г., 1946).
По гистологическим признакам строения ведущего (эпителиального) клеточного дифферона различают однослойные и многослойные эпителии. Однослойные эпителии по форме составляющих их клеток бывают плоские, кубические, призматические или цилиндрические.
Однослойные эпителии подразделяют на однорядные, если ядра всех клеток лежат на одном уровне, и многорядные, в которых ядра расположены на разных уровнях, т. е. в несколько рядов.
Многослойные эпителии подразделяются на ороговевающие и неороговевающие. Многослойные эпителии называют плоскими, учитывая форму клеток наружного слоя. Клетки базального и других слоев могут иметь при этом цилиндрическую или неправильную форму. Кроме названных, выделяют еще переходный эпителий, строение которого меняется в зависимости от степени его растяжения.
На основе данных об органоспецифической детерминации эпителии подразделяются на следующие типы: кожный, кишечный, почечный, целомический и нейроглиальный. В составе каждого типа различают несколько разновидностей эпителиев с учетом их строения и функций. Эпителии перечисленных типов стойко детерминированы. Однако при патологии возможна трансформация одного вида эпителия в другой, но лишь в пределах одного тканевого типа. Например, среди эпителиев кожного типа многорядный мерцательный эпителий воздухоносных путей может переходить в многослойный плоский. Такое явление получило название метаплазия.
Несмотря на разнообразие строения, выполняемых функций и происхождения из разных источников, все эпителии имеют ряд общих признаков, на основе которых их объединяют в систему или группу эпителиальных тканей. Эти общие морфофункциональные признаки эпителиев следующие.
Большинство эпителиев по своей цитоархитектонике представляют собой однослойные или многослойные пласты плотно сомкнутых клеток. Клетки соединены с помощью межклеточных контактов. Эпителий находится в тесных взаимодействиях с подлежащей соединительной тканью. На границе между этими тканями имеется базальная мембрана (пластина). Эта структура участвует в формировании эпителиально-соединительнотканных взаимоотношений, выполняет функции прикрепления с помощью полудесмосом эпителиоцитов, трофическую и барьерную. Толщина базальной мембраны обычно не превышает 1 мкм. Хотя в некоторых органах ее толщина значительно возрастает. Электронно-микроскопически в составе мембраны выделяют светлую (расположенную ближе к эпителию) и темную пластинки. Последняя содержит коллаген IV-ro типа, обеспечивающий механические свойства мембраны. С помощью адгезивных белков — фибронектина и ламинина осуществляется прикрепление эпителиоцитов к мембране. Через базальную мембрану путем диффузии веществ происходит питание эпителия. Базальную мембрану рассматривают в качестве барьера для роста эпителия вглубь. При опухолевых разрастаниях эпителия она разрушается, что позволяет измененным раковым клеткам врастать в подлежащую соединительную ткань (Гаршин В.Г., 1939).
Эпителиоциты обладают гетерополярностью. Строение апикальной и базальной частей клетки разное. В многослойных пластах клетки различных слоев отличаются друг от друга по структуре и функциям. Это называют вертикальной анизоморфией. Эпителии обладают высокой способностью к регенерации за счет митозов камбиальных клеток. В зависимости от местоположения камбиальных клеток в эпителиальных тканях различают диффузный и локализованный камбий.
Читайте также: Как рассчитать расход ткани для постельного белья
ТКАНЕВОЙ КАМБИЙ
ТКАНЕВОЙ КАМБИЙ (лат. cambium смена, обмен) — малодифференцированные клетки, входящие в состав тканей и обладающие способностью к размножению и дифференцировке (специализации).
Термины «камбий», «камбиальные клетки» введены А. А. Заварзиным в 30-х гг. 20 в. С наличием Т. к. связывали физиол. регенерацию тканей. Но, как выяснилось позднее, регенерация (см.) может происходить и без размножения клеток. Наличие камбия не определяет способности к регенерации, а указывает лишь на механизм ее реализации в конкретной ткани. Камбиальные элементы, по А. А. Заварзину, являются не простыми остатками зародышевых листков, а их малодифференцированнымп потомками. Они располагаются либо в виде одиночных клеток среди дифференцированных клеток, напр, в поперечнополосатой мышечной ткани, либо группами — в многослойном плоском эпителии. В нек-рых случаях камбиальные клетки топографически отделены от дифференцированных форм (напр., бластные клетки красного костного мозга от клеток крови).
В настоящее время термины «камбий», «камбиальные клетки» употребляются редко, уступая место более точным определениям — «стволовые клетки» (полииотентные), «полустволовые клетки» (частично детерминированные, или коммутированные). В начале 20 в. А. А. Максимов высказал предположение о существовании в организме стволовой кроветворной клетки. В 00-х гт. 20 в. эта гипотеза подкрепилась экспериментальными данными Тил-ла и Мак-Каллока (J. Е. Till, E. A. McCulloch) о наличии в крови и кроветворных органах полипотентной клетки, по морфол. признакам похожей на лимфоцит. Идентифицированы стволовые клетки и для ряда других тканей. Все они характеризуются наличием диффузного хроматина в ядре, содержанием одного или большего количества ядрышек, слабым развитием органелл цитоплазмы, за исключением свободных рибосом (см. Клетка). Форма стволовых клеток различных систем клеточных дифференцировок неодинакова. В противоположность остальным пролиферирующим клеткам при делении стволовых клеток образуются потомки, к-рые необязательно подвергаются дальнейшей дифференцировки гг поэтому могут сохранять свой пролиферативный потенциал. Существенную роль в дифференцировке потомков стволовых клеток играет микроокружение. Если стволовые клетки — это длительно, медленно самоподдерживающаяся популяция клеток, то коммутированные клетки, берущие начало от стволовых клеток, интенсивно, многократно размножаются и подвергаются медленной дифференцировке, в ходе к-рой на уровне синтеза информационной РНК осуществляется программа их детерминации (см. Гистогенез). Детерминированные клетки перестают реагировать на микроокружение, но не становятся отличными от того тина клеток, к к-рому они принадлежали прежде (напр., класс морфологически распознаваемых пролиферирующих клеток красного костного мозга). В основе этих процессов лежит репрессия определенных генов на уровне трансляции генетического кода молекулы РНК на синтезируемые клеткой тканево-специфические молекулы белка (см. Ген, Трансляция). Для нек-рых частично детерминированных клеток доказана невозможность обратного перехода в стволовые клетки. В отношении других клеток однозначных данных нет, что, возможно, обусловлено неодинаковой степенью жесткости детерминации (программирования) развивающихся клеток и тканей.
Развитие учения о стволовых клетках уточняет прежние представления о Т. к. Разработка методов идентификации стволовых клеток в разных тканях имеет не только теоретический интерес, но и служит решению практически важных проблем, таких как дифференцировка, регенерация и др.
Библиогр.: Данин и Е. С. Понятие о тканевой камбиальности в современной гистологии, Усп. совр. биол., т. 29, № 3, с. 379, 1950; он же, Самообновление тканей и теория камбиальности академика А. А. Заварзина, Арх. анат., гистол., и эмбриол., т. 34, № 1, с. 16, 1957; 3 а-в а р з и н А. А. Курс гистологии и микроскопической анатомии, JI.,1939; он же, Избранные труды, т. 2, с. 217, М.— Л., 1953; Р а й ц и н а С. С. Идентификация стволовых клеток в некоторых системах клеточных дифференцировок у млекопитающих, Усп. совр. биол., т. 90, в. 1(4), с. 123, 1980; Фриденштейн А. Я. и Л у р и я Е. А. Клеточные основы кроветворного микроокружения, М., 1980; Чертков И. Л. и Фриденштейн А. Я. Клеточные основы кроветворения, М., 1977.
Без камбиальная ткань гистология
Эпителиальные ткани — древнейшие гистологические структуры, которые в фило- и онтогенезе возникают первыми. Основное свойство эпителиев — пограничность. Эпителиальные ткани (от греч. epi — над и thele — кожица) располагаются на границах двух сред, отделяя организм или органы от окружающей среды.
Читайте также: Цветочки из ткани для сумки
Эпителии, как правило, имеют вид клеточных пластов и образуют наружный покров тела, выстилку серозных оболочек, просветов органов, сообщающихся с внешней средой во взрослом состоянии или в эмбриогенезе. Через эпителии осуществляется обмен веществ между организмом и окружающей средой. Важной функцией эпителиальных тканей является защита подлежащих тканей организма от механических, физических, химических и других повреждающих воздействий. Некоторые эпителии специализированы на выработке специфических веществ — регуляторов деятельности других тканей организма.
Производными покровных эпителиев являются железистые эпителии. Особый вид эпителия — эпителий органов чувств.
Эпителии развиваются с 3-4-й недели эмбриогенеза человека из материала всех зародышевых листков. Некоторые эпителии, например эпидермис, формируются как полидифферонные ткани, так как в их состав включаются клеточные диффероны, развивающиеся из разных эмбриональных источников (клетки Лангерганса, меланоциты и др.).
В классификациях эпителия по происхождению за основу, как правило, берется источник развития ведущего клеточного дифферона — дифферона эпителиоцитов. Цитохимическим маркером эпителиоцитов являются белки — цитокератины, образующие тонофиламенты. Цитокератины характеризуются большим разнообразием и служат диагностическим маркером конкретного вида эпителия.
Различают эктодермальные, энтодермальные и мезодермальные эпителии. В зависимости от эмбрионального зачатка, служащего источником развития ведущего клеточного дифферона, эпителии подразделяются на типы: эпидермальный, энтеродермальный, целонефродермальный, эпендимоглиальный и ангиодермальный (Хлопин Н.Г., 1946).
По гистологическим признакам строения ведущего (эпителиального) клеточного дифферона различают однослойные и многослойные эпителии. Однослойные эпителии по форме составляющих их клеток бывают плоские, кубические, призматические или цилиндрические.
Однослойные эпителии подразделяют на однорядные, если ядра всех клеток лежат на одном уровне, и многорядные, в которых ядра расположены на разных уровнях, т. е. в несколько рядов.
Многослойные эпителии подразделяются на ороговевающие и неороговевающие. Многослойные эпителии называют плоскими, учитывая форму клеток наружного слоя. Клетки базального и других слоев могут иметь при этом цилиндрическую или неправильную форму. Кроме названных, выделяют еще переходный эпителий, строение которого меняется в зависимости от степени его растяжения.
На основе данных об органоспецифической детерминации эпителии подразделяются на следующие типы: кожный, кишечный, почечный, целомический и нейроглиальный. В составе каждого типа различают несколько разновидностей эпителиев с учетом их строения и функций. Эпителии перечисленных типов стойко детерминированы. Однако при патологии возможна трансформация одного вида эпителия в другой, но лишь в пределах одного тканевого типа. Например, среди эпителиев кожного типа многорядный мерцательный эпителий воздухоносных путей может переходить в многослойный плоский. Такое явление получило название метаплазия.
Несмотря на разнообразие строения, выполняемых функций и происхождения из разных источников, все эпителии имеют ряд общих признаков, на основе которых их объединяют в систему или группу эпителиальных тканей. Эти общие морфофункциональные признаки эпителиев следующие.
Большинство эпителиев по своей цитоархитектонике представляют собой однослойные или многослойные пласты плотно сомкнутых клеток. Клетки соединены с помощью межклеточных контактов. Эпителий находится в тесных взаимодействиях с подлежащей соединительной тканью. На границе между этими тканями имеется базальная мембрана (пластина). Эта структура участвует в формировании эпителиально-соединительнотканных взаимоотношений, выполняет функции прикрепления с помощью полудесмосом эпителиоцитов, трофическую и барьерную. Толщина базальной мембраны обычно не превышает 1 мкм. Хотя в некоторых органах ее толщина значительно возрастает. Электронно-микроскопически в составе мембраны выделяют светлую (расположенную ближе к эпителию) и темную пластинки. Последняя содержит коллаген IV-ro типа, обеспечивающий механические свойства мембраны. С помощью адгезивных белков — фибронектина и ламинина осуществляется прикрепление эпителиоцитов к мембране. Через базальную мембрану путем диффузии веществ происходит питание эпителия. Базальную мембрану рассматривают в качестве барьера для роста эпителия вглубь. При опухолевых разрастаниях эпителия она разрушается, что позволяет измененным раковым клеткам врастать в подлежащую соединительную ткань (Гаршин В.Г., 1939).
Эпителиоциты обладают гетерополярностью. Строение апикальной и базальной частей клетки разное. В многослойных пластах клетки различных слоев отличаются друг от друга по структуре и функциям. Это называют вертикальной анизоморфией. Эпителии обладают высокой способностью к регенерации за счет митозов камбиальных клеток. В зависимости от местоположения камбиальных клеток в эпителиальных тканях различают диффузный и локализованный камбий.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом