
БИОЭЛЕКТРИЧЕСКИЕ ЯВЛЕНИЯ — процессы распределения и транспорта электрических зарядов в живом организме, обусловленные присутствием в живых клетках и тканях большого количества фиксированных (заряженные группы био-макромолекул) и подвижных (свободные ионы и электроны) электрических зарядов. Многие вещества, поступающие в организм и выводимые из него в процессе метаболизма, находятся в виде ионов. Перераспределение и транспорт этих зарядов служат причиной возникновения биоэлектрических потенциалов (см.). Наиболее крутой градиент ионов обнаруживается на клеточных мембранах (см. Мембраны биологические), которые способны эффективно регулировать неравномерное и избирательное распределение ионов. У большинства клеток мембранный потенциал, называемый потенциалом покоя, зависит от градиента ионов калия. Его величина может быть рассчитана по уравнению Нернста:

где R — газовая константа; T — абсолютная температура; n — заряд иона; F — число Фарадея; [К+ ]вн.— концентрация ионов калия внутри клетки; [К+] нар— концентрация ионов калия в наружном растворе. Потенциал покоя имеет знак «минус» на внутренней поверхности мембраны и знак «плюс» на внешней.
В возбудимых образованиях (напр., нервные и мышечные клетки) регистрируется особый вид мембранного потенциала — так наз. потенциал действия, который, в отличие от потенциала покоя, представляет собой пиковый потенциал, передвигающийся в форме волны возбуждения по поверхности клетки со скоростью до нескольких десятков метров в секунду. В возбужденном участке потенциал приобретает обратный знак (т. е. потенциал реверсируется) и внутренняя поверхность заряжается положительно, а наружная — отрицательно. Абсолютная величина потенциала, как правило, превышает уровень потенциала покоя. Согласно ионной теории Ходжкина (A. L. Hodgkin, 1958) возникновение потенциала действия обусловлено избирательным увеличением проницаемости мембраны для ионов натрия.
Основные биоэнергетические процессы в живом организме связаны с биологическим окислением (см. Окисление биологическое), в процессе к-рого электроны движутся по цепочке дыхательных ферментов, расположенных во внутренней мембране митохондрий, по градиенту окислительно-восстановительного потенциала. В соответствии с хемиосмотической гипотезой Митчелла (P. Mitchell, 1966) перенос электронов по дыхательной цепи, звенья которой располагаются поперек мембраны, сопровождается появлением мембранного потенциала.
Живой организм является не только генератором биоэлектричества, но и пассивным проводником электрического тока, возникающего в организме или приложенного извне (см. Электропроводность биологических систем). Электропроводность живых клеток и тканей зависит от импеданса, т. е. суммы активного (омического) и реактивного (емкостного) сопротивлений. Прохождение постоянного электрического тока через живые клетки сопровождается поляризацией, к-рая может быть рассчитана по формуле:
где I — сила тока, R — электрическое сопротивление, U — напряжение тока, P — напряжение поляризации (см. Поляризация). При пропускании переменного тока напряжение поляризации уменьшается с возрастанием частоты. Это явление носит название дисперсии электропроводности (см. Дисперсия). Б. Н. Тарусовым в 1938 г. было показано, что дисперсия электропроводности может служить чувствительным индикатором жизнеспособности клеток и тканей.
Как правило, поверхность живых клеток и тканей несет значительный отрицательный заряд, т. к. ионогенные группы на поверхности клеток, представленные концевыми группами макромолекул клеточной поверхности, при физиологических значениях pH ионизованы. Под действием сил электростатического притяжения положительные ионы из раствора будут притягиваться к поверхности и создавать двойной электрический слой. Часть энергии двойного слоя можно измерить по скорости движения заряженной клетки в электрическом поле. Этот так наз. электрокинетический потенциал (см. Электрокинетические явления) рассчитывается по уравнению Смолуховского: Z = 4πηu/DE, где D — диэлектрическая постоянная среды, η — коэффициент вязкости, E — градиент напряжения, u — электрофоретическая подвижность. В наст, время электрокинетический потенциал используют как индикатор состояния клеточной поверхности. В частности, было показано, что подвижность асцитных клеток несколько ниже по сравнению с нормальными клетками.
К электрокинетическим явлениям относят также потенциал течения. В живом организме имеется постоянное течение жидкости. Всякое движение жидкости (напр., течение крови по артериям и венам) сопровождается появлением потенциала течения между жидкой фазой и стенкой сосуда. Поскольку удельная электропроводность крови велика, эти потенциалы малы — не больше 1 —2 мв. Изменение этого потенциала, вероятно, может иметь и диагностическое значение.
К электрокинетическим явлениям относится и электроосмос, который наблюдается при движении жидкости через пористую перегородку под влиянием внешнего электрического поля. В ряде случаев положительный и отрицательный аномальный осмос в почках объясняется электроосмотическим переносом воды.
Библиография: Катц Б. Нерв, мышца и синапс, пер. с англ., М., 1968, библиогр.; Курелла Г. А. Биоэлектрические потенциалы, в кн.: Биофизика, под ред. Б. Н. Тарусова и О. Р. Колье, с. 238, М., 1968, библиогр.; Скулачев В. П. Трансформация энергии в биомембранах, М., 1972, библиогр.
БИОЭЛЕКТРИЧЕСКИЕ ЯВЛЕНИЯ
Физиологическое состояние живой ткани характеризуется электрическими токами, которые от нее отводятся; это биоэлектрические токи, или биопотенциалы. Различают биотоки трех типов: 1) токи покоя, или клеточные потенциалы, 2) токи, или потенциалы повреждения и 3) токи действия, или акционные токи.
Читайте также: Какое животное культивируют ради получения шелковых тканей
Токи покоя, или клеточные потенциалы отводятся от живых неповрежденных клеток. Когда нет раздражения, а значит и возбуждения, имеется разность потенциалов между наружной поверхностью оболочки (мембраны) живой клетки, заряженной положительно, и внутренней ее поверхностью, или цитоплазмой, заряженной отрицательно. Для отведения потенциала покоя внутрь клетки вводится микроэлектрод, имеющий внешний диаметр менее 0,5 мкм (микрометра, или микрона), а на наружную поверхность клетки накладывается микроэлектрод, имеющий внешний диаметр в 1—4 мкм.
Величина токов покоя в разных клетках различна и достигает 100 же (милливольт) и больше.
Токи, или потенциалы повреждения отводятся от поврежденного и неповрежденного участков живой ткани, так как поврежденный участок заряжен отрицательно, а неповрежденный в отсутствии возбуждения— положительно. Они являются реакцией живой протоплазмы на повреждение и изменяются во времени и пространстве, снижаясь сразу после повреждения от нескольких десятков милливольт до нескольких милливольт.
Токи действия, или акционные токи имеют большое значение для величины возбуждения и его распространения. Различают однофазные и двухфазные токи действия.
Однофазный ток действия отводится от ограниченного участка ткани при очень слабом подпороговом раздражении. Он является низковольтным, местным, нераспространяющимся. Чем больше сила подпорогового раздражителя, тем больше величина однофазного тока действия. Следовательно, однофазный ток действия характеризует очень слабое местное возбуждение с сохранением возбудимости.
Двухфазный ток действия отводится от ткани, раздражаемой пороговыми и надпороговыми раздражителями. При усилении раздражителя от подпорогового до порогового возбуждение возрастает, достигает критической величины, которая в дальнейшем не изменяется и начинает волнообразно распространяться в обе стороны от раздражаемого участка. При этом отводится высоковольтный потенциал, который в нервных волокнах в 10 раз выше порога возбудимости.
Высоковольтный потенциал распространяющегося тока действия, или пик, имеет почти одинаковую высоту (амплитуду) на всем протяжении возбудимой ткани, но высота пика различна у разных тканей и у нервных волокон разного строения и диаметра. Продолжительность отдельного пика также одинакова на всем протяжении возбудимой ткани. Амплитуда и длительность пика изменяются в зависимости от физиологического состояния ткани и скорости проведения возбуждения.
На высоте пика живая ткань находится в состоянии максимального возбуждения, и если нанести в это время повторное раздражение, то оно не вызовет нового возбуждения и не произведет никакого изменения деятельности ткани или органа. Этот период отсутствия эффекта обозначается как абсолютная рефрактерная фаза. Продолжительность этой фазы в нервных волокнах от 0,4 до 2 мсек. После пика регистрируется низковольтный отрицательный следовой потенциал, который отличается от пика неустойчивостью. Во время протекания низковольтного отрицательного потенциала, достигающего всего 0,05 потенциала пика и продолжающегося в нервных волокнах от 1 до 10 мсек, возбудимость постепенно восстанавливается. Это фаза относительной рефрактерности, в которой ткань или орган реагируют на раздражение, но ниже обычного уровня. И, наконец, фаза относительной рефрактерности переходит в фазу повышенной возбудимости, которая обозначается как супернормальный период, или экзальтационная фаза, во время которой орган реагирует на раздражение значительно выше исходного уровня. Эта фаза совпадает с низковольтным положительным следовым потенциалом, составляющим приблизительно 0,002 потенциала пика.
Рефрактерность — свойство всех возбудимых тканей, но в разных тканях ее продолжительность различна, например, в скелетной мышце она продолжается больше, чем в нервах, а в сердечной мышце она продолжается еще дольше. Благодаря абсолютной и относительной рефрактерным фазам возбуждение проводится ритмически, отдельными волнами, или импульсами, так как в продолжение абсолютной рефрактерной фазы возбудимость отсутствует. В этой фазе возбуждение переходит в торможение. Следовательно, нерв может проводить не больше 2000 импульсов в секунду, но в естественных условиях при температуре тела он проводит не больше 200 импульсов в секунду.
Лабильность. Н. Е. Введенский обнаружил, что живые ткани обладают свойством лабилыюсти, или функциональной подвижности. В одних тканях возбуждение быстро возникает и быстро прекращается, в других — медленно возникает, медленно протекает и сравнительно долго не прекращается. Лабильность измеряется числом волн возбуждения в одну секунду.
Следовательно, в тканях, обладающих большой или высокой лабильностью, в течение одной секунды проводится значительно больше волн возбуждения, чем в тканях, которым свойственна малая или низкая лабильность.
Торможение. Когда возбуждение, проявляющееся в деятельности ткани или органа, возрастает до критической величины, то при определенных условиях оно переходит в торможение, проявляющееся во временном активном задерживании деятельности ткани или органа. Н. Е. Введенский обнаружил, что переход возбуждения в торможение происходит постепенно, через несколько стадий. Вначале наблюдается уравнительная стадия, во время которой слабые и сильные раздражители вызывают одинаковую по величине деятельность органа или ткани, например, одинаковой высоты сокращение скелетной мышцы. По мере усиления торможения возникает вторая стадия — парадоксальная. В этой стадии сильные раздражители тормозят работу органа, а слабые вызывают возбуждение и, следовательно, работу органа. И, наконец, наступает последняя стадия — тормозная, в которой и слабые и сильные раздражители вызывают торможение. При переходе торможения в возбуждение эти стадии развиваются в обратном порядке.
Читайте также: Конспект занятия по аппликации из ткани в старшей группе
Явление перехода возбуждения в торможеиие и наоборот Н. Е. Введенский назвал парабиозом.
Единство возбуждения и торможения. Возбуждение и торможение, как следует из сказанного, не являются процессами, существующими независимо друг от друга, а представляют собой две взаимосвязанные фазы единого нервного процесса. При определенных условиях раздражения и состояния лабильности ткани возникает возбуждение, а при других условиях раздражения и состояния лабильности ткани возбуждение переходит в торможение. Для каждой ткани существуют наиболее благоприятные ритм, частота и сила раздражения, вызывающие максимально возможное ее возбуждение и деятельность. Н. Е. Введенский назвал наиболее благоприятную частоту раздражения оптимумом частоты, наиболее благоприятную силу раздражения — оптимумом силы, а чрезмерную частоту или силу раздражения, при которой возникает-торможение, он назвал пессимумом частоты и пессимумом силы. В настоящее время различают несколько других видов торможения в нервной системе.
Башкирский государственный медицинский университет
Нормальная физиология
Теория
Биоэлектрические явления в живых тканях. Потенциал покоя
В 1949-1952 гг. Ходжкин, Хаксли и Катц модифицировали существовавшие ранее теории и экспериментально обосновали основные положения мембранно-ионной теории .
Согласно этой теории возникновение МП (мембранного потенциала) обусловлено:
- Разной концентрацией ионов калия, натрия, кальция и хлора внутри клетки и во внеклеточной среде.
- Разной проницаемостью мембраны клеток для этих ионов в покое и при возбуждении.
Состояние покоя — это такое состояние, когда отсутствует раздражающее действие на организм. Это состояние относительное, т.к. абсолютный покой не совместим с жизнью.
Деятельное состояние — это изменение состояния организма, возникающее под действием факторов внешней среды.
Раздражимость — это способность организма отвечать на действие раздражающих факторов изменением его структурных и функциональных свойств.
Возбудимость
Возбудимость — способность ткани отвечать на раздражение специализированной реакцией — возбуждением. Возбудимость является одной из форм раздражимости.
Возбудимыми тканями называются такие ткани, клетки которых способны в ответ на раздражение генерировать специфическую реакцию — возбуждение .
Возбуждение — это специализированная ответная реакция живого объекта на действие раздражителя, проявляющаяся в изменениях его определенных параметров.
К возбудимым тканям относятся:
Общие признаки возбуждения (присущи всем возбудимым тканям):
- Изменение уровня обменных процессов в тканях;
- Выделение различных видов энергии — тепловой, электрической.
Специфические признаки возбуждения (характерны для определенного вида тканей):
- Мышечная ткань — сокращение,
- Железистая — выделение секрета,
- Нервная — генерация и проведение нервного импульса.
- эпителиальная,
- соединительная ткани.
Физиологические свойства возбудимых тканей
Основными свойствами всех возбудимых тканей являются:
Лабильность (функциональная подвижность) — это способность ткани реагировать на различные воздействия с определенной скоростью, то есть способность воспроизводить определенную частоту раздражения.
Мерой лабильности является наибольшее число ответных реакций, которыми возбудимая ткань способна реагировать за единицу времени в соответствии с частотой приложенных к ней раздражений.
Проводимость — это способность ткани и клетки передавать возбуждение.
Раздражение — это процесс воздействия на живую ткань агентов внешней по отношению к этой ткани среды.
Раздражитель — это причина, способная вызвать ответную реакцию.
Классификация раздражителей
- физические,
- химические,
- физико-химические,
- биологические.
По биологическому значению раздражители делятся на 2 группы :
- Адекватные — раздражители, к воздействию которых ткани в процессе эволюции приспособлены в наибольшей степени.
- Неадекватные — раздражители, к воздействию которых возбудимые ткани специально не приспособлены.
Состав плазматической мембраны
- Липиды (в основном фосфолипиды),
- Белки (гликопротеины),
- Углеводы (мукополисахариды).
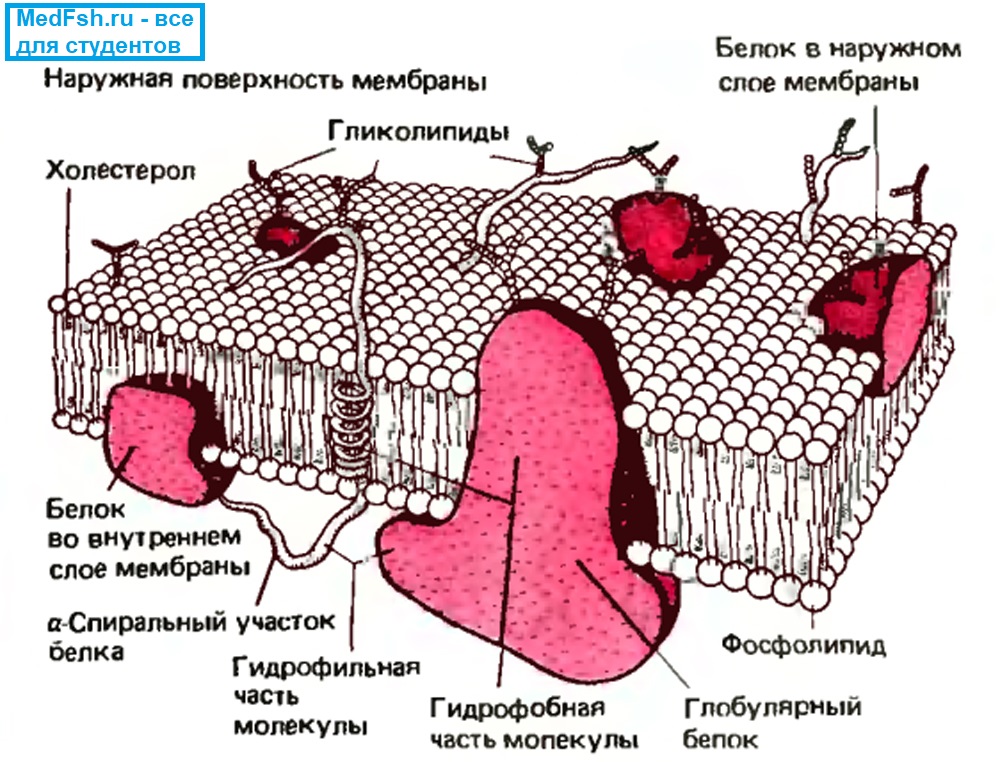
Липиды очень плотно упакованы в мембране, между ними нет практически никаких расстояний, поэтому мембрана плохо пропускает воду, практически непроницаема для ионов и других крупных молекул.
Белковые молекулы могут быть погружены в слой липидов с внеклеточной или с цитоплазматической стороны, либо могут целиком пронизывать мембрану.
Если белки прикреплены к поверхности мембраны, их называют периферическими. С внутренней стороны это будут белки-ферменты, а с наружной — белки-рецепторы.
Если белки пронизывают всю толщу мембраны клеток, то их называют интегральными или трансмембранными.
Такие белки образуют структуры, обеспечивающие движение ионов через мембрану.
Если белки образуют стенки поры, сквозь которую путем простой диффузии проходят ионы, то это ионные каналы.
Читайте также: Флис это синтетика или натуральная ткань
Если трансмембранные белки перекачивают ионы против концентрационного и электрического градиентов, то это ионные насосы.
Все каналы, имеющиеся в живых тканях, можно разделить на 2 типа:
- первый тип — каналы покоя, которые спонтанно открываются и закрываются без всяких внешних воздействий;
- второй тип — gate-каналы (воротные каналы) — в покое они закрыты и открываются под действием раздражителей.
- неспецифические (каналы утечки, всегда открыты),
- специфические (селективные), обладающие способностью пропускать только определенные ионы при изменении заряда на мембране или действии химических веществ.
Транспорт веществ
Транспорт частиц через каналы является жизненно важным для клеток процессом.
Обычно транспорт веществ делят на пассивный (без затраты энергии), т.е. транспорт веществ по концентрационному, осмотическому и электрохимическому градиентам и активный (с затратой энергии).
Различают первично- и вторично-активный транспорт
Первично-активный транспорт ионов обеспечивается специальными ионными насосами, осуществляется с затратой энергии АТФ, против концентрационного градиента, т.е. перенос веществ происходит из меньшей концентрации через мембрану в большую концентрацию.
- Это вид транспорта для переноса веществ (глюкозы, аминокислот и др.) через мембрану также против градиента, но без затраты энергии .
- Эти вещества проходят через мембрану с помощью специальных переносчиков (напр. ионов Na), на транспорт которых и затрачивается энергия, а эти вещества движутся как бы попутно.
Мембранный потенциал или потенциал покоя
В состоянии покоя между наружной и внутренней поверхностью мембраны возникает разность зарядов или потенциалов, которая в дальнейшем была названа мембранным потенциалом (МП) или потенциалом покоя (ПП) .
Положительные заряды концентрируются на наружной поверхности мембраны, а отрицательные заряды — на внутренней поверхности.
Мембранный потенциал измеряется в отрицательных значениях, т.к. внутренняя поверхность мембраны заряжена отрицательно. Его величина колеблется от -60 до -90 мВ у разных клеток.
Методы измерения мембранного потенциала
В зависимости от места приложения электродов :
- внеклеточный с помощью макроэлектродов,
- внутриклеточный с помощью микроэлектродов.
1. Исследование ПП с помощью макроэлектродов осуществляют, прикладывая один из них к неповрежденному , а другой — к поврежденному участку ткани.
Микроэлектрод представляет собой микропипетку диаметром 0,5 — 1 мкм, наполненную концентрированным солевым раствором (KCl). Внутри него может также находится неполяризующийся электрод — из серебра, золота или платины.
Второй электрод помещается во внеклеточную жидкость.
Оба электрода присоединяются к усилителю и осциллографу для регистрации потенциала. В момент прокалывания мембраны, осциллограф регистрирует появление отрицательного потенциала , соответствующего ПП.
1-ая причина — ионная асимметрия:
- Ионов K находится в клетке в 30-50 раз больше, чем снаружи;
- Ионов Na больше находится вне клетки, чем внутри нее в 8-10 раз;
- Ионов Ca во много раз больше вне клетки;
- Ионов Cl также больше во внеклеточной жидкости, чем внутри клетки в 50 раз;
- Внутри клетки находится больше органических анионов по сравнению с наружной поверхностью.
Таким образом, для этих ионов направление концентрационного градиента различно!
- Для K из клетки (из большей концентрации к меньшей);
- Для Na, Ca и Cl в клетку.
2-ая причина, обуславливающая поляризацию мембраны:
- различная проницаемость мембраны для различных ионов. В состоянии покоя мембрана в 25 раз более проницаема для ионов K, чем для Na, т.к. количество калиевых каналов на единицу площади мембраны намного больше, чем натриевых.
- Поскольку концентрация ионов K в цитоплазме намного выше, чем снаружи клетки, они начинают двигаться через канал и выходят из клетки.
- Ионы K несут положительные заряды, поэтому мембрана снаружи заряжается положительно.
Отрицательный заряд на внутренней поверхности мембраны обусловлен наличием органических анионов — крупномолекулярных соединений, которые заряжены отрицательно, и для которых мембрана непроницаема (глютамат, аспартат, органические фосфаты, сульфаты и др.)
Таким образов, на мембране формируется калиево-равновесный потенциал, т.к. уравновешиваются силы диффузии (выход K из клетки по градиенту концентрации) и электростатического взаимодействия (отталкивание выходящих ионов K положительным зарядом на наружной поверхности мембраны).
Калий является основным ионом, обеспечивающим формирование МП (ПП), что подтверждается формулой Нернста . По ней можно, зная концентрацию онов калия внутри и снаружи клетки, подсчитать величину ПП.

Натрий-калиевый насос или помпа
Для непрерывного активного переноса ионов Na и ионов K через мембрану клетки против концентрационного градиента существует Na-Л обменный насос. При каждом цикле он переносит 3 иона Na наружу и 2 иона K внутрь клетки.
Таким образом, МП представляет собой разность потенциалов между наружной и внутренней поверхностью мембраны клетки. Он является результатом разделения зарядов относительно клеточной мембраны , которое возникает за счет движения заряженных ионов (натрия и, преимущественно, калия) по концентрационным градиентам через ионные каналы, открытые в покое и за счет работы Na-K насоса .
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом