
Биоэлектрические явления в живых тканях. Природа возбуждения.
Первые попытки последовательной разработки учения о «животном электричестве» связаны с именем Л. Гальвани. Он обратил внимание на сокращение мышц препарата задних лапок лягушки, подвешенного на медном крючке, при прикосновении лапок к железным перилам балкона. На основании этих наблюдений Л. Гальвани пришел к выводу, что сокращение мышц лапок вызвано «животным электричеством», которое возникает в спинном мозге и передается по металлическим проводникам к мышцам лапки. Этот опыт в настоящее время известен как первый опыт Гальвани.
Физик Л. Вольта, повторив первый опыт Гальвани, пришел к заключению, что описанные явления нельзя считать обусловленными наличием» «животного электричества». Источником тока по мнению А. Вольта является не спинной мозг, как полагал Л. Гальвани, а разность потенциалов, образующаяся в месте контакта разнородных металлов — меди и железа. В ответ на зги возражения Л. Гальвани усовершенствовал опыт, исключив из него металлы. Он препарировал седалищный нерв вдоль бедра лапки лягушки, затем набрасывал нерв на мышцы голени — возникало сокращение мышцы. Этот опыт известен как второй опыт Гальвани, или опыт без металлов.
Позже было замечено, что сокращение мышцы во втором опыте Гальвани возникает, если нерв одновременно соприкасается с поврежденной и неповрежденной поверхностями мышцы. Дюбуа-Рей-моном было установлено, что поврежденный участок мышцы несет отрицательный заряд, а неповрежденный участок — положительный. При набрасывании нерва на поврежденный и неповрежденный участки мышцы возникает ток, который раздражает нерв и вызывает сокращение мышцы. Этот ток был назван током покоя или током повреждения.
Дюбуа-Реймон, таким образом, впервые показал, что наружная поверхность мышцы заряжена положительно по отношению к ее внутреннему содержимому. Следовательно, в состоянии покоя между наружной и внутренней поверхностями мембраны клетки существует разность потенциалов, которая затем была названа мембранным потенциалом покоя или мембранным потенциалом. Его величина у разных клеток колеблется от 60 до 90 мВ.
Было разработано несколько теорий возникновения и поддержания мембранного потенциала покоя. В 1949-52 гг. Ходжкин, Хаксли, Катц модифицировали и экспериментально обосновали мембранно-ионную теорию. Согласно этой теории мембранный потенциал покоя (МПП) обусловлен неодинаковой концентрацией ионов натрия, калия, кальция, хлора внутри клетки и во внеклеточной жидкости, а также неодинаковой проницаемостью для этих ионов поверхностной мембраны клетки. Цитоплазма нервных и мышечных клеток содержит в 30-50 раз больше ионов калия, в 8-10 раз меньше ионов натрия и в 50 раз меньше ионов хлора, чем внеклеточная жидкость. Следовательно, в состояний покоя существует асимметрия концентрации ионов внутри клетки ив окружающей ее среде.
Клетку ограничивает тончайшая оболочка — мембрана. Функции – пограничная (цитоплазма от внеклеточной жидкости)
Биотрансформирующая (барьер для веществ идущих в клетку)
Рецепторная ( мембрана имеет набор структур участвующих в специальных определений физических и химических факторов)
Мембрана образует межклеточные контакты
Транспортная ( переносит разм в-ва в связи с изменение проницаемости)
В состав мембраны входят липиды (в основном фосфолипиды), белки и мукополисахариды. Согласно жидкостно-мозаичной модели мембраны она состоит из бимолекулярного слоя фосфолипидов, в который включены белки. Одни белки пронизывают мембрану насквозь, а другие погружены в ее толщу. В мембране имеются ионные каналы, образованные макромолекулами белка, пронизывающих липидный слой. Каналы мембраны делятся на неспецифические (каналы утечки) и специфические (селективные, обладающие способностью пропускан только определенные ионы). Неспецифические каналы пропускают различные ионы и открыты постоянно. Специфические каналы открываются и закрываются в ответ на изменения МПП. Эти каналы называются потенциалозависимыми.
Селективные потенциалозависмые ионные каналы подразделяются на: натриевые, калиевые, кальциевые и хлорные. Однако их селективность часто не абсолютна, а название канала указывает лишь на тот .ион, для которого данный канал наиболее проницаем.
Ионный канал состоит из собственно канала (транспортной части) и воротного механизма («ворот»), который управляется электрическим полем мембраны. В каждом канале предполагают наличие двух типов «ворот» — быстрых активационных (m) и медленных инактивационных (h). «Ворота» могут быть полностью открыты или закрыты. Например, в натриевом канале в состоянии покоя «ворота» m закрыты, а «ворота» h — открыты. При уменьшении заряда мембраны (деполяризации) в начальный момент «ворота» m и h открыты — канал находится в проводящем состоянии- Через открытые каналы ионы движутся по концентрационному и электрохимическому градиенту. Затем инактивационные «ворота» закрываются — канал инак-тивируется. По мере восстановления МПП инактивационные «ворота» медленно открываются, а активационные быстро закрываются и канал возвращается в исходное состояние.
Мукополисахариды, располагаясь в виде «деревьев» на поверхности мембраны, осуществляют рецепторные функции.
В состоянии физиологического покоя мембрана нервных волокон в 25 раз более проницаема для ионов калия, чем для ионов натрия.
Поляризация мембраны при открытых калиевых каналах и наличии трансмембранного градиента концентраций калия, объясняется прежде всего утечкой внутриклеточного калия в окружающую клетку среду. Выход положительно заряженных ионов калия приводит к появлению положительного заряда на наружной поверхности мембраны. Органические анионы — крупномолекулярные соединения, которые несут отрицательный заряд, и для которых мембрана клетки непроницаема, придают в этих условиях внутренней поверхности мембраны отрицательный заряд.
Читайте также: Ацетат ткань из чего получают
В состоянии покоя наблюдаются небольшие потоки ионов калия и натрия (калия больше, чем натрия) через мембрану по их концентрационному градиенту, что в конечном итоге должно было бы привести к выравниванию концентрации этих ионов внутри клетки и в окружающей ее среде. Но в живых клетках этого не происходит, так как в клеточной мембране существует особый молекулярный механизм, который получил название натрий-калиевого насоса. Он обеспечивает выведение из цитоплазмы клетки ионов натрия и введении в цитоплазму ионов калия. Ионный насос перемещает ионы против их концентрационного градиента, следовательно, он работает с затратой энергии.
Таким образом, возникновение и поддержание мембранного потенциала покоя обусловлено избирательной проницаемостью мембраны клетки и работой натрий-калиевого насоса. Мембранный потенциал покоя создает электрическое поле. Электрическое поле мембранного потенциала покоя обеспечивает закрытое состояние активационных «ворот» натриевых каналов и открытое состояние инактивационных «ворот».
Регистрация электрических потенциалов в нервном и мышечном волокне или в нервной клетке показала, что при возбуждении происходит изменение МПП, возникает потенциал действия. Под влиянием раздражителя. пороговой или сверхпороговой величины проницаемость мембраны клетки для ионов натрия возрастает. Ионы натрия устремляются внутрь клетки, что приводит к уменьшению величины мембранного потенциала покоя — деполяризация мембраны (рис. 2). В начале деполяризация развивается медленно. При уменьшении МПП до критического уровня деполяризации проницаемость мембраны для ионов натрия увеличивается в 500 раз и превышает проницаемость для ионов калия в 20 раз. В результате проникновения ионов натрия в цитоплазму и их взаимодействия с анионами разность потенциалов на мембране исчезает, а затем происходит перезарядка клеточной мембраны (инверсия заряда) — внутренняя поверхность мембраны заряжается положительно по отношению к ее наружной. Этот потенциал превышения достигает величины 30-50 мВ, после чего закрываются быстрые натриевые каналы — происходит инактивация натриевой проницаемости) и открываются калиевые каналы. Начинается процесс восстановления исходного уровня мембранного потенциала покоя — реполяризация мембраны.
При внутриклеточном отведении регистрируются следующие состояния мембраны:
• местное возбуждение, локальный ответ (начальная деполяризация мембраны);
• деполяризация мембраны (восходящая часть спайка, включая инверсию);
• реполяризация мембраны (нисходящая часть потенциала действия);
• следовая деполяризация (соответствует отрицательному следовому потенциалу);
• следовая гиперполяризация (соответствует положительному следовому потенциалу).
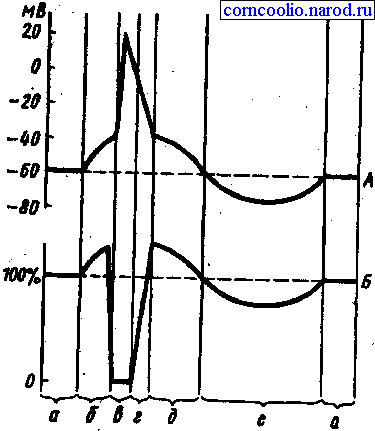
Рис. 2. Соотношение одиночного цикла возбуждения (А) и фаз возбудимости (Б). А: а — мембранный потенциал покоя; б — предспайк, локальный ответ, ВПСП; в — спайк, потенциал действия, деполяризация и инверсия; г — потенциал действия, реполяризация; д — отрицательный следовой потенциал, следовая деполяризация; е — положительный следовой потенциал, следовая гиперполяризация. Б: а — исходный уровень возбудимости; б — фаза первичной экзальтации, повышенная возбудимость; в — фаза абсолютной рефрактерности; г — фаза относительной рефрактернности; д — фаза вторичной экзальтации; е — фаза вторичной рефрактерности
Изменение возбудимости при возбуждении. При развитии потенциала действия происходит изменение возбудимости ткани, причем, это изменение протекает но фазам (рис. 2). Состоянию исходной поляризации мембраны, которую отражает мембранный потенциал покоя, соответствует исходное состояние ее возбудимости и, следовательно, клетки — это нормальный уровень возбудимости. В период предспайка возбудимость ткани повышена, эта фаза возбудимости получила название первичной экзальтации. Во время развития предспайка мембранный потенциал покоя приближается к критическому уровню деполяризации и для достижения последнего достаточна сила раздражителя меньшая, чем пороговая (подпороговая).
В период развития спайка (пикового потенциала) идет лавинообразное поступление ионов натрия внутрь клетки, в результате чего происходит перезарядка мембраны и она утрачивает способность отвечать возбуждением на раздражители даже сверхпороговой силы. Эта фаза возбудимости получила название абсолютной рефрактерности (абсолютной невозбудимости). Она длится до конца перезарядки мембраны. Абсолютная рефрактерность, т. е. полная невозбудимость мембраны возникает в связи с тем, что натриевые каналы в начале полностью открываются, а затем инактивируются.
После окончания фазы перезарядки мембраны возбудимость ее постепенно восстанавливается до исходного уровня — фаза относительной рефрактерности. Она продолжается до восстановления заряда мембраны до величины, соответствующей критическому уровню деполяризации. Так как в этот период мембранный потенциал покоя еще не восстановлен, то возбудимость ткани понижена и новое возбуждение может возникнуть только при действии сверхпорогового раздражителя. Снижение возбудимости в фазу относительной рефрактерности связано с частичной инактивацией натриевых каналов и активацией калиевых.
Периоду отрицательного следового потенциала соответствует повышенный уровень возбудимости — фаза вторичной экзальтации. Так как мембранный потенциал в эту фазу ближе к критическому уровню деполяризации, но сравнению с состоянием покои (исходной поляризацией) , то порог раздражения снижен, т. е. возбудимость повышена. В эту фазу новое возбуждение может возникнуть при действии раздражителей подпороговой силы. Натриевые каналы в эту фазу инактивированы не полностью. В период развития положительного следового потенциала возбудимость ткани понижена — фаза вторичной рефрактерности. В эту фазу мембранный потенциал увеличивается (состояние гиперполяризации мембраны), удаляясь от критического уровня деполяризации, порог раздражения повышается, и новое возбуждение может возникнуть только при действии раздражителей сверхпороговой величины. Гиперполяризация мембраны развивается вследствие трех причин: во-первых, продолжающимися выходом ионов калия; во-вторых, открытием, возможно, каналов для хлора и поступление этих ионов в цитоплазму клетки; в-третьих, усиленной работой натрий-калиевого насоса.
Читайте также: Какие ватрушки лучше ткань или пвх
Биоэлектрические явления в возбудимых тканях. Природа возбуждения
Биоэлектрические явления в возбудимых тканях. Природа возбуждения
Возбуждение представляет собой сложную совокупность физических, химических и физико-химических процессов, в результате которых происходит быстрое и кратковременное изменение электрического потенциала мембраны.
Первые исследования электрической активности живых тканей были проведены Л. Гальвани. Он обратил внимание на ссн сращение мышц препарата задних лапок лягушки, подвешенной на медном крючке, при соприкосновении с железными перилами балкона (первый опыт Гальвани). На основании этих наблюдений им был сделал вывод, что сокращение лапок вызвано «животным электричеством», которое возникает в спинном мозге и передается по металлическим проводникам (крючку и перилам) к мышцам.
Физик А. Вольта, повторив этот опыт, пришел к другому заключению. Источником тока, по его мнению, является не спинной мозг и «животное электричество», а разность потенциалов, образующаяся в месте контакта разнородных металлов – меди и железа, а нервно-мышечный препарат лягушки является лишь проводником электричества. В ответ на эти возражения Л. Гальвани усовершенствовал опыт, исключив из него металлы. Он препарировал седалищный нерв вдоль бедра лапки лягушки, затем набрасывал нерв на мышцы голени, что вызывало сокращение мышцы (второй опыт Гальвани), тем самым доказав существование «животного электричества».
Позднее Дюбуа-Реймоном было установлено, что поврежденный участок мышцы имеет отрицательный заряд, а неповрежденный участок – положительный. При набрасывании нерва между поврежденным и неповрежденным участками мышцы возникает ток, который раздражает нерв и вызывает сокращение мышцы. Этот ток был назван током покоя, или током повреждения. Так было показано, что наружная поверхность мышечных клеток заряжена положительно по отношению к внутреннему содержимому.
В состоянии покоя между наружной и внутренней поверхностями мембраны клетки существует разность потенциалов, которая называется мембранным потенциалом [МП), или, если это клетка возбудимой ткани, – потенциалом покоя. Так как внутренняя сторона мембраны заряжена отрицательно по отношению к наружной, то, принимая потенциал наружного раствора за нуль, МП записывают со знаком «минус». Его величина у разных клеток колеблется от минус 30 до минус 100 мВ.
Первая теория возникновения и поддержания мембранного потенциала была разработана Ю. Бернштейном (1902). Исходя из того, что мембрана клеток обладает высокой проницаемостью для ионов калия и малой проницаемостью для других ионов, он показал, что величину мембранного потенциала можно определить, используя формулу Нернста.
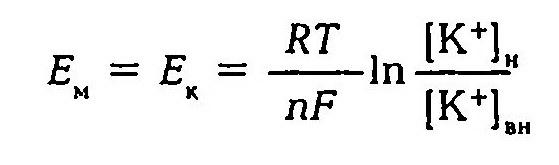
В 1949–1952 гг. А. Ходжкин, Э. Хаксли, Б. Катц создали со-временную мембранно-ионную теорию, согласно которой мембранный потенциал обусловлен не только концентрацией ионов калия, но и натрия и хлора, а также неодинаковой проницаемостью для этих ионов мембраны клетки. Цитоплазма нервных и мышечных клеток содержит в 30 -50 раз больше ионов калия, в 8–10 раз меньше ионов натрия и в 50 раз меньше ионов хлора, чем внеклеточная жидкость. Проницаемость мембраны для ионов обусловлена ионными каналами, макромолекулами белка, пронизывающими липидный слой. Одни каналы открыты постоянно, другие (потенциалозависимые) открываются и закрываются в ответ на изменения МП. Потенциалозависимые каналы подразделяются на натриевые, калиевые, кальциевые и хлорные. В состоянии физиологического покоя мембрана нервных клеток в 25 раз более проницаема для ионов калия, чем для ионов натрия.
Таким образом, согласно обновленной мембранной теории асимметричное распределение ионов по обе стороны мембраны и связанное с этим создание и поддержание мембранного потенциала обусловлено как избирательной проницаемостью мембраны для различных ионов, так и их концентрацией по обе стороны от мембраны, а более точно величину мембранного потенциала можно рассчитать по формуле.
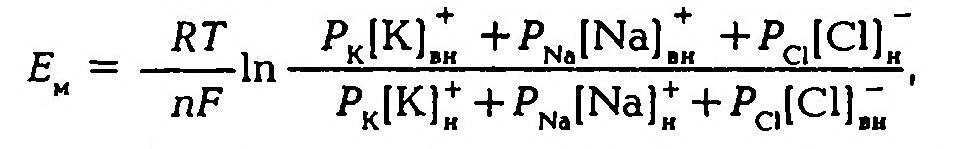
Поляризация мембраны в покое объясняется наличием открытых калиевых каналов и трансмембранным градиентом концентраций калия, что приводит к выходу части внутриклеточного калия в окружающую клетку среду, т. е. к появлению положительного заряда на наружной поверхности мембраны. Органические анионы – крупномолекулярные соединения, для которых мембрана клетки непроницаема, создают на внутренней поверхности мембраны отрицательный заряд. Поэтому чем больше разница концентраций калия по обе стороны от мембраны, тем больше его выходит и тем выше значения МП. Переход ионов калия и натрия через мембрану по их концентрационному градиенту в конечном итоге должен был бы привести к выравниванию концентрации этих ионов внутри клетки и в окружающей ее среде. Но в живых клетках этого не происходит, так как в клеточной мембране имеются натрий-калиевые насосы, которые обеспечивают выведение из клетки ионов натрия и введение в нее ионов калия, работая с затратой энергии. Они принимают и прямое участие в создании МП, так как за единицу времени ионов натрия выводится из клетки больше, чем вводится калия (в соотношении 3:2), что обеспечивает постоянный ток положительных ионов из клетки. То что выведение натрия зависит от наличия метаболической энергии, доказывается тем, что под действием динитрофенола, который блокирует метаболические процессы, выход натрия снижается примерно в 100 раз. Таким образом, возникновение и поддержание мембранного потенциала обусловлено избирательной проницаемостью мембраны клетки и работой натрий-калиевого насоса.