
Биосинтез фосфолипидов по сравнению с синтезом ТАГ имеет существенные особенности. Они заключаются в дополнительной активаци и компонентов ФЛ – фосфатидной кислоты или холина и этаноламина.
1 путь – «спасательный»
Благодаря этому пути холин и этаноламин используются повторно и не катаболизируют. Активация холина (или этаноламина ) происходит через промежуточное образование фосфорилированных производных с последующим присоединением ЦМФ. В следующей реакции фосфохолин (или фосфоэтаноламин) переносится на ДАГ. Этот путь особенно характерен для легких и кишечника, но идет и в других тканях.
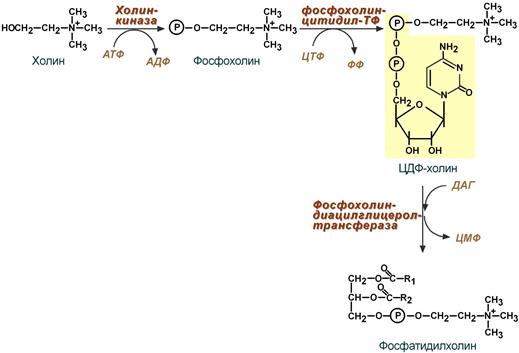
Реакции синтеза фосфолипидов
с использованием 1,2-ДАГ на примере фосфатидилхолина
2 путь – основной, синтез de novo
Здесь холин (или этаноламин) не встраиваются в готовом виде, а образуются уже в молекуле фосфолипида.
Активация фосфатидной кислоты заключается в присоединении к ней ЦМФ с образованием ЦДФ-ДАГ. Далее к нему присоединяется шестиатомный спирт инозитол или серин с образованием фосфатидилинозитола и фосфатидилсерина . Синтезированный фосфатидилсерин подвергается декарбоксилированию с образованием фосфатидилэтаноламина . Последний метилируется при участии S-аденозилметионина в фосфатидилхолин .
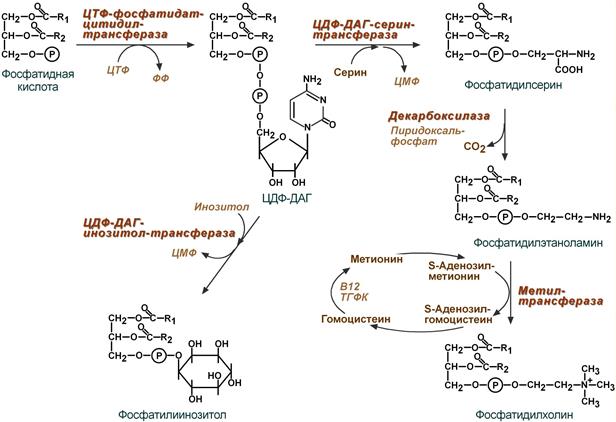
Реакции синтеза фосфолипидов
с использованием фосфатидной кислоты
3 путь – обратное превращение
Между фосфатидилэтаноламином и серином может происходить реакция с образованием в результате реакции фосфатидилсерина и свободного этаноламина .
Липотропные вещества
Все вещества, способствующие синтезу ФЛ и препятствующие синтезу ТАГ, и способные предотращать жировую инфильтрацию печени, называются липотропными факторами . К ним относятся:
1. Структурные компоненты фосфолипидов: полиненасыщенные жирные кислоты, инозитол, серин, холин, этаноламин.
2. Метионин – в виде S-аденозилметионина является донором метильных групп для синтеза холина и фосфатидилхолина.
- пиридоксин (В6), способствующий образованию ФЭА из ФС.
- цианкобаламин (В12) и фолиевая кислота, участвующие в реакциях обмена серина, глицина и метионина при ресинтезе метионина из гомоцистеина. Следовательно они, хотя и не напрямую, но необходимы для синтеза фосфатидилхолина.
Биосинтез фосфолипидов в тканях
• Основные клеточные фосфолипиды синтезируются в мембране ЭПР, главным образом, со стороны цитозоля.
• Регуляция синтеза новых липидов контролируется локализацией ферментов, участвующих в этом процессе.
• Биосинтез холестерина регулируется протеолизом фактора транскрипции, который интегрирован в мембрану ЭПР.
Наряду с транслокацией и подготовкой белков к секреторному процессу, эндоплазматический ретикулум (ЭПР) функционирует как место первичного образования клеточных фосфолипидов. Эти молекулы синтезируются в мембране эндоплазматического ретикулума (ЭПР) и затем распределяются по отдельным мембранам и органеллам внутри клетки. К их числу относится плазматическая мембрана, митохондрии и органеллы секреторного пути.
При возникновении необходимости клетка должна обладать способностью увеличивать количество мембран. Наиболее очевидно это проявляется при удвоении плазматической мембраны и всех органелл, которое происходит в ходе клеточного цикла. Наряду с этим, при гиперфункции, отдельные органеллы могут увеличиваться в размерах. Например, при созревании предшественников В-лимфоцитов в плазматические клетки, секретирующие антитела, ЭПР сильно увеличивается, что соответствует потребностям усиленного транспорта белков по секреторному пути.
Образование фосфолипидов из растворимых предшественников de novo происходит, главным образом, на мембране ЭПР со стороны цитоплазмы, в ходе процесса, называемого цикл Кеннеди. Как показано на при росте мембраны ЭПР две молекулы жирных кислот, связанные с ацетил КоА, реагируют с глицерин-3-фосфатом, образуя диацилглицерин (ДАТ). В отличие от своих предшественников, ДАГ достаточно гидрофобен для того, чтобы интегрироваться в мембрану ЭПР.
Читайте также: Функции лимфы как ткани
Попав в мембрану, ДАГ взаимодействует с «головой» будущей молекулы фосфолипида. Как показано на рисунке ниже, основные фосфолипиды различаются, главным образом, природой «головы» молекулы. Все они образуются при присоединении «головы» к ДАГ. Вначале происходит ее фосфорилирование, а затем присоединение к цитидиндифосфату (ЦДФ). После этого «голова» с одной из фосфатных групп переносится на ДАГ, и синтез завершается.
Присоединение головной группы холина к ЦЦФ с образованием фосфатидилхолина иллюстрирует использование клеткой субклеточной локализации белков для регуляции процессов биосинтеза. В этой реакции участвует фермент цитидилтрансфераза (ЦТ), и она контролирует скорость всего процесса синтеза фосфатидилхолина. В клетке ЦТ находится в двух различных формах: в неактивной, в цитозоле, и в активной, с цитозольной стороны мембраны ЭПР.
Механизм, который обеспечивает существование двух этих форм фермента, представляется не вполне ясным. Очевидно, однако, что условия внутри клетки, которые сигнализируют о необходимости усиления синтеза фосфатидилхолина, могут контролировать переход ЦТ из одной формы в другую. Например, увеличение в клетке концентрации свободного ДАТ или жирных кислот вызывает миграцию ЦТ на мембрану ЭПР, где эти компоненты реагируют с холином. В то же время, когда в клетке содержится много фосфатидилхолина, ЦТ локализуется в цитозоле. Таким образом, предотвращается синтез фосфатидилхолина.
Наряду с фосфатидилхолином, который является наиболее распространенным мембранным фосфолипидом, в цикле Кеннеди также образуются фосфатидилэтаноламин и фосфатидилинозитол. У дрожжей S. cerevisiae синтезируется фосфатидилсерин. У животных, однако, этот фосфолипид образуется за счет другой реакции, которая не требует участия ЦЦФ, однако также протекает с цитозольной стороны мембраны ЭПР.
Интересная особенность синтеза фосфатидилэтаноламина заключается в том, что хотя этот фосфолипид образуется в цикле Кеннеди, он также может синтезироваться в митохондриях. В этих органеллах он образуется при модификации фосфатидилсерина, который образовался в ЭПР. Образование фосфатидилэтаноламина в клеточных компартментах, отличающихся от тех, в которых синтезируется его предшественник, предполагает существование механизма транспорта липидов между двумя компартментами.
Имеются данные, позволяющие предполагать, что этот транспорт осуществляется посредством специального субдомена ЭПР, который называется мембраной, связанной с митохондрией (МСМ). МСМ представляет собой участок ЭПР, который физически ассоциирован с митохондрией. Он характеризуется плоской поверхностью и тесным контактом с органеллой. В МСМ находятся ферменты синтеза фосфатидилсерина. Возможно, что область контакта мембраны с митохондрией обеспечивает быстрый перенос фосфатидилсерина в органеллу, хотя механизм этого переноса остается неясным.
Наряду с участием в синтезе фосфолипидов, эндоплазматический ретикулум (ЭПР) является местом синтеза стеринов. К числу основных стеринов мембран клетки относится холестерин; его биосинтез осуществляется в несколько этапов. Первые несколько реакций происходят в цитозоле, а остальные в мембране ЭПР. Также в ЭПР находятся элементы, необходимые для регуляции образования холестерина. Ключевыми медиаторами этого регуляторного пути являются белки, связывающиеся с регуляторным стеринреспонсивным элементом (SREBP).
Читайте также: Чем можно убрать термоклей с ткани
Эти белки обычно интегрированы в мембрану эндоплазматического ретикулума (ЭПР) и со стороны люмена содержат небольшую петлю, соединяющую два трансмембранных домена, N- и С-концевые участки которых ориентированы в сторону цитозоля. Также в ЭПР находится белок SCAP (белок, активирующий разрезание SREBP). Этот белок контролирует уровень внутриклеточного холестерина. Когда возникает необходимость в повышенном синтезе холестерина, SCAP транспортирует SREBP из ЭПР в аппарат Гольджи. Там происходит разрезание SREBP, и N-терминальный участок молекулы высвобождается в цитозоль. (В реакции участвуют те же протеазы, которые расщепляют АТФ 6 при возникновении стрессорных условий в ЭПР. Затем N-терминальный домен SREBP мигрирует в ядро, где действует как фактор транскрипции, активируя экспрессию генов в каскаде реакций биосинтеза холестерина.
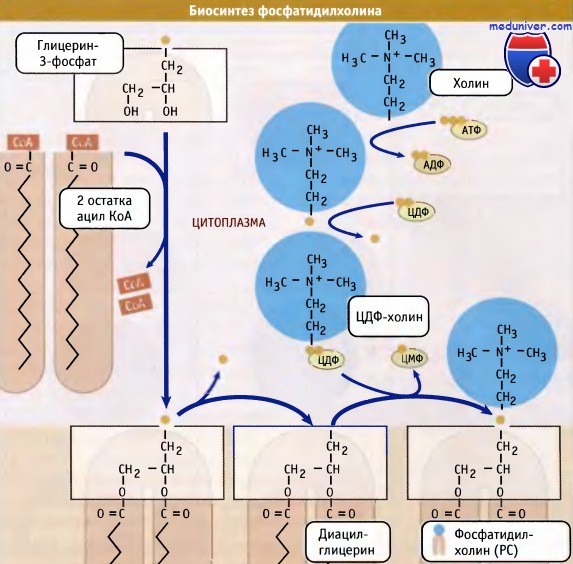 При объединении водорастворимых цитоплазматических компонентов (глицерин-3-фосфата, ацил КоА и полярной головки) образуется молекула фосфолипида.
При объединении водорастворимых цитоплазматических компонентов (глицерин-3-фосфата, ацил КоА и полярной головки) образуется молекула фосфолипида.
Новая растворимая в липидах молекула включается в мембрану. 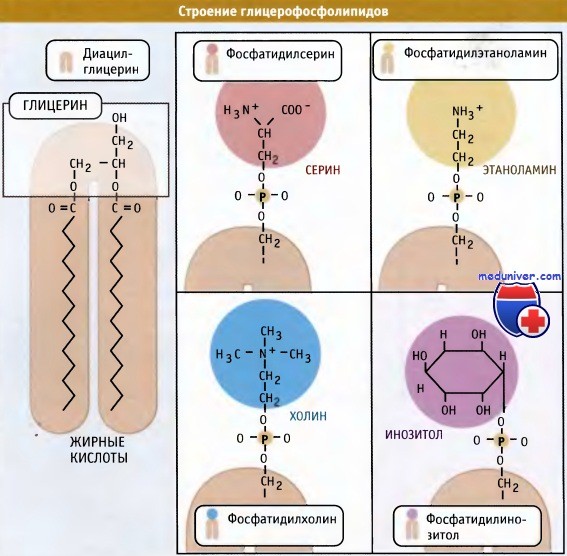 Основные мембранные фосфолипиды различаются по составу головок.
Основные мембранные фосфолипиды различаются по составу головок.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Синтез фосфолипидов
В отличие от триацилглицеролов и жирных кислот фосфолипиды не являются существенным энергетическим материалом. Фосфолипиды, как было сказано ранее, играют важную роль в структуре и функции клеточных мембран.
Биосинтез фосфолипидов интенсивно происходит в печени, стенке кишечника, молочной железе и других тканях. Наиболее важные фосфолипиды синтезируются главным образом в эндоплазматической сетке клетки.
Центральную роль в биосинтезе фосфолипидов играют L-диацилглицеролы (в синтезе фосфатидилхолинов и фосфатидилэтаноламинов), фосфатидная кислота (в синтезе фосфатидилинозитов) и сфингозин (в синтезе сфингомиелинов).
В этих биосинтезах принимает участие цитидинтрифосфат (ЦТФ), выполняющий роль активатора.
Основными компонентами мембран являются фосфатидилэтаноламин и фосфатидилхолин.
В процессе синтеза фосфатидилэтаноламина вначале происходит активация этаноамина под действием этаноламинкиназы:
Этаноламинфосфат реагирует затем с цитидинтрифосфатом (ЦТФ), в результате чего образуется цитидиндифосфатэтаноламин (ЦДФ-этаноламин) и пирофосфат (РРі). Эту реакцию катализирует фосфоэтаноламинцитидинтрансфераза:
Активированный ЦДФ–этаноламин при участии фермента этаноламинфосфаттрансферазы вступает в реакцию с L–диацилглицеролом с образованием фосфатидиламина:
L-диацилглицерол образуется при гидролизе фосфатидной кислоты. Аналогичные реакции ведут к образованию ЦДФ-холина.; последний вступает в реакцию с L-диацилглицеролом, в результате чего образуется фосфатидилхолин.
В синтезе фосфатидилэтаноламина и фосфатидилхолина важную роль играет ЦТФ. Подобно тому синтезу олиго- и полисахаридов, мы и здесь видим, каким образом нуклеотиды могут выполнять функцию переносчиков опреденных химических групп в обмене веществ клеток.
Помимо синтеза фосфолипидов таким способом, в клетке они легко подвергаются взаимопревращениям. Фосфатидилэтаноламин является предшественником фосфатидилхолина. Так, в печени в результате последовательного переноса трех метильных групп от трех молекул S-аденозилметионина образуется фосфатидилхолин:
В переносе метильных групп важную роль играют тетрогидрофолевая кислота (витамин Вс) и метилкобаломин ( витамин В12).
Синтез фосфатидилсерина происходит другим путем с предварительным вовлечением фосфатидной кислоты:
Затем происходит перенос серина на фосфатидильный остаток с образованием фосфатидилсерина:
У млекопитающих фосфатидилсерин образуется в реакции обмена этаноламина на серин следующим путем:
Таким же путем образуется фосфатидилинозитол.
В биосинтезе сфингомиелина принимает участие церамид (N-ацетилсфингозин), который образуется при взаимодействии спирта сфингозина и ацетил-КоА. Сфингомиелин синтезируется в результате взаимодействия церамида с ЦДФ-холином:
Читайте также: Брошь зонтик из ткани мастер класс
Все выше синтезированные фосфолипиды поступают с помощью липидпереносящих белков из цитоплазмы к мембранам и участвуют в их построении.
МЕТАБОЛИЗМ ФОСФОЛИПИДОВ
В отличие от триглицеридов и жирных кислот фосфолипиды не являются существенным энергетическим материалом. Фосфолипиды играют важную роль в структуре и функции клеточных мембран, активации мембранных и лизосомальных ферментов, в проведении нервных импульсов, свертывании крови, иммунологических реакциях, процессах клеточной пролиферации и регенерации тканей, в переносе электронов в цепи «дыхательных» ферментов. Особая роль фосфолипидам отводится в формировании липо-протеидных комплексов.
Биосинтез фосфолипидов интенсивно происходит в печени, стенке кишечника, семенниках, яичниках, молочной железе и других тканях. Наиболее важные фосфолипиды синтезируются главным образом в эндоплазма-тической сети клетки.
Центральную роль в биосинтезе фосфолипидов играют 1,2-диглицериды (в синтезе фосфатидилхолинов и фосфатидилэтаноламинов), фосфатидная кислота (в синтезе фосфатидилинозитов) и сфингозин (в синтезе сфин-гомиелинов). Цитидинтрифосфат (ЦТФ) участвует в синтезе практически всех фосфолипидов. В качестве примера рассмотрим синтез отдельных представителей фосфолипидов.
Биосинтез фосфатидилэтаноламина. Первоначально этаноламин при участии соответствующей киназы фосфорилируется с образованием фосфоэтаноламина:
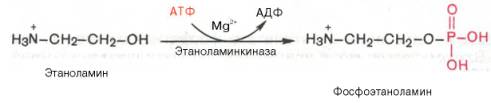
Затем фосфоэтаноламин взаимодействует с ЦТФ, в результате чего образуются цитидиндифосфатэтаноламин (ЦДФ-этаноламин) и пирофосфат (PPi):
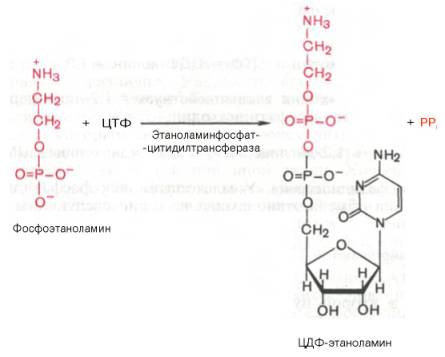
В следующей реакции ЦДФ-этаноламин, взаимодействуя с 1,2-дигли-церидом, образующимся при дефосфорилировании фосфатидной кислоты, превращается в фосфатидилэтаноламин. Реакция катализируется ферментом этаноламинфосфотрансферазой:
ЦДФ-этаноламин + 1,2-диглицерид –> Фосфатидилэтаноламин + ЦМФ.
Биосинтез фосфатидилхолина (лецитина). Фосфатидилэтаноламин является предшественником фосфатидилхолина. В результате последовательного переноса трех метальных групп от трех молекул S-аденозилметионина (донор метальных групп, см. главу 6) к аминогруппе остатка этаноламина образуется фосфатидилхолин:
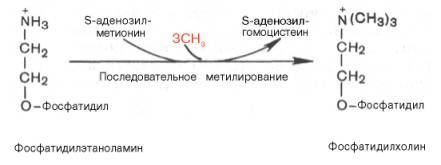
Существует еще один путь синтеза фосфатидилхолина в клетках животных. В этом случае, как и при синтезе фосфатидилэтаноламина, используется ЦТФ в качестве переносчика, но уже не фосфоэтаноламина, а фос-фохолина. На первом этапе синтеза свободный холин активируется под действием холинкиназы с образованием фосфохолина:
Затем фосфохолин реагирует с ЦТФ, образуя цитидиндифосфатхолин (ЦДФ-холин):
Фосфохолин + ЦТФ –> ЦДФ-холин + РРi.
В дальнейшем ЦДФ-холин взаимодействует с 1,2-диглицеридом, в результате чего образуется фосфатидилхолин:
ЦДФ-холин + 1,2-диглицерид –> Фосфатидилхолин + ЦМФ.
Биосинтез фосфатидилсерина. У млекопитающих фосфатидилсерин образуется в реакции обмена этаноламина на серин следующим путем:
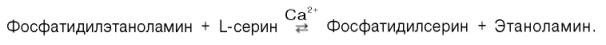
Существует и второй путь образования фосфатидилсерина, который связан с предварительным вовлечением фосфатидной кислоты в синтез фосфоглицеридов:
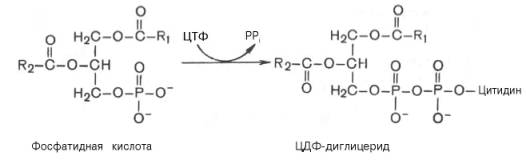
Затем происходит перенос серина на фосфатидильный остаток с образованием фосфатидилсерина:
ЦДФ-диглицерид + L-серин –> Фосфатидилсерин + ЦМФ.
Таким же путем образуется фосфатидилинозитол.
Биосинтез сфингомиелина. Интермедиатом в биосинтезе сфингомиелина является церамид (N-ацилсфингозин), который образуется при взаимодействии сфингозина с ацил-КоА. Сфингомиелин синтезируется в результате взаимодействия (реакции) церамида с ЦДФ-холином:

Следует отметить, что различие в синтезе холин- и этаноламинсодер-жащих фосфолипидов, с одной стороны, и инозитсодержащих фосфоли-пидов – с другой, заключается в том, что в первом случае при участии ЦТФ образуется ЦДФ-холин или ЦДФ-этаноламин – реакционноспособные азотистые основания, а во втором случае при участии ЦТФ образуется ЦДФ-диглицерид – реакционноспособная форма диглицерида.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом
- Правообладателям
- Политика конфиденциальности