

Механизм синтеза пиримидиновых нуклеотидов почти полностью расшифрован благодаря исследованиям Рейхарда. Показано, что в клетках животных и микроорганизмов промежуточными продуктами синтеза также не являются свободные пиримидиновые основания и что остаток рибозы присоединяется к уже сформировавшемуся пиримидиновому кольцу. Хотя синтез пиримидиновых нуклеотидов начинается с элементарных уровней (СО2, NH3, аспартата и др.), специфическую роль в этом синтезе играет оротовая кислота.
Оказалось, что оротовая кислота, впервые открытая в коровьем молоке, выполняет ключевую роль в синтезе, являясь непосредственным предшественником всех пиримидиновых нуклеотидов. Меченная 14 С и 15 N оротовая кислота открывалась в пиримидиновых основаниях ДНК и РНК тканей животных в опытах in vitro и in vivo. Экспериментами показaно образование оротовой кислоты в срезах печени крыс.
Последовательность химических реакций синтеза пиримидиновых нуклеотидов, в частности уридиловой кислоты, можно представить следующими уравнениями реакций:

Как видно из этих уравнений, первая стадия синтеза уридиловой кислоты включает образование карбамоилфосфата, механизм которого рассмотрен выше (см. Биосинтез мочевины). В клетках печени донатором NH2-группы являются глутамин и свободный аммиак (NH3), в то время как во всех остальных клетках организма в синтезе карбамоилфосфата используется только глутамин. Укажем также, что карбамоилфосфатсинтетаза требует для проявления своей активности присутствия N-ацетилглутамата в качестве аллостерического эффектора.
Во второй стадии карбамоилфосфат обратимо реагирует с аспарагиновой кислотой, причем равновесие реакции смещено в сторону синтеза карбамоиласпартата. Последний подвергается циклизации (под действием дигидрооротазы) с отщеплением молекулы Н20 и образуется дигидрооротовая кислота, которая, подвергаясь дегидрированию, превращается в оротовую. В этой реакции участвует специфический НАД-содержащий фермент дигидрооротатдегидрогеназа. Оротовая кислота обратимо реагирует с ФРПФ, являющимся донатором рибозо-5-фосфата, с образованием оротидин-5-фосфата (ОМФ). Декарбоксилирование последнего приводит к образованию первого пиримидинового нуклеотида — уридин-5-фосфата (УМФ), т. е. уридиловой кислоты.
Превращение УМФ в УДФ и УТФ осуществляется, как и в случае пуриновых нуклеотидов, путем фосфорилирования:
УМФ + АТФ УДФ + АДФ
УДФ + АТФ УТФ + АДФ
Биосинтез цитидиловых нуклеотидов. Имеющиеся данные свидетельствуют о том, что предшественником цитидиловых нуклеотидов является уридиловая кислота, точнее УТФ, который превращается в ЦТФ. Из последнего образуются другие цитидиловые нуклеотиды:

У прокариотов используется преимущественно свободный аммиак, в то время как в клетках животных ЦТФ-синтетаза катализирует включение амидной группы глутамина в 4-м положении пиримидинового кольца УТФ. Синтез ЦДФ и ЦМФ осуществляется при участии адениловых нуклеотидов путем фосфорилирования.
Биосинтез тимидиловых нуклеотидов. Поскольку тимидиловые нуклеотиды входят в состав ДНК, содержащей дезоксирибозу, рассмотрим сначала механизмы синтеза дезоксирибонуклеотидов. При помощи метода меченых атомов было показано, что этот синтез начинается не со свободной дезоксирибозы, а путем прямого восстановления рибонуклеотидов у 2-го атома углерода. При инкубации меченых предшественников (рибонуклеотидов) в бесклеточной системе бактерий метку обнаруживали в составе дезоксирибонуклеотидов. По данным Рейхарда, у Е. coli все четыре рибонуклеозиддифосфата восстанавливаются в соответствующие дезоксианалоги: dАДФ, dГДФ, dЦДФ, dУДФ при участии сложной ферментной системы, состоящей по меньшей мере из четырех разных ферментов.
Читайте также: Из каких тканей состоит зародыш семени
Химический смысл превращения рибонуклеотидов в дезоксирибонуклеотиды сводится к элементарному акту — восстановлению рибозы в 2-дезоксирибозу, требующего наличия двух атомов водорода; непосредственным источником водородных ионов оказался восстановленный термостабильный белок — тиоредоксин (содержит две свободные SH-группы на 108 аминокислотных остатков). Тиоредоксин легко окисляется, превращаясь в дисульфидную S-S-форму; для его восстановления в системе имеется специфический ФАД-содержащий фермент тиоредоксин-редуктаза (молекулярная масса 68 000), требующая наличия восстановленного НАДФН2. Обозначив условно рибонуклеозиддифосфат символом ХДФ, образование дезоксирибонуклеотидов можно представить следующими двумя уравнениями реакции:

Первая реакция требует затраты энергии. Схематически обе стадии могут быть представлены в следующем виде:

Для синтеза тимидиловых нуклеотидов, помимо дезоксирибозы, требуется также метилированное производное урацила — тимин. Оказалось, что в клетках имеется особый фермент тимидилатсинтаза, катализирующая метилирование не свободного урацила, а dУМФ; реакция протекает по уравнению:

Видно, что донатором метильной группы в тимидилатсинтазной реакции является N5, N10-метилентетрагидрофолиевая кислота, которая одновременно отдает и водородный протон, поэтому конечным продуктом кофермента является не тетрагидро-, а дигидрофолиевая кислота (ДГФ). Из образовавшегося ТМФ путем фосфотрансферазных реакций образуются dTДФ и dТТФ.
Синтез всех остальных дезоксирибонуклеозид-5′-трифосфатов, непосредственно участвующих в синтезе ДНК (см. ниже), также осуществляется путем фосфорилирования дезоксирибонуклеозид-5′-дифосфатов в присутствии АТФ:
АТФ + dАДФ —> АДФ + dАТФ; АТФ + dЦДФ —> АДФ + dЦTФ;
АТФ + dГДФ —> АДФ + dГТФ; АТФ + dТДФ —> АДФ + dТТФ.
Таким образом, выше рассмотрены главные механизмы синтеза предшественников ДНК и РНК. Указано на существенность ряда ключевых субстратов (например, оротовой кислоты, аспартата, глутамина и др.) для действия специфических ферментов синтеза нуклеотидов. Ниже в двух схемах суммированы имеющиеся данные о взаимопревращениях пуриновых и пиримидиновых нуклеотидов, а также о связи их с синтезом нуклеиновых кислот. Как видно из представленных данных, в образовании пуриновых и пиримидиновых нуклеотидов специфическое участие принимает 5-фосфорибозил-I-пирофосфат, являющийся донатором фосфорибозильного остатка в биосинтезе как оротидиловой кислоты, так и инозиновой кислоты; последние являются ключевыми субстратами синтеза нуклеиновых кислот в клетках.
Зная природу непосредственных предшественников нуклеиновых кислот, можно перейти к рассмотрению современных представлений о механизме синтеза нуклеиновых кислот.
БИОХИМИЯ УЧЕБНИК ДЛЯ ВУЗОВ — Е. С. Северина — 2004
РАЗДЕЛ 10. ОБМЕН НУКЛЕОТИДОВ
V. Биосинтез пиримидиновых нуклеотидов
Фонд пиримидиновых нуклеотидов, подобно пуриновым нуклеотидам, в основном синтезируется из простых предшественников de novo, и только 10 — 20% от общего количества образуется по «запасным» путям из азотистых оснований или нуклеозидов.
Читайте также: Полиэстеровая ткань что это за материал
А. Образование пиримидиновых нуклеотидов de novo
В отличие от синтеза пуринов, где формирование гетероциклического основания осуществляется на остатке рибозо-5-фосфата, пиримидиновое кольцо синтезируется из простых предшественников: глутамина, СО2 и аспарагиновой кислоты и затем связывается с рибозо-5-фосфатом, полученным от ФРДФ.
Процесс протекает в цитозоле клеток. Синтез ключевого пиримидинового нуклеотида — УМФ идёт с участием 3 ферментов, 2 из которых полифункциональны.
У млекопитающих ключевой, регуляторной реакцией в синтезе пиримидиновых нуклеотидов является синтез карбамоилфосфата из глутамина, СО2 и АТФ, в реакции катализируемой карбамоилфосфатсинтетазой II (КФС II), которая протекает в цитозоле клеток (рис. 10-12). В реакции NH2-rpynna карбамоилфосфата образуется за счёт амидной группы глутамина, что отличает эту реакцию от реакции синтеза карбамоилфосфата в митохондриях в процессе синтеза мочевины из СО2, NH3и АТФ с участием КФС I.
Рис. 10-12. Синтез карбамоилфосфата.

Карбамоилфосфат, использующийся на образование пиримидиновых нуклеотидов, является продуктом полифункционального фермента, который наряду с активностью КФС II содержит каталитические центры аспартаттранскарбамоилазы и дигидрооротазы. Этот фермент назвали «КАД-фермент» — по начальным буквам ферментативных активностей, которыми обладают отдельные каталитические домены этого белка. Объединение первых трёх ферментов метаболического пути в единый полифункциональный комплекс позволяет использовать почти весь синтезированный в первой реакции карбамоилфосфат на взаимодействие с аспартатом и образование карбамоиласпартата, от которого отщепляется вода и образуется циклический продукт — дигидрооротат (рис. 10-13).
Рис. 10-13. Биосинтез УМФ de novo.

Отщепляясь от КАД-фермента, дигидрооротат подвергается дегидрированию NAD-зaвиcимой дигидрооротатдегидрогеназой и превращается в свободное пиримидиновое основание — оротовую кислоту, или оротат.
В цитозоле оротат становится субстратом бифункционального фермента — УМФ-синтазы, которая обнаруживает оротатфосфорибозилтрансферазную и ОМФ-декарбоксилазную активности. Первоначально фосфорибозильный остаток от ФРДФ переносится на оротат и образуется нуклеотид — оротидин-5′-монофосфат (ОМФ), декарбоксилирование которого даёт уридин-5-мо- нофосфат (УМФ).
Таким образом, шесть последовательных реакций синтеза пиримидиновых нуклеотидов осуществляются тремя ферментами, которые кодируются в геноме человека тремя различными структурными генами.
Биосинтез УДФ, УТФ и цитидиловых нуклеотидов
УМФ под действием специфических нуклеозидмонофосфат (НМФ) и нуклеозиддифосфат (НДФ) киназ превращается в УДФ и УТФ в результате переноса у-фосфатного остатка АТФ на соответствующий субстрат.
НМФ-киназа катализирует следующую реакцию:
ЦТФ синтетаза катализирует амидирование УТФ (рис. 10-14), осуществляя АТФ-зависимое замещение кетогруппы урацила на амидную группу глутамина с образованием цитидин-5′-трифосфата (ЦТФ).
Рис. 10-14. Синтез ЦТФ из УТФ.

Б. ‹Запасные› пути синтеза пиримидиновых нуклеотидов
Использование пиримидиновых оснований и нуклеозцдов в реакциях реутилизации препятствует катаболизму этих соединений до конечных продуктов с расщеплением пиримидинового кольца. В ресинтезе пиримидинов участвуют некоторые ферменты катаболизма нуклеотидов. Так, уридинфосфорилаза в обратимой реакции может рибозилировать урацил с образованием уридина.
Урацил + Рибозо-1-фосфат —> Уридин + Н3РО4.
Читайте также: Папа страус ткани кто производит
Превращение нуклеозидов в нуклеотиды катализирует уридин-цитидинкиназа.
Часть ЦМФ может превращаться в УМФ под действием цитидиндезаминазы и пополнять запасы уридиловых нуклеотидов.
В. Регуляция синтеза пиримидиновых нуклеотидов
Регуляторным ферментом в синтезе пиримидиновых нуклеотидов является полифункциональный КАД-фермент. УМФ и пуриновые нуклеотиды аллостерически ингибируют, а ФРДФ активирует его карбамоилсинтетазную активность, тогда как активность аспартаттранскарбамоилазного домена ингибирует ЦТФ, но активирует АТФ (рис. 10-15).
Рис. 10-15. Регуляция синтеза пиримидиновых нуклеотидов. КАД-фермент катализирует реакции 1,2, 3; дигидрооротатдегидрогеназа — реакцию 4; УМФ синтетаза — реакции 5 и 6; НМФ киназа — реакцию 7; НДФ киназа — реакцию 8; ЦТФ синтетаза — реацию 9.

Этот способ регуляции позволяет предотвратить избыточный синтез не только УМФ, но и всех других пиримидиновых нуклеотидов и обеспечить сбалансированное образование всех четырёх основных пуриновых и пиримидиновых нуклеотидов, необходимых для синтеза РНК.
Биологическая библиотека — материалы для студентов, учителей, учеников и их родителей.
Наш сайт не претендует на авторство размещенных материалов. Мы только конвертируем в удобный формат материалы, которые находятся в открытом доступе и присланные нашими посетителями.
Если вы являетесь обладателем авторского права на любой размещенный у нас материал и намерены удалить его или получить ссылки на место коммерческого размещения материалов, обратитесь для согласования к администратору сайта.
Разрешается копировать материалы с обязательной гипертекстовой ссылкой на сайт, будьте благодарными мы затратили много усилий чтобы привести информацию в удобный вид.
© 2018-2022 Все права на дизайн сайта принадлежат С.Є.А.
Синтез пиримидиновых нуклеотидов линейный
Синтез пиримидиновых оснований происходит во всех клетках организма. В реакциях синтеза участвует аспарагиновая кислота, глутамин, СО2, затрачивается 2 молекулы АТФ. В отличие от разветвленного синтеза пуринов этот синтез происходит линейно, т.е. пиримидиновые нуклеотиды образуются последовательно, друг за другом.
Условно можно выделить 3 общих этапа синтеза и реакции синтеза УТФ и ЦТФ:
1. Образование карбамоилфосфата
Образование карбамоилфосфата в отличие от синтеза мочевины происходит в цитозоле большинства клеток организма.
2. Образование пиримидинового кольца
Формирование пиримидинового кольца происходит после присоединения аспартата и реакций дегидратации и окисления. Первым пиримидиновым основанием является оротовая кислота .
3. Синтез оротидинмонофосфата и уридинмонофосфорной кислоты
В реакции с фосфорибозилдифосфатом (ФРДФ) к оротовой кислоте присоединяется рибозо-5-фосфат и образуется оротидилмонофосфат, при декарбоксилировании превращающийся в уридинмонофосфат (УМФ).
Источником фосфорибозилдифосфата является первая из двух реакций синтеза фосфорибозиламина при образовании пуринов.
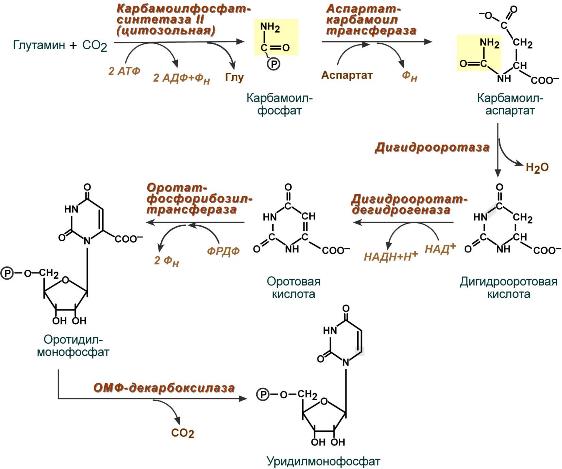
Синтез уридинмонофосфата
4. Синтез уридинтрифосфата
Синтез УТФ осуществляется из УМФ в 2 стадии посредством переноса макроэргических фосфатных групп от АТФ.
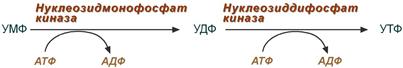
Синтез УТФ
5. Синтез цитидинтрифосфата
Образование цитидинтрифосфата (ЦТФ) происходит из УТФ с затратой энергии АТФ при участии глутамина, являющегося донором NH2-группы.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом
- Правообладателям
- Политика конфиденциальности
Мастерица © 2023
Информация, опубликованная на сайте, носит исключительно ознакомительный характер