
Мышечные ткани — это ткани, для которых способность к сокращению является главным свойством. Мышечные ткани составляют активную часть опорно-двигательного аппарата (пассивной частью являются кости, соединения костей).
Общими свойствами всех мышечных тканей является сократимость и возбудимость. К данной группе тканей относятся гладкая, поперечнополосатая скелетная и поперечнополосатая сердечная мышечные ткани. Клетки мышечной ткани имеют хорошо развитый цитоскелет, содержат много митохондрий.

Гладкая (висцеральная) мускулатура
Эта мышечная ткань встречается в стенках внутренних органах (бронхи, кишечник, желудок, мочевой пузырь), в стенках сосудов, протоках желез. Эволюционно является наиболее древним видом мускулатуры.
Состоит из веретенообразных миоцитов — коротких одноядерных клеток. Между клетками имеются межклеточные контакты — нексусы (лат. nexus — связь). Благодаря нексусам возбуждение, возникшее в одной клетке, волнообразно распространяется на все остальные клетки.

Гладкая мышечная ткань отличается своей способностью к длительному тоническому напряжению, что очень важно для работы внутренних органов (к примеру мочевого пузыря), сокращается медленно, практически не утомляется. Скелетная мышечная ткань, которую мы изучим чуть позже, такой способностью не обладает — сокращается и утомляется быстро.
Осуществляется сокращение с помощью клеточных органоидов — миофиламентов, которые расположены в клетке хаотично и не имеют такой упорядоченной структуры, как миофибриллы в скелетной мускулатуре (все познается в сравнении, уже скоро мы их тоже изучим).
Особо заметим, что в гладкой мышечной ткани миофиламенты собираются в миофибриллы только во время сокращения. У таких временных миофибрилл не может быть регулярной организации, а значит ни у таких миофибрилл, ни у гладких миоцитов не может быть поперечной исчерченности.
Гладкая мышечная ткань сокращается непроизвольно (неподвластна воле человека). Работа гладких мышц обеспечивается вегетативной (автономной) нервной системой. К примеру невозможно по желанию сузить или расширить бронхи, кровеносные сосуды, зрачок.

Гладкая мышечная ткань называется неисчерченной, так как не обладает поперечной исчерченностью, характерной для поперечнополосатых скелетной и сердечной мышечных тканей.
Скелетная (поперечнополосатая) мышечная ткань
Скелетная мышечная ткань образует диафрагму (дыхательную мышцу), мускулатуру туловища, конечностей, головы, голосовых связок.
В отличие от гладкой мускулатуры, скелетная образована не отдельными одноядерными клетками, а длинными многоядерными волокнами, имеющими до 100 и более ядер — миосимпластами. Миосимпласт (греч. sim — вместе + plast — образованный) представляет совокупность слившихся клеток, имеет длину от нескольких миллиметров до нескольких сантиметров (соответствует длине мышцы).
Внутри миосимпласта находится саркоплазма, снаружи миосимпласт покрыт сарколеммой. Сократительные элементы — миофибриллы (лат. fibra — волоконце) — длинные тяжеобразные органеллы в миосимпласте (около 1400).

Характерная черта данной ткани — поперечная исчерченность, выражающаяся в равномерном чередовании светлых и темных полос на мышечном волокне. Это происходит потому, что границы саркомеров в соседних миофибриллах совпадают, вследствие чего все волокно приобретает поперечную исчерченность. Теперь самое время изучить микроскопическую основу мышцы — саркомер.
Саркомер (от греч. sarco — мясо (мышца) + mere — маленький)
Саркомер — элементарная сократительная единица поперечнополосатых мышц, структурная единица миофибриллы. В состав саркомера (и миофибриллы в целом) входят миофиламенты (лат. filamentum — нить) двух типов, которые обеспечивают сократимость мышечной ткани.
Саркомер состоит из актиновых (тонких) и миозиновых (толстых) филаментов, которые образованы главным образом белками актином и миозином. Сокращение происходит за счет взаимного перемещения миофиламентов: они тянутся навстречу друг другу, саркомер укорачивается (и мышца в целом).

Источником энергии для сокращения служат молекулы АТФ. К тому же невозможно представить сокращение мышц без участия ионов кальция: именно они связываются с тропонином, что приводит к изменению конформации тропомиозина (тропонин и тропомиозин — регуляторные белки между нитями актина), за счет чего становится возможно соединение актина и миозина. При сокращении мышц выделяется тепло (сократительный термогенез).
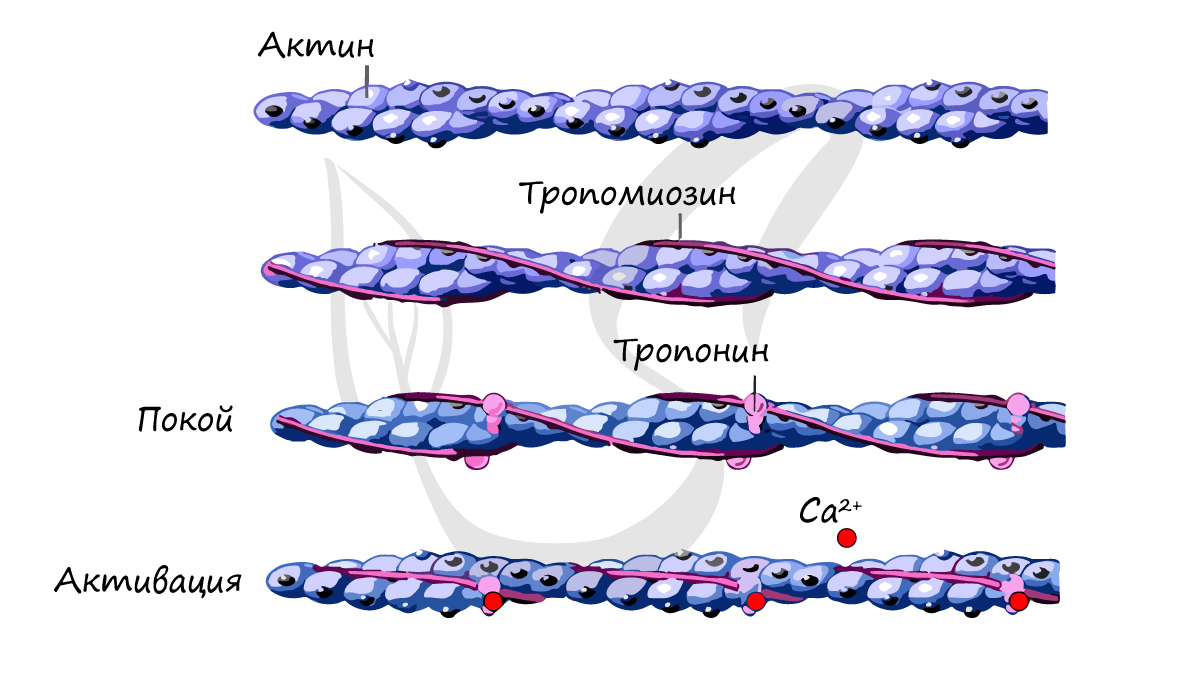
Замечу, что трупное окоченение (лат. rigor mortis) — посмертное затвердевание мышц — связано именно с ионами кальция, которые устремляются в область низкой концентрации (в саркоплазму миосимпласта), способствуя связыванию актина и миозина.
Читайте также: Из куска ткани сшили 24 детских платья
После смерти в мышце перестает синтезироваться АТФ, ее уровень быстро снижается. Как следствие этого перестает функционировать Ca-АТФаза — насос, выкачивающий ионы Ca из саркоплазмы в саркоплазматический ретикулум (мембранная органелла мышечных клеток (сходная с ЭПС), в которой запасаются ионы Ca).
В саркоплазме повышается концентрация ионов Ca — замыкаются мостики между актином и миозином, однако разомкнуться они уже не могут, в связи с чем наблюдается стойкая мышечная контрактура (лат. contractura — стягивание, сужение): конечности очень сложно разогнуть или согнуть.

Вернемся к скелетным мышцам. Имеется еще ряд важных моментов, о которых нужно знать.
В процесс возбуждения вовлекается изолированно один миосимпласт, соседние миосимпласты (волокна) не возбуждают друг друга, в отличие от гладких миоцитов, где возбуждение предается между соседними клетками через нексусы. Скелетные мышцы сокращаются быстро и быстро утомляются (у гладких мышц фазы сокращения и расслабления растянуты во времени, мало утомляются) .
Скелетные мышцы сокращаются произвольно: они подконтрольны нашему сознанию. К примеру, по желанию мы можем изменить скорость движения руки, темп бега, силу прыжка. Мышцы покрыты фасцией, крепятся к костям сухожилиями, и, сокращаясь, приводят в движение суставы.

Сердечная поперечнополосатая мышечная ткань
Сердечная мышечная ткань образует мышечную оболочку сердца — миокард (от др.-греч. μῦς «мышца» + καρδία — «сердце»). Миокард — средний слой сердца, составляющий основную часть его массы. При работе сердечная мышечная ткань не утомляется.

Сердечная мышечная ткань состоит из кардиомиоцитов — одиночных клеток, имеющих поперечную исчерченность. Соединяясь друг с другом, кардиомиоциты образуют функциональные волокна.
Этот тип мышечной ткани удивительным образом сочетает свойства двух предыдущих, изученных нами, тканей (возбудимость, сократимость) и имеет одно новое уникальное свойство — автоматизм.
Автоматизм — способность сердечной мышечной ткани возбуждаться и сокращаться самопроизвольно, без влияний извне. Это легко можно подтвердить, наблюдая сокращения изолированного сердца лягушки в физиологическом растворе: сокращения сердца в нем будут продолжаться несколько десятков минут после отделения сердца от организма.

Места контактов соседних кардиомиоцитов — вставочные диски (в их составе находятся нексусы), благодаря которым возбуждение одной клетки передается на соседние, таким образом волнообразно охватываются возбуждением и сокращаются новые участки миокарда.
Большое число контактов между кардиомиоцитами обеспечивает высокую эффективность и надежность проведения возбуждения по миокарду. Сокращается эта ткань непроизвольно, не утомляется.
На рисунке или микропрепарате узнать данную ткань можно по центральному положению ядер в клетках, поперечной исчерченности, наличию вставочных дисков и анастомозов (греч. anastomosis — отверстие) — мест соединений боковых поверхностей функциональных волокон (кардиомиоцитов).

В норме возбуждение проводится по проводящей системе сердца от предсердий к желудочкам (однонаправленно). Участок сердечной мышцы, в котором генерируются импульсы, определяющие частоту сердечных сокращений — водитель сердечного ритма.
Автоматизм возможен благодаря наличию в миокарде особых пейсмекерных (англ. pacemaker — задающий ритм) клеток, которые также называют водителями ритма. Они спонтанно генерируют нервные импульсы, которые охватывают весь миокард, в результате чего осуществляется сокращение. Именно благодаря водителям ритма сердце лягушки продолжает биться, будучи полностью отделенным от тела.
Ответ мышц на физическую нагрузку
Физические нагрузки приводят к гипертрофии мышц (от др.-греч. ὑπερ- чрез, слишком + τροφή — еда, пища) — в них увеличивается количество мышечных волокон, объем мышечной массы нарастает.

В условиях гиподинамии (от греч. ὑπό — под и δύνᾰμις — сила), то есть пониженной активности, мышцы уменьшаются вплоть до полной атрофии (греч. а – «не» + trophe – питание). В худшем случае волокна мышечной ткани перерождаются в соединительную ткань, после чего пациент становится обездвиженным.

Необходимо отметить, что сердечная мышечная ткань также дает ответную реакцию на чрезмерную нагрузку: сердце увеличивается в размере, нарастает масса миокарда. Причиной могут быть генетические заболевания, повышенное артериальное давление. Гипертрофия сердца — состояние, требующее вмешательства врача и наблюдения за пациентом.
Читайте также: Какие признаки характерны для тканей внутренней среды
В большинстве случае гипертрофия сердца обратима, а у спортсменов наблюдается так называемая физиологическая гипертрофия (вариант нормы).

Происхождение мышц
Мышцы развиваются из среднего зародышевого листка — мезодермы.

© Беллевич Юрий Сергеевич 2018-2022
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Бронхи какая мышечная ткань
В состав легких входят внутрилегочные бронхи разного калибра и респираторный отдел, представленный ацинусами. Система внутрилегочных бронхов начинается с долевых бронхов, на которые деляется главные (внелегочные) бронхи.
Внутрилегочные бронхи
Войдя в легочную паренхиму, долевые бронхи дихотомически делятся на бронхи крупного, среднего и мелкого калибра. Конечным звеном внутрилегочного бронхиального дерева являются терминальные бронхиолы.
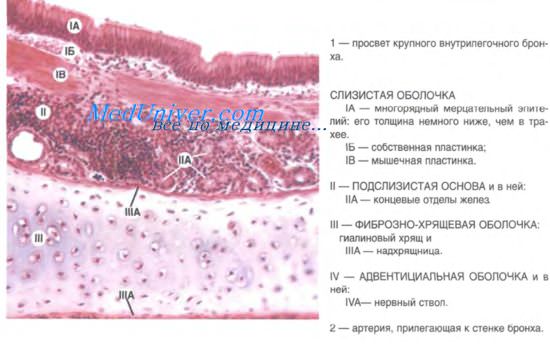
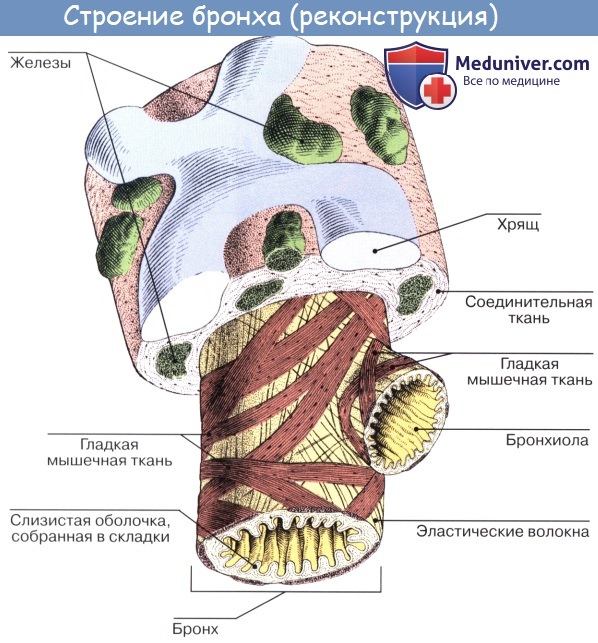
В главных бронхах стенка по строению сходна с трахеей. Волокнисто-хрящевая оболочка, однако, в отличие от трахеи состоит не из полуколец, а из замкнутых хрящевых колец.
Стенка внутрилегочных бронхов также состоит из слизистой оболочки с подслизистой основой, волокнисто-хрящевой и адвентициальной оболочек. Слизистая оболочка включает эпителий, собственную и мышечную пластинки. Последняя образована косоциркулярными пучками гладких миоцитов.
По мере дихотомического деления и уменьшения калибра внутрилегочных бронхов появляются существенные отличия в строении их оболочек. Так, многорядный мерцательный эпителий слизистой оболочки крупных и средних бронхов переходит в однослойный кубический — в мелких, количество бокаловидных клеток при этом уменьшается.
В подслизистой основе бронхов постепенно исчезают Многоклеточные слизисто-белковые железы. Изменения волокнисто-хрящевой оболочки сводятся к тому, что в ее составе по мере дихотомического деления бронхов уменьшается содержание хрящевой ткани, поскольку стенка средних и мелких бронхов, расположенных в глубине паренхимы легких, должна быть более податливой при расправлении и спадении альвеол.
Если в стенке крупных бронхов гиалиновая хрящевая ткань образует протяженные пластинки, соединенные друг с другом коллагеновыми волокнами и пучками гладких миоцитов, то в бронхах среднего калибра хрящ представлен мелкими островками и становится эластическим. Наконец, в мелких бронхах, лежащих наиболее глубоко, хрящевая ткань исчезает. Вместе с тем, по мере деления бронхов и уменьшения их калибра становится более выраженной циркулярная гладкомышечная пластинка, располагающаяся на границе слизистой оболочки и подслизистой основы.
В стенках мелких бронхов она занимает значительную часть. Гладкомышечная ткань стенки мелких бронхов высокочувствительна к концентрации СО2 увеличение его концентрации в воздухе вызывает расширение мелких бронхов. В связи с сокращением мышечных клеток слизистая оболочка мелких бронхов образует многочисленные складки. На всем протяжении бронхиального дерева в составе слизистой оболочки и подслизистой основы присутствуют скопления лимфоидной ткани, осуществляющей местные защитные реакции.
Бронхиальное дерево легких заканчивается терминальными бронхиолами. Их эпителий становится низким кубическим и, кроме реснитчатых, бокаловидных и базальных клеток, содержит секреторные клетки Клара, или бронхиолярные экзокриноциты. Последние в апикальной части накапливают специфические секреторные гранулы, являющиеся компонентом сурфактанта, а также содержат ферменты, инактивирующие токсины. Под базальной мембраной эпителия в тонкой собственной пластинке слизистой оболочки продольно идут пучки эластических волокон, между которыми располагаются гладкие мышечные клетки.
Физиологическая регенерация бронхиального эпителия происходит за счет митотического деления базальных клеток. Время обновления клеток эпителиального пласта составляет около недели.
Бронхи какая мышечная ткань
Строение легких. Разветвление бронхов. Макро-микроскопическое строение легкого.
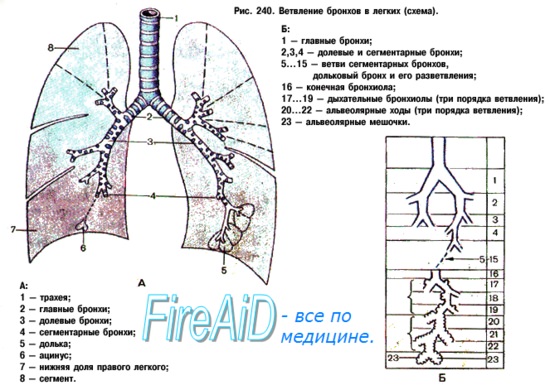
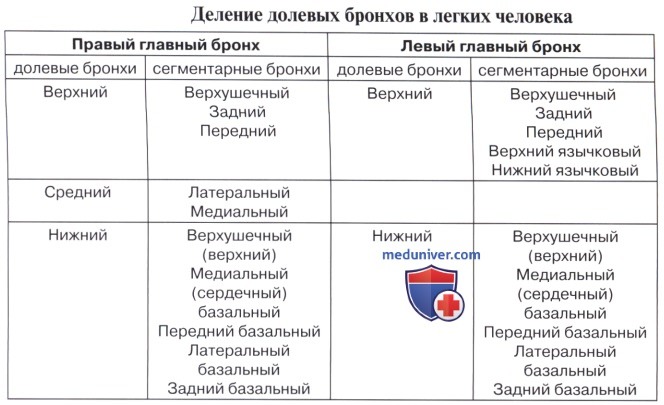
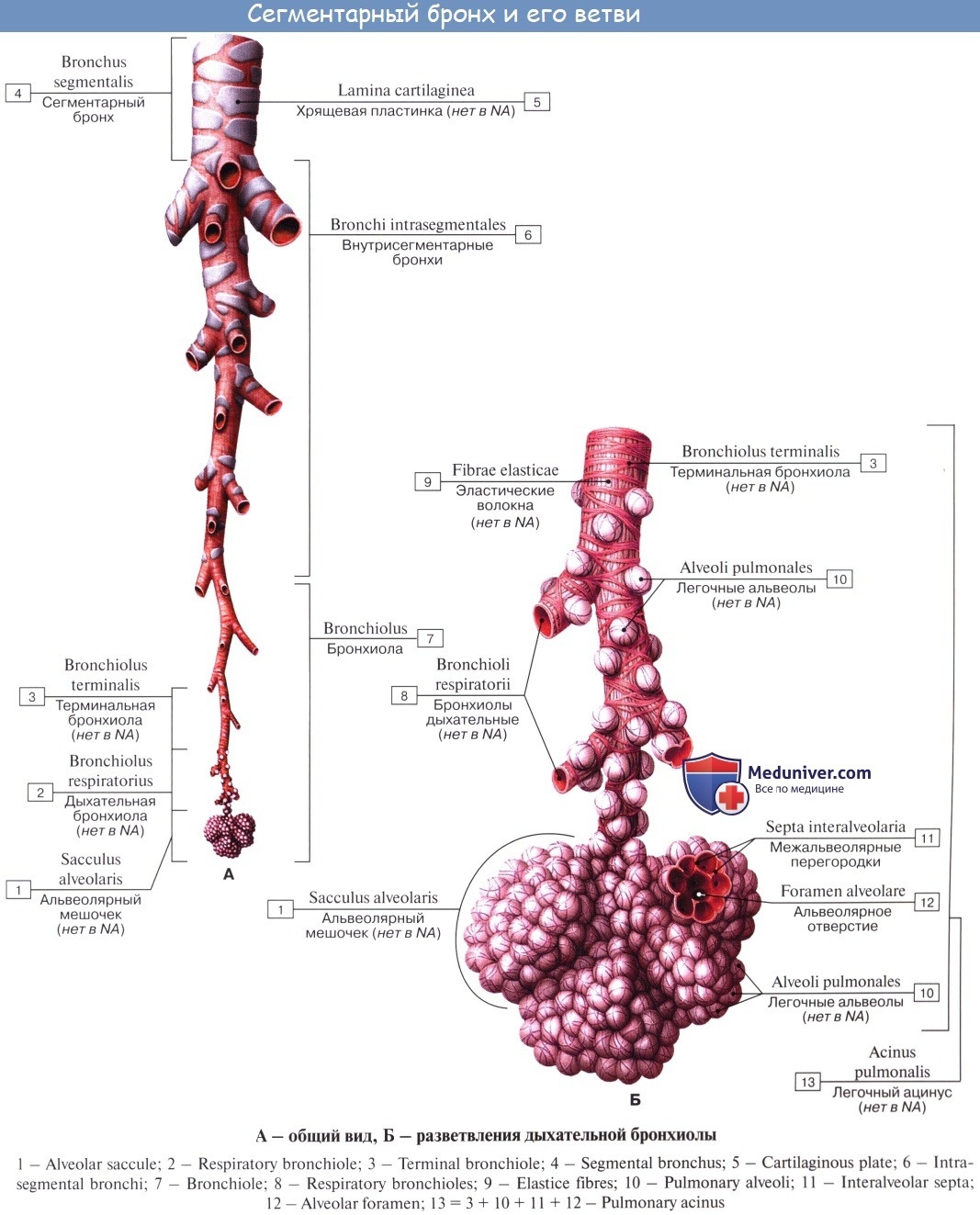
Соответственно делению легких на доли каждый из двух главных бронхов, bronchus principalis, подходя к воротам легкого, начинает делиться на долевые бронхи, bronchi lobares. Правый верхний долевой бронх, направляясь к центру верхней доли, проходит над легочной артерией и называется надартериальным; остальные долевые бронхи правого легкого и все долевые бронхи левого проходят под артерией и называются подартериальными. Долевые бронхи, вступая в вещество легкого, отдают от себя ряд более мелких, третичных, бронхов, называемых сегментарными, bronchi segmentates, так как они вентилируют определенные участки легкого — сегменты. Сегментарные бронхи в свою очередь делятся дихотомически (каждый на два) на более мелкие бронхи 4-го и последующих порядков вплоть до конечных и дыхательных бронхиол (см. ниже).
Скелет бронхов устроен по-разному вне и внутри легкого соответственно разным условиям механического воздействия на стенки бронхов вне и внутри органа: вне легкого скелет бронхов состоит из хрящевых полуколец, а при подходе к воротам легкого между хрящевыми полукольцами появляются хрящевые связи, вследствие чего структура их стенки становится решетчатой.
В сегментарных бронхах и их дальнейших разветвлениях хрящи не имеют более формы полуколец, а распадаются на отдельные пластинки, величина которых уменьшается по мере уменьшения калибра бронхов; в конечных бронхиолах хрящи исчезают. В них исчезают и слизистые железы, но реснитчатый эпителий остается.
Мышечный слой состоит из циркулярно расположенных кнутри от хрящей неисчерченных мышечных волокон. У мест деления бронхов располагаются особые циркулярные мышечные пучки, которые могут сузить или полностью закрыть вход в тот или иной бронх.
Макро-микроскопическое строение легкого
Сегменты легких состоят из вторичных долек, lobuli pulmonis secundarii, занимающих периферию сегмента слоем толщиной до 4 см. Вторичная долька представляет собой пирамидальной формы участок легочной паренхимы до 1 см в диаметре. Она отделена соединительнотканными перегородками от соседних вторичных долек.
Междольковая соединительная ткань содержит вены и сети лимфатических капилляров и способствует подвижности долек при дыхательных движениях легкого. Очень часто в ней откладывается вдыхаемая угольная пыль, вследствие чего границы долек становятся ясно заметными.
В верхушку каждой дольки входит один мелкий (1 мм в диаметре) бронх (в среднем 8-го порядка), содержащий еще в своих стенках хрящ (дольковый бронх). Число дольковых бронхов в каждом легком достигает 800. Каждый дольковый бронх разветвляется внутри дольки на 16—18 более т тонких (0,3 — 0,5 мм в диаметре) конечных бронхиол, bronchioli terminates, которые не содержат хряща и желез.
Все бронхи, начиная от главных и кончая конечными бронхиолами, составляют единое бронхиальное дерево, служащее для проведения струи воздуха при вдохе и выдохе; дыхательный газообмен между воздухом и кровью в них не происходит. Концевые бронхиолы, дихотомически ветвясь, дают начало нескольким порядкам дыхательных бронхиол, bronchioli respiratorii, отличающихся тем, что на их стенках появляются уже легочные пузырьки, или альвеолы, alveoli pulmonis. От каждой дыхательной бронхиолы радиарно отходят альвеолярные ходы, ductuli alveoldres, заканчивающиеся слепыми альвеолярными мешочками, sacculi alveoldres. Стенку каждого из них оплетает густая сеть кровеносных капилляров. Через стенку альвеол совершается газообмен.
Дыхательные бронхиолы, альвеолярные ходы и альвеолярные мешочки с альвеолами составляют единое альвеолярное дерево, или дыхательную паренхиму легкого. Перечисленные структуры, происходящие из одной конечной бронхиолы, образуют функционально-анатомическую единицу ее, называемую ацинус, acinus (гроздь).
Альвеолярные ходы и мешочки, относящиеся к одной дыхательной бронхиоле последнего порядка, составляют первичную дольку, lobulus pulmonis primarius. Их около 16 в ацинусе.
Число ацинусов в обоих легких достигает 30 000, а альвеол 300 — 350 млн. Площадь дыхательной поверхности легких колеблется от 35 м2 при выдохе до 100 м2 при глубоком вдохе. Из совокупности ацинусов слагаются дольки, из долек — сегменты, из сегментов — доли, а из долей — целое легкое.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом