
Непосредственно под мукозным эпителием слизистых оболочек в тесной связи с эпителиальными клетками располагаются лимфоциты пейеровых бляшек кишечника, лимфоидных фолликулов аппендикса, миндалин глотки, лимфоидных фолликулов подслизистого слоя верхних дыхательных путей и бронхов, мочеполового тракта. Все эти лимфоидные скопления получили собирательное название — мукозно-ассоциированная лимфоидная ткань.
В лимфоидных тканях, ассоциированных со слизистыми оболочками, проходят все стадии специфического иммунного ответа в тех случаях, когда антиген проникает в организм через слизистые оболочки. В мукозно-ассоциированных лимфоидных тканях активированные В-лимфоциты дифференцируются в плазматические клетки, продуцирующие специфические антитела, относящиеся к классу иммуноглобулинов A (IgA). Иммуноглобулины А, пройдя через эпителиальные клетки, где они присоединяют секреторный компонент, выходят на поверхность слизистых оболочек в форме секреторного иммуноглобулина A (SIgA), который обеспечивает местную антибактериальную и антивирусную защиту.
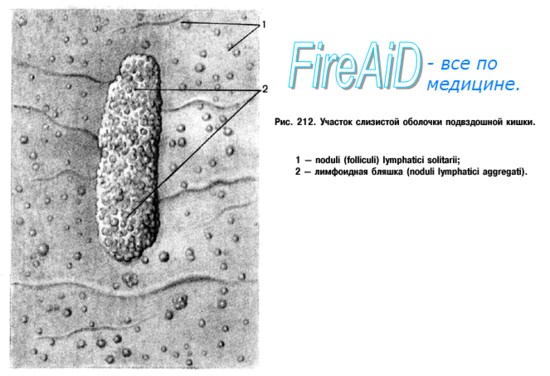
В качестве одного из периферических органов иммунной системы мукозно-ассоциированная лимфоидная ткань является местом:
1) распознавания антигена,
2) антигензависимой пролиферации и дифференцировки Т- и В-лимфоцитов,
3) активации Т- и В-лимфоцитов,
4) продукции цитокинов,
5) продукции и секреции специфических антител — иммуноглобулинов.
Защита легких [Иммунная система, Иммунитет]
Большую роль в развитии и течении заболеваний легких играют нарушения в системе местной защиты легких, предохраняющей респираторный тракт от воздействия патогенных факторов и обеспечивающей саногенез в бронхолегочной ткани.
Легкие в силу анатомо-топографических и функциональных особенностей подвергаются воздействию многочисленных неблагоприятных факторов окружающей среды. Ежедневно через легкие проходит около 10000 л воздуха, содержащего пыль, органические и неорганические частицы, микроорганизмы, резистентность к которым обусловлена эффективностью системы местной защиты.
Общность эмбриогенеза вилочковой железы с телебранхиальными тельцами, образующими так называемый бранхиальный комплекс, ставит бронхолегочную систему во взаимозависимость с главным органом иммунной системы — тимуса.
Сурфактант (сурфактантная система)
Одним из механизмов защиты легких является сурфактантная система, состоящая из фосфолипидов (фракции ненасыщенного фосфатидилхолина, лецитина), гликопротеидов, способствующих адгезии клеток к базальной мембране. Фосфолипиды сурфактанта обладают способностью активировать бактерицидность альвеолярных макрофагов, оказывать хемотаксическое действие. Антиоксидантная активность сурфактанта надежно защищает стенки альвеол от повреждающего действия окислителей и перекисей. Секреторная активность альвеолоцитов II типа (гранулярных пневмоцитов или сурфактантобластов) стимулируется простагландинами, ацетилхолином, препаратом лазолван, а цитостатики и β-адреноблокаторы угнетают секрецию сурфактанта.
Иммуноглобулины
К гуморальному звену местной зашиты относятся иммуноглобулины: IgA, IgG, IgM. Основная роль в защите трахеобронхиального дерева принадлежит IgA относительное содержание которого в бронхиальном секрете значительно выше, чем в сыворотке. Местный синтез IgA плазматическими клетками подслизистого слоя индуцируется воздействием антигенов. Важная роль в противомикробной защите более дистальных отделов принадлежит IgG. количество которого совпадает с его содержанием в сыворотке крови. Содержание IgG1 и IgG2 в бронхиальном секрете и сыворотке одинаково, а количество IgG3 и IgG4 в бронхиальном содержимом выше, чем в сыворотке, что, вероятно, свидетельствует об их местном синтезе.
Комплемент
Одним из факторов гуморального звена местной защиты бронхиального дерева является комплемент. При классическом (иммунном) пути активации комплемента принимают участие IgG, IgM, иммунные комплексы, С-реактивный белок. Важнейшим компонентом классического пути является С3-конвертаза, анафилатоксины С3а и С5а, обладающие способностью увеличивать сосудистую проницаемость, вызывать бронхоспазм. Активатором альтернативного (пропердинового) пути комплемента являются IgA, бактериальные липополисахариды, а также ферментативный комплекс С3-конвертазы классического пути. В защите легочной ткани от инфекций имеет значение СЗ, при дефиците которого наблюдаются частые инфекционные осложнения.
Лизоцим
Лизоцим — фермент, способный лизировать клеточную оболочку некоторых бактерий. Основными источниками лизоцима являются моноциты и нейтрофилы периферической крови. По сравнению с другими органами легкие наиболее богаты лизоцимом. У недоношенных детей секреция фермента в 2 раза ниже нормы. Дефицит лизоцима может быть одной из причин бактериальных инфекций органов дыхания При использовании препаратов, угнетающих фагоцитоз, снижается количество секреции лизоцима.
Читайте также: Коллекция тканей животного происхождения
Лактоферрин
Лактоферрин — один из наименее изученных факторов гуморального звена, представляющий собой связывающий железо гликопротеин. Лактоферрин продуцируется железистыми клетками слизистых оболочек. Одно из основных’ свойств лактоферрина — бактерицидная активность по отношению к различным микроорганизмам из-за блокады поступления к ним ионов железа. В присутствии секреторного IgA не происходит связывания лактоферрина с поверхностью микроорганизма, что свидетельствует о кооперативном взаимодействии факторов гуморального звена
Фибронектин
Фибронектин — гликопротеин, относящийся к опсонинам, основным источником его являются альвеолярные макрофаги.
Интерферон
Различают α-, β- и γ-ИФН, их свойства зависят от клеточного источника продукции. Лейкоцитарный интерферон — α-интерферон, фибробластный — β -интерферон, иммунный — γ-интерферон. Биологическое значение интерферона состоит в угнетении размножения вирусов, осуществлении противоопухолевого и противобактериального действия (преимущественно α — и β-ИФН), а также в иммуномодулирующем эффекте, усилении фагоцитоза (преимущественно γ -ИФН).
Ингибиторы протеаз
Ингибиторы протеаз — это факторы гуморального звена, защищающие слизистую от повреждающего действия протеолитических ферментов. Среди последних особенно активны эластаза, коллагеназа. Дефицит ингибиторов протеаз приводит к повреждению тканей легких протеолитическими ферментами. Система ингибиторов протеаз включает α1-антитрипсин и α2-макроглобулин. α1-Антитрипсин синтезируется в гепатоцитах и поступает в легкие из сыворотки крови. Это самый мощный фактор системы, так как на его долю приходится около 90 % общей ингибирующей активности сыворотки.
Альвеолярные макрофаги
Клеточное звено местной защиты — альвеолярные макрофаги (AM) — часть системы фагоцитирующих мононуклеаров организма. Появление альвеолярных макрофагов к концу беременности свидетельствует о зрелости местных защитных механизмов. Под влиянием антигистаминных препаратов и повышения уровня цАМФ угнетается подвижность и поглотительная активность фагоцитирующих клеток, а активизируются эти функции под влиянием повышения уровня цГМФ и гистамина. Материал с сайта http://wiki-med.com
Гранулоциты
К клеточному звену местной защиты бронхолегочной системы относятся также нейтрофильные и эозинофильные гранулоциты, которые составляют около 1 % всей клеточной популяции.
Бронхоассоциированная лимфоидная ткань (БАЛТ)
Бронхоассоциированная лимфоидная ткань — это скопление лимфоидных узелков и лимфоидных агрегатов в различных отделах органов дыхания. Она является резервуаром иммунокомпетентных клеток, проникающих в просвет воздухоносных путей. Около 70% таких клеток относятся к популяции Т-лимфоцитов и лишь 7 % — к В лимфоцитам. Большинство В-лимфоцитов несут поверхностные IgM и IgD. Особенностью В-лимфоцитов бронхассоциированной лимфоидной ткани является преимущественный синтез секреторного IgA — основного фактора гуморальною звена местной защиты легких.
Нарушение функции Т-лимфоцитов бронхоальвеолярной лимфоидной ткани может вести, с одной стороны, к недостаточной активации AM, а с другой — к нарушению кооперационного взаимодействия между Т- и В-лимфоцитами, способствующему снижению продукции антител.
Структура бронхо-ассоциированной лимфоидной ткани
Еще одной составляющей частью общей мукозальной иммунной системы является бронхо-ассоциированная лимфоидная ткань (BALT). Описывая в 1973 году структуру и функцию BALT авторы Bienenstock J. et al. (1973) дали классическое определение бронхо-ассоциированной лимфоидной ткани, как агрегированная лимфоидная структура, отделенная от бронхиального просвета специализированным лимфоэпителием. Существование BALT было описано у крыс, кроликов (Finke D. et al., 2001; Delventhal S. et al., 1992). Вместе с тем наличие BALT у мышей и человека продолжает оставаться дискуссионным вопросом. Основываясь на результатах, полученных при изучении BALT у кроликов и крыс, можно дать следующее описание этой части общей иммунной системы слизистых. Наиболее важной структурой является лимфоидный фолликул, образованный преимущественно В-лимфоцитами, где есть небольшое количество Т-лимфоцитов. Скопление лимфоцитов в лимфоидном фолликуле формирует классический герминальный центр. В парафолликулярной области, расположены высоко эндотелиальные венулы, через которые в область BALT поступают лимфоциты. Следует подчеркнуть, что также, как и для Пейеровых бляшек, высокоэндотелиальные венулы являются единственными воротами для миграции лимфоцитов из крови в бронхо-ассоциированную лимфоидную ткань (Otsuki Y. et al., 1989; Chamberlain D.W. et al., 1973). Фолликулы частично окружены лимфатическими сосудами, заполненными лимфоцитами. По этим сосудам лимфоциты покидают лимфоидные фолликулы и мигрируют в дренирующий бронхиальный лимфатический узел (Otsuki Y. et al., 1989). По направлению к бронхиальному эпителию в некоторых случаях удается видеть выпячивание в просвет бронха региона под названием dome (купол), описанного ранее для Пейеровых бляшек. Изолированные лимфоидные фолликулы лежат на базальной мембране под эпителием и покрыты сверху фолликуло-асоциированным эпителием (Bockman D.E. et al., (1973); Owen R.I. et al., (1977)). Эти фолликулы обычно имеют достаточно выраженное количество агрегированных В-клеток с небольшим количеством специализированных Т-лимфоцитов (Sminia T. et al., 1989). Более крупные агрегаты лимфоидных фолликулов, подобные описанным Пейеровым бляшкам, также обнаруживаются в верхних дыхательных путях и чаще всего в месте бифуркации бронхиального дерева. Такие агрегаты описаны у большого количества видов животных, включая грызунов, кур, кошек, кроликов, овец, свиней и морских свинок (Bienenstock J. et al., 1973; Bienenstock J. et al., 1976; Bienenstock J. et al., 1973; Bienenstock J. et al., 2005). В бронхиальной ткани также описано присутствие дискретных лимфоцитов у млекопитающих и птиц, которые состоят преимущественно из В-лимфоцитов, экспрессирующих IgA.
Читайте также: Как обрабатывают горловину трикотажной ткани
У некоторых животных среди эпителиальных клеток описано наличие М-клеток, характерных для эпителия Пейеровых бляшек. Как мы уже сказали, в области лимфоидного фолликула большинство клеток представляют собой В-лимфоциты, хотя здесь встречаются CD4+ Т-лимфоциты, а также макрофаги. Через высокоэндотелиальные венулы в парафолликулярную область из крови поступают СD4+-клетки, CD8+-клетки и В-лимфоциты, которые становятся клетками, заполняющими бронхо-ассоциированную лимфоидную ткань.
Важно подчеркнуть, что так же, как Пейеровы бляшки и назофарингеальная лимфоидная ткань, бронхиальная лимфоидная ткань названа индуктивной частью (зоной) BALT. Считают, что так же, как и в случае с Пейеровыми бляшками или с назофарингеальной лимфоидной тканью, в зоне BALT М-клетки поглощают антиген, далее они его презентируют дендритным клеткам. Дендритные клетки в свою очередь процессируют антиген и только после этого инициируют развитие специфической иммунной реакции, включая продукцию специфического IgA и созревание специфических CD8+ цитотоксических лимфоцитов (Sminia T. et al., 1989; Gebert A. et al., 1999). Коммиттированные к продукции IgA В–лимфоциты покидают индуктивную зону BALT и мигрируют в бронхиальный лимфоузел, после чего они мигрируют в эфферентную часть (зону) BALT, которой является lamina propria бронхиальной слизистой.
Вся эта лимфоидная ткань на территории легких вовлечена в иммунный ответ по отношению к ингаляционным антигенам. Как и во всем организме эти иммунные реакции предполагают включение Т- и В-клеточного ответа. По мнению McDermott M.R. et al. (1979) есть все доказательства того, что бронхо-ассоциированная лимфоидная ткань является частью общей мукозальной иммунной системы, и что лимфоциты, сенсибилизированные или примированные на территории бронхо-ассоциированной лимфоидной ткани, могут мигрировать на территории других частей этой общей иммунной системы слизистых.
На самом деле существование лимфоидных фолликулов в бронхиальной стенке предположительно было впервые описано Burdon-Sanderson J. в 1870 году. В 1875 году Klein E. предполагал, что такие фолликулы подобны тем лимфоидным фолликулам, которые обнаружены в других частях слизистых оболочек, включая миндалины и кишечник. Эпителий, который покрывает такие лимфоидные фолликулы, имеет редуцированное количество ворсинок, небольшое количество бокаловидных клеток и клеток, которые окрашиваются Альциан голубым. Лимфоциты часто окружены М-клетками, что приводит к формированию клеточных кластеров (Racz P. et al., 1977) с наличием клеток-«нянь»; при этом чаще всего описываются Т-лимфоциты без признаков апоптоза. Такие кластеры, впервые описанные Nieuwenhuis P. в 1971 году впоследствии подтверждены Wekerle H. и соавторами в 1980 году, были описаны на периферии BALT у крыс (Otsuki Y. et al., 1989). M-клетки происходят из базальных стволовых клеток респираторного эпителия и трансформируются в фенотипически отличающиеся клетки под прямым воздействием близлежащей лимфоидной ткани. У мышей М-клетки экспрессируют специфические молекулы на своей поверхности, что может быть селективно выявлено после связывания с улекс лектином (Clark M.A. et al., 1995); это же было описано в бронхо-ассоциированной лимфоидной ткани (Tango M. et al., 2000). М-клетки бронхо-ассоциированной лимфоидной ткани, также как и М-клетки кишечника, покрывающие Пейеровы бляшки, способны селективно фагоцитировать и пиноцитировать частицы и молекулы из своего непосредственного микроокружения (Racz P. et al., 1977). Предполагают, что М-клетки могут захватывать из бронхиального просвета молекулы и частицы, и далее создавать условия для их поглощения клетками в лимфоидном фолликуле (Pabst O. et al., 2005). Лимфоидные фолликулы бронхо-ассоциированной лимфоидной ткани не имеют афферентных лимфатических путей, в то же время эфферентные лимфатические пути дренируются региональными лимфатическими узлами. Это обстоятельство отличает фолликулы бронхо-ассоциированной лимфоидной ткани, так же, как и любой другой части мукозальной иммунной системы, от других вторичных лимфоидных тканей, которые располагаются вне участков слизистой оболочки. Лимфоциты, которые поступают в бронхо-ассоциированную лимфоидную ткань, используют для этой цели посткапиллярные высокоэндотелиальные венулы, или венулы с высоким эндотелием. Этот эндотелий посткапиллярных венул экспрессирует большой уровень сосудистых клеточных адгезионных молекул 1 (VCAM-1), которые не встречаются в других вторичных лимфоидных тканях (Xu B. et al., 2003). Показана также высокая экспрессия на эндотелии посткапиллярных высокоэндотелиальных венул бронхо-ассоциированной лимфоидной ткани α4β интегрина, который обуславливает селективную миграцию лимфоцитов на территорию бронхо-ассоциированной лимфоидной ткани. Например, антитела к таким структурам, блокируют миграцию Т-клеток памяти, но не наивных Т- и В-клеток в бронхо-ассоциированной лимфоидной ткани. Это еще раз подтверждает, что бронхо-ассоциированная лимфоидная ткань является уникальным мукозальным лимфоидным местом.
Читайте также: Удаление масляного пятна с ткани
Большая часть клеток на территории бронхо-ассоциированной лимфоидной ткани являются В-клетками и большинство из них экспрессирует поверхностные IgA и IgM (Otsuki Y. et al., 1989). Эти результаты получены на крысах, однако они очень подобны тем, которые были описаны для кроликов (Rudzik O. et al., 1975). Еще раз следует подчеркнуть, что BALT может селективно захватывать материал из просвета бронхов – в качестве такого материала могут быть бактерии или какие-либо другие частицы (Bienenstock J. et al., 1982; Tenner-Racz K. et al., 1979). Это осуществляют М-клетки за счет фагоцитоза и пиноцитоза (Dziedzic D. et al., 1990).
В парафолликулярной области бронхо-ассоциированной лимфоидной ткани расположена область, где в основном присутствуют Т-лимфоциты. Описано также присутствие дендритных клеток в бронхо-ассоциированной лимфоидной ткани.
- Правообладателям
- Политика конфиденциальности