
Клетки белой и бурой жировой ткани значительно отличаются друг от друга. Клетка бурой жировой ткани имеет по всей цитоплазме много мелких жировых капель, окружающих большое количество митохондрий, обильно снабжена окончаниями симпатических нервов и кровеносными сосудами. В эмбриогенезе она развивается из миобластов миотома сомитов, т.е. из предшественников скелетной мускулатуры, а не из предшественников соединительной ткани, подобно белым адипоцитам.
Бурая жировая ткань у младенцев составляет около 5% от массы их тела. Она расположена подкожно, между лопатками, в виде небольших отложений вокруг мышц и кровеносных сосудов шеи, в щеках (комочки Биша), а также в большом количестве – в подмышечных впадинах. Более глубокие отложения находятся в переднем средостении, вдоль позвоночника, вокруг почек, надпочечников, аорты, между лопатками, около почек и щитовидной железы. Между лопатками, на грудной клетке и на плечах у младенцев бурые адипоциты граничат с белыми, но нет четкой грани между клетками, среди бурых адипоцитов на «территории» бурой жировой ткани располагаются в некотором количестве белые адипоциты.
По мере взросления количество клеток бурой жировой ткани в указанных зонах снижается, но совсем не исчезает. По частоте встречаемости в организме одна клетка бурого жира, в среднем, приходится на 100-200 белых адипоцитов.
Функции
Функцией бурой жировой ткани является оперативное обеспечение организма тепловой энергией ( адаптивный термогенез ). Основной механизм термогенеза определяется тем, что в митохондриях клеток бурых адипоцитов процессы окисления и фосфорилирования не сопряжены (разобщены) и при окислении поступающих сюда жирных кислот происходит, в основном, выделение тепла, а не синтез АТФ.
Разобщение окисления и фосфорилирования обеспечивают специальные белки – термогенины (разобщающие белки, uncoupling proteins, UCP). Существуют несколько изоформ разобщающих белков: UCP-1 находится преимущественно в бурой жировой ткани, UCP-2 – в бурой и белой, UCP-3 – в cкелетных мышцах, UCP-4 и UCP-5 – преимущественно в мозге.
Адаптивный термогенез – это повышение теплопродукции в ответ:
- на изменение окружающей среды (снижение температуры) и на переохлаждение организма. При этом активируется симпатическая нервная система и стимулируется липолиз в бурых адипоцитах,
- на стимуляцию катехоламинами (через β3-адренорецепторы) – повышение транскрипции гена UCP-1, что приводит к увеличению энергозатрат,
- на избыток потребления пищи или изменение ее состава (поступление жиров),
- на секрецию лептина (усиление транскрипции гена UCP-1),
- на какие-то патологические воздействия.
У человека адаптивный термогенез ярко выражен у новорожденных, но при взрослении становится менее востребованным, заменяясь на иные способы сохранения тепла.
У взрослых отсутствие (резкое снижение) бурой жировой ткани встречается в 10% всех случаев ожирения.
Бурой жировой ткани много у животных, которые впадают в зимний сон и зимнюю спячку, температура тела у них поддерживается за счет адаптивного термогенеза.
Небольшие количества бурой жировой ткани у здоровых взрослых людей осуществляют постпрандиальный (возникающий после еды) термогенез , что в определенной степени препятствует отложению жира на фоне избытка пищи. При ожирении клетки бурой жировой ткани практически отсутствуют и, соответственно, процесс постпрандиального термогенеза не происходит.
Бежевая жировая ткань
Установлено, что помимо белой и бурой жировых тканей имеется промежуточная разновидность – бежевая жировая ткань. Эта ткань является промежуточной между белой и бурой жировыми тканями. Бежевые адипоциты первоначально имеют крайне низкое количество термогенина, но, как и классические бурые адипоциты, реагируют на определенную стимуляцию (понижение температуры, мышечная работа) началом высокой экспрессии термогенина и превращением в активные теплообразующие адипоциты.
При работе скелетных мышц в них повышается содержание особого белка — фактора транскрипции PGC-1a (peroxisome proliferator-activated receptor Gamma Coactivator 1-alpha), этот белок также экспрессируется в бурой жировой ткани , сердце и почках. Он стимулирует увеличение количества другого белка FNDC5 (fibronectin type III domain-containing protein 5).
У белка FNDC5 имеется две функции:
- Даже в очень низких концентрациях он увеличивает синтез мРНК белка термогенина в 70-100 раз.
- При его расщеплении образуется секретируемый мышцами полипептид, который был назван ирисином . Этот гормон является посредником между физическими нагрузками и различными тканями организма, в том числе белой и бурой жировыми тканями.
Вырабатываясь в работающих мышцах, ирисин транспортируется по крови в белые адипоциты, меняет акценты транскрипции и преобразует белую жировую ткань в бежевую.
Ирисин вырабатывается мышечной тканью не только при физической работе, но и при действии на организм холода. Так, при дрожании от холода в течение 10-15 минут вырабатывается такое же количество ирисина, как и при часовой езде на велосипеде со средней интенсивностью.
Название ирисин дано по имени древнегреческой богини Ириды, которая являлась богиней радуги и вестницей богов Зевса и Геры.
Интересно, что белок FNDC5 также стимулирует синтез нейротрофического фактора BDNF (Brain-Derived Neurotrophic Factor) в зубчатой извилине гиппокампа головного мозга, т.е. в той зоне которая участвует в процессах обучения и памяти. BDNF, в свою очередь, способствует образованию новых нейронов из стволовых клеток и новых синапсов, которые закрепляют новые знания и консолидируют память.
Получены данные взаимосвязи мозговой и физической активности, на основании которых предполагается, что посильные упражнения на выносливость для пожилых людей помогут им сохранить умственное здоровье.
Липолиз триглицеридов в белой и бурой жировой ткани
ОСОБЕННОСТИ МЕТАБОЛИЗМА БЕЛОЙ ЖИРОВОЙ ТКАНИ
Энергетический обмен низкий, преимущественно анаэробный, ткань потребляет мало кислорода. Энергия АТФ в основном тратится на транспорт жирных кислот через клеточные мембраны (с участием карнитина).
Белковый обмен низкий, белки синтезируются адипоцитами преимущественно для соб-ственных нужд. На экспорт в жировой ткани синтезируются лептин, белки острой фазы вос-паления (α1-кислый гликопротеин, гаптоглобин), компоненты системы комплимента (адипсин, комплемент С3, фактор В), интерлейкины.
Углеводный обмен. Невысокий, преобладает катаболизм. Углеводный обмен в жировой ткани тесно связан с липидным.
Жировая ткань стоит на 2 месте по обмену липидов после печени. Здесь происходят реак-ции липолиза и липогенеза.
Читайте также: Ткани их свойства способ ухода
Липогенез. В жировой ткани синтез липидов идет в абсорбтивный период по глицеро-фосфатному пути. Процесс стимулируется инсулином.
1. Под действием инсулина на рибосомах стимулируется синтез ЛПЛ.
2. ЛПЛ выходит из адипоцита и фиксируется на поверхности стенки капилляра с помощью гепарансульфата.
3. ЛПЛ гидролизует ТГ в составе липопротеинов
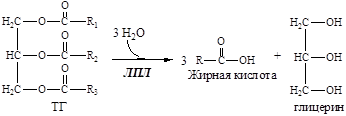
4. Образовавшийся глицерин уноситься кровью в печень.
5. Жирные кислоты из крови транспортируются в адипоцит.
6. Кроме поступающих из вне экзогенных жирных кислот, в адипоците синтезируются жир-ные кислоты из глюкозы. Процесс стимулируется инсулином.
7. Жирные кислоты в адипоците под действием Ацил-КоА синтетазы превращаются в Ацил-КоА.

7. Глюкоза поступает в адипоцит с участием ГЛЮТ-4 (активатор инсулин).
8. В адипоците глюкоза вступает в гликолиз с образованием ФДА (активатор инсулин).
9. В цитоплазме ФДА восстанавливается глицерол-ф ДГ до глицерофосфата:
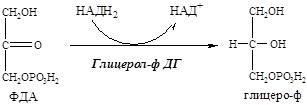
Так как в жировой ткани нет глицерокиназы, глицерофосфат образуется только из глюкозы (не может из глицерина).
10. В митохондриях глицерофосфат под действием глицеролфосфат ацилтрансферазы пре-вращается в лизофосфатид:
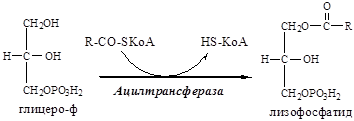
11. В митохондриях лизофосфатид под действием лизофосфатид ацилтрансферазы превраща-ется в фосфатид:

11. Фосфатид под действием фосфотидат фосфогидролазы превращается в 1,2-ДГ:
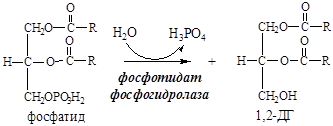
12. 1,2-ДГ под действием ацилтрансферазы превращается в ТГ:

13. Молекулы ТГ объединяются в крупные жировые капли.

2. Липолиз. Липолиз в жировой ткани активируется при дефиците глюкозы в крови (постаб-сорбционный период, голодание, физическая нагрузка). Процесс стимулируется глюкагоном, адреналином, в меньшей степени СТГ и глюкокортикоидами.
В результате липолиза концентрация свободных жирных кислот в крови возрастает в 2 раза.


ОСОБЕННОСТИ МЕТАБОЛИЗМА БУРОЙ ЖИРОВОЙ ТКАНИ
Энергетический обмен. Ткань потребляет много кислорода, активно окисляет глюкозу и жирные кислоты. Энергетический обмен высокий. При этом, АТФ образуется только в реак-циях субстратного фосфорилирования (2 реакции гликолиза, 1 реакции ЦТК). Причина — раз-общение в митохондриях белком термогенином (РБ-1) процессов окисления и фосфорилиро-вания, низкая активность АТФ синтетазы, отсутствие дыхательного контроля со стороны АДФ. В бурой жировой ткани вся образующаяся при окислении энергия рассеивается в виде тепла (термогенез).
Термогенез в бурой жировая ткань активируется при переохлаждении СНС, а также при излишке липидов в крови, под действием лептина. Благодаря этому повышается температура тела и снижается концентрация липидов в крови. Отсутствие бурой жировой ткани у взрос-лых людей является причиной 10% всех случаев ожирения.
42.Реакции β — окисления жирных кислот. Регуляция
β-окисление — специфический путь катаболизма ЖК с неразветвленной средней и корот-кой углеводородной цепью. β-окисление протекает в матриксе митохондрий, при котором от С конца ЖК последовательно отделяется по 2 атома С в виде Ацетил-КоА. β-окисление ЖК происходит только в аэробных условиях и является источником большого количества энер-гии.
β-окисление ЖК активно протекает в красных скелетных мышцах, сердечной мышце, почках и печени. ЖК не служат источником энергии для нервных тканей, так как ЖК не про-ходят через гематоэнцефалический барьер, как и другие гидрофобные вещества.
β-окисление ЖК увеличивается в постабсорбтивный период, при голодании и физической работе. При этом концентрация ЖК в крови увеличивается в результате мобилизации ЖК из жировых ткани.
Активация ЖК
Активация ЖК происходит в результате образования макроэргической связи между ЖК и HSКоА с образованием Ацил-КоА. Реакцию катализирует фермент Ацил-КоА синтетаза:
Пирофосфат гидролизуется ферментом пирофосфатазой: Н4Р2О7 + Н2О → 2Н3РО4
Ацил-КоА синтетазы находятся как в цитозоле (на внешней мембране митохондрий), так и в матриксе митохондрий. Эти ферменты отличаются по специфичности к ЖК с различной длиной углеводородной цепи.
Транспорт ЖК
Транспорт ЖК в матрикс митохондрий зависит от длины углеродной цепи.
ЖК с короткой и средней длиной цепи (от 4 до 12 атомов С) могут проникать в матрикс митохондрий путём диффузии. Активация этих ЖК происходит ацил-КоА синтетазами в матриксе митохондрий.
ЖК с длинной цепью, сначала активируются в цитозоле (ацил-КоА синтетазами на внешней мембране митохондрий), а затем переносятся в матрикс митохондрий специальной транспортной системой с помощью карнитина. Карнитин поступает с пищей или синтезируется из лизина и метионина с участием витамина С.
· В наружной мембране митохондрий фермент карнитинацилтрансфераза I (карнитин-пальмитоилтрансфераза I) катализирует перенос ацила с КоА на карнитин с образованием ацилкарнитина;
· Ацилкарнитин проходит через межмембранное пространство к наружной стороне внутренней мембраны и транспортируется с помощью карнитинацилкарнитинтранслоказы на внутреннюю поверхность внутренней мембраны митохондрий;
· Фермент карнитинацилтрансфераза II катализирует перенос ацила с карнитина на внутримитохондриальный HSКоА с образованием Ацил-КоА;
· Свободный карнитин возвращается на цитозольную сторону внутренней мембраны митохондрий той же транслоказой.
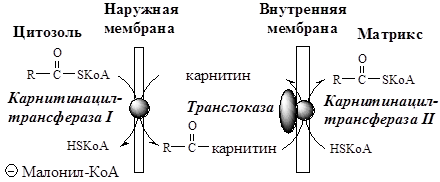
Реакции β-окисление ЖК
1. β-окисление начинается с дегидрирования ацил-КоА ФАД-зависимой Ацил-КоА дегидрогеназой с образованием двойной связи (транс) между α- и β-атомами С в Еноил-КоА. Восстановленный ФАДН2 окисляясь в ЦПЭ, обеспечивает синтез 2 молекул АТФ;
2. Еноил-КоА гидратаза присоединяет воду к двойной связи Еноил-КоА с образованием β-оксиацил-КоА;
3. β-оксиацил-КоА окисляется НАД зависимой дегидрогеназой до β-кетоацил-КоА. Восстановленный НАДН2, окисляясь в ЦПЭ, обеспечивает синтез 3 молекул АТФ;
4. Тиолаза с участием HКоА отщепляет от β-кетоацил-КоА Ацетил-КоА. В результате 4 реакций образуется Ацил-КоА, который короче предыдущего Ацил-КоА на 2 углерода. Образованный Ацетил-КоА окисляясь в ЦТК, обеспечивает синтез в ЦПЭ 12 молекул АТФ.
Затем Ацил-КоА снова вступает в реакции β-окисления. Циклы продолжаются до тех пор, пока Ацил-КоА не превратится в Ацетил-КоА с 2 атома С (если ЖК имела четное количество атомов С) или Бутирил-КоА с 3 атомами С (если ЖК имела нечетное количество атомов С).
Параграф 44 1 Липолиз
Автор текста – Анисимова Елена Сергеевна.
Авторские права защищены. Продавать текст нельзя.
Курсив не зубрить.
Замечания можно присылать по почте: exam_bch@mail.ru
https://vk.com/bch_5
Параграф 44:
Обмен жира.
См. также файл с таблицами и формулами.
Обмен жира – это процессы распада жира (липолиза) и синтеза жира.
См.также п.40, файл «44 формулы жир», п.21, 22.
Формула глицерина.
Формула жирной кислоты (см. п.40).
Формула жира.
Структурная формула жира (атомы водорода не структурно показаны).
Читайте также: Как покрасить ткань кофе в домашних условиях для тильды
44. 1. 1. Определение липолиза.
44. 1. 2. Где происходит липолиз? (локализация липолиза)
44. 1. 3. Значение липолиза.
44. 1. 4. Регуляция липолиза. (п.6-8, 91).
44. 1. 5. Реакции и ферменты липолиза.
44. 1. 7. Виды жировой ткани и их значение.
Три атома углерода,
при каждом есть ОН (гидроксильная группа).
Все три атома ОН групп глицерина (Н)
могут заменяться на остатки жирных кислот (ацилы),
связи с которыми называются сложноэфирными.
Формула глицерина:
Пока в отдельном файле.
Формула жирной кислоты (см. п.40):
Есть группа атомов СООН,
которая называется КАРБОКСИЛЬНОЙ ГРУППОЙ
Её структурная формула ОН – С = О,
а не С-О-О-Н,
С-О-О-Н – это пероксидная группа перекисей – п.27.
Вещества с карбоксильной СООН группой называются
КАРБОНОВЫМИ КИСЛОТАМИ (п.29).
Если в формуле карбоновой кислоты 4 и более атомов углерода,
то карбоновая кислота называется
высшей карбоновой кислотой или ЖИРНОЙ КИСЛОТОЙ.
Средняя длина углеродной цепочки – 16 атомов углерода (пальмитиновая кислота).
Формула пальмитата (типичной жирной кислоты):
СН3 – (СН2)14 – СООН.
Без ОН атомов карбоксильной группы
бывшая карбоновая кислота
называется остатком карбоновой кислоты
или (кратко) АЦИЛОМ:
Формулы кислот и ацилов:
пока в отдельном файле.
Ацилы могут присоединяться к атомам О групп ОН «вместо» атомов Н.
(Присоединяется атом С, с которым были соединены ОН группы).
Связи ацилов с атомами О называются сложноэфирными,
потому что соединение ацила и спирта называется сложным эфиром.
Если вместо атомов Н групп ОН глицерина (см. выше формулу)
присоединить атомы углерода ацилов
(те атомы С, которые были связаны с ОН атомами кислот),
то получается формула жира:
тут надо добавить таблицу
«формула жира»,
но пока она в отдельном файле.
Структурная формула жира (атомы водорода не структурно показаны).
Жир состоит из остатка глицерина и трёх остатков жирных кислот (ацилов), соединённых сложноэфирными связями. (Ацилы находятся «на месте» атомов водорода ОН групп глицерина).
Синоним жира – триацилглицериН (ТАГ) = триглицериД.
44. 1. Определение липолиза:
ЛИПОЛИЗ – это
превращение молекулы жира
в молекулу глицерина
и три молекулы жирных кислот:
НЭЖК – неэ(с)терифицированных жирных кислот, то есть не связанных сложноэфирными связями.
Можно считать липолиз процессом РАСПАДА ЖИРА.
При липолизе происходит расщепление сложноэфирных связей
между остатком глицерина и остатками жирных кислот (ацилами):
сначала расщепляются «крайние» (1 и 3) связи, а затем вторая.
При расщеплении этих связей «по месту разрыва связей» присоединяются молекулы воды,
то есть связи расщепляются путём гидролиза, гидролитически.
44. 2. Где происходит липолиз?
(локализация липолиза)
Липолиз протекает там, где есть жир.
Липолизу подвергается:
1) жир в белой и бурой жировой тканях,
2) пищевой жир в ЖКТ (см. п.43),
3) жир в составе ЛПОНП и хиломикронов в крови (см. п.49).
В белой и бурой жировой ткани жир расщепляется трилицерид/липазой
(первый, ключевой фермент, затем другие работают – см. далее),
пищевой жир в ЖКТ расщепляется панкреатической липазой, п.43
жир липопротеинов расщепляется липазой липопротеинов и печёночной липазой.
Далее обсуждается только липолиз в жировой ткани.
1. Благодаря липолизу человек может ХУДЕТЬ,
причём даже слишком сильно худеть:
до дефицита жировой ткани, который называется липодистрофией (см. п.91).
Липодистрофия бывает при избытке лептина, так как этот гормон усиливает липолиз.
Поэтому замедленный липолиз может стать причиной лишнего жира (ожирения),
а причиной замедления липолиза может быть дефицит гормонов
(или рецепторов этих гормонов), которые стимулируют липолиз:
— дефицит глюкагона (в сочетании с изобилием инсулина) из-за постоянной сытости,
— дефицит катехоламинов при гиподинамии,
— дефицит йодтиронинов при дефиците йода в пище и при болезнях щитовидной железы (п.104),
— дефицит АКТГ при недостаточности гипофиза и при избытке ГКС (п.108),
— дефицит лептина при мутациях генов, которые его кодируют
(тогда их называют «генами тучности»,
а до мутации они были генами стройности),
— дефицит СТГ при недосыпании (СТГ активно вырабатывается после 8 часов сна).
Худеть – хорошо,
слишком сильно худеть – плохо,
так как жировая ткань выполняет важные функции – п.40.
2. Но главное другое:
липолиз даёт организму жирные кислоты,
которыми питается большинство тканей организма,
в том числе такие важные, как сердце
(но не головной мозг и эритроциты:
эритроциты могут питаться только глюкозой – см. п. 121,
а головной мозг может питаться глюкозой
и кетоновыми телами,
но глюкозу при этом получать обязательно).
Питание тканей жирными кислотами позволяет тканям меньше использовать глюкозу,
оставляя её для головного мозга и эритроцитов.
Благодаря запасам жира и липолизу человек
может выжить при отсутствии пищи до двух месяцев (60 дней),
сохранять работоспособность в дни, когда не ел.
искать пищу или дорогу к людям.
При этом речь идёт о запасах жира нормальных стройных людей:
при весе около 50 кг на долю жира приходится около 10 кг.
Необязательно иметь лишний вес, чтобы прожить без пищи 2 месяца,
а если человек имеет 20 кг лишнего веса,
то это не означает, что он сможет прожить без пищи 4 месяца,
так как для этого нужен не жир, а белок
в качестве источника сырья для получения глюкозы: п.33.
Что означает выражение «ткани питаются жирными кислотами»:
это означает, что клетки тканей используют жирные кислоты
для выработки энергии в форме АТФ и тепла – п.22.
(АТФ используется для процессов в клетках и даёт ощущение сил,
а тепло поддерживает температуру в клетках на уровне 37 градусов Цельсия).
Для этого жирные кислоты должны подвергнуться бета-окислению – п.45.
3. Патологическое значение липолиза:
если липолиз протекает слишком активно,
то это приводит не только к дефициту жира,
но и к избытку НЭЖК в крови,
что далее приводит к образованию избытка кетоновых тел,
образующихся из НЭЖК в печени в процессе кетогенеза.
А избыток кетоновых тел приводит к КЕТОАЦИДОЗУ (п.47),
что может угрожать жизни (п.103),
может привести к сильной тошноте и сильной «неукротимой» рвоте («кетонемической»).
Читайте также: Что значит отделка ткани
Причинами слишком активного липолиза может быть:
дефицит гормона, тормозящего липолиз –
— инсулина при сахарном диабете (п.103) или
— избыток гормонов, стимулирующих липолиз:
— — избыток катехоламинов
— — — при стрессах,
— — — гиперфункции мозгового вещества коры надпочечников,
— — избыток глюкагона
— — — при голоде или
— — — гиперфункции альфа-клеток островков Лангерганса,
— — избыток АКТГ и СТГ
— — — при гиперфункции гипофиза,
— — избыток йодтиронинов
— — — при гиперфункции ЩВЖ (тиреотоксикозе),
— — избыток лептина.
44. 4. Регуляция липолиза. (п.6-8, 91).
Липолиз должен протекать в нужное время с нужной скоростью.
Иначе слишком активный липолиз приведёт к кетоацидозу,
а недостаточный липолиз приведёт к ожирению.
Например, липолиз нужен во время голода и физической работы.
И не нужен во время сытости и покоя.
Активность липолиза зависит от влияния на него гормонов,
которые влияют на скорость липолиза
через ферменты, катализирующие реакции липолиза:
чтобы изменить активность липолиза,
нужно изменить активность его ферментов.
П.6.
Активность липолиза снижается, когда он:
— когда он не нужен:
— — при сытости и покое,
— — под действием гормонона ИНСУЛИНА,
поэтому при дефиците инсулина липолиз протекает активнее, чем нужно,
что может привести к кетоацидозу (п.103).
И поэтому же при сытости и покое липолиз замедлен,
что мешает избавляться от лишнего веса,
а для избавления от него нужно:
— умеренное питание
(менее 2000 килокалорий в сутки, но не менее 1500)
— и физические нагрузки.
Активность липолиза повышается, когда он:
— когда он нужен:
— — то есть при голоде
— — и стрессах,
— — при работе
под действием гормонов:
— глюкагона (при голоде, это гормон голода),
— катехоламинов
(при стрессах и работе: это гормоны стресса),
а также:
— лептина («гормон стройности»),
— йодтиронинов,
— СТГ и АКТГ.
Дефицит этих гормонов
(из-за недосыпания – СТГ,
из-за дефицита йода в пище – ЙТ,
недостаточности желёз)
приводит к замедлению липолиза,
и препятствует избавлению от лишнего веса
(то есть ведёт к ожирению, может стать его причиной),
а избыток этих гормонов приводит к ускорению липолиза,
что помогает похудеть,
но создаёт риск кетоацидоза (тошноты, рвоты).
44. 5. Реакции и ферменты липолиза.
См. формулы в файле «44 ФОРМУЛЫ ЖИР ЛИПОЛИЗ».
В молекуле жира – три ацила,
присоединённых к остатку глицерина.
В первой реакции отщепляется первый ацил,
во второй – ещё один,
а в третьей – ещё один.
В итоге образуются три молекулы жирных кислот и молекула глицерина.
В первой реакции липолиза:
в молекуле жира расщепляется связь с первым ацилом,
в результате чего образуется одна молекула жирной кислоты (НЭЖК) и диглицериД (диацилглицериН).
Фермент, который катализирует эту реакцию, называется триглицерид/липазой, то есть липазой, субстратом которой является триглицерид.
НЭЖК выходит в кровь для транспорта в ткани и связывается там альбумином,
а диглицерид вступает в следующую реакцию.
Во второй реакции липолиза:
в молекуле диглицерида
расщепляется связь с третьим ацилом
(тем, который был третьим до отщепления первого),
в результате чего образуется одна молекула жирной кислоты (НЭЖК)
и моноглицериД (моноацилглицериН).
Фермент, который катализирует эту реакцию,
называется диглицерид/липазой,
то есть липазой, субстратом которой является диглицерид.
НЭЖК выходит в кровь для транспорта в ткани,
а моноглицерид вступает в следующую реакцию.
В третьей реакции липолиза:
в молекуле моноглицериДа расщепляется связь с последним ацилом,
в результате чего образуется одна молекула жирной кислоты (НЭЖК)
и молекула глицериНа.
Фермент, который катализирует эту реакцию,
называется моноглицерид/липазой,
то есть липазой, субстратом которой является моноглицерид.
НЭЖК выходит в кровь для транспорта в ткани, связывается в крови альбумином,
а глицерид транспортируется с током крови в печень,
где используется для синтеза жира или глюкозы (ГНГ – п.33).
Сказанное относится к липолизу в белой жировой ткани.
При липолизе в БУРОЙ жировой ткани образовавшиеся при липолизе
НЭЖК не выходят в кровь для питания тканей,
а подвергаются бету-окислению в клетках самой бурой жировой ткани.
Это приводит к выработке энергии
(с участием ЦТК и дыхательной цепи).
Причём часть энергии, которая рассеивается в виде тепла,
выше, чем в других клетках
за счёт РАЗОБЩЕНЯ (п.23),
а разобщителями являются сами жирные кислоты.
Это нужно для того, чтобы согревать кровь,
которая идёт к сердцу, мозгу и другим жизненно важным органам.
44. 7. Виды жировой ткани и их значение.
Есть БЕЛАЯ и бурая жировая ткань.
Жировая ткань – это не жир, не молекулы жира.
Жировые ткани состоят из клеток,
способных в больших количествах накапливать молекулы жира.
Различия в значении белой и бурой тканей:
Белая является «складом» жирных кислот
в виде ацилов в составе жира
(в этом её основное значение),
которые после липолиза поступают в разные ткани
для выработки энергии в тканях.
А бурая жировая ткань – это «грелка»,
которая согревает кровь, идущую к особо важным органам.
Согревает за счёт выработки в ней тепла:
1) выделяющегося при работе дыхательной цепи, потребляющей НАДН и ФАДН2,
2) вырабатывающиеся при работе ЦТК и бета-окислении жирных кислот,
3) образующихся при липолизе НЭЖК в клетках бурой жировой ткани.
Но обе ткани – склад жирных кислот в виде ацилов в молекулах жира.
Различия в локализации (расположении):
белая:
подкожная жировая клетчатка, сальники,
бурая:
между лопатками и т.д.
На долю белой может приходиться от 20% веса тела и больше,
а на долю бурой намного меньше (1%).
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом