
Метаболическая активность и термогенез в бурой жировой ткани – чувствительный маркер тиреоидного статуса
У пациентов с гипертиреозом наблюдалось повышение содержания бурой жировой ткани в надключичной области, а изменение уровня свободного трийодтиронина при раннем эутиреозе корректировало с метаболической активностью адипоцитов и скоростью обмена веществ.
Бурая жировая ткань (БЖТ) – один из двух типов жировой ткани млекопитающих, которая хорошо развита у новорожденных детей и животных, впадающих в спячку. Адипоциты БЖТ содержат много липидных капель (в отличие от одной капли в клетках белой жировой ткани), богаты митохондриями и кровеносными сосудами и иннервируются волокнами симпатической нервной системы. Морфологические характеристики ткани обусловлены способностью адипоцитов БЖТ к несократительному, т.е. не связанному с мышечной активностью термогенезу.
В процессе эмбрионального развития БЖТ появляется на 5-м месяце беременности, и у новорожденных детей ее масса составляет до 5% массы тела. У взрослых содержание бурого жира снижается, однако ткань остается метаболически активной и участвует в процессах энергетического обмена, окисления липидов и инсулинорезистентности. В экспериментальных исследованиях продемонстрирована чувствительность БЖТ к гормонам щитовидной железы, в частности трийодтиронину (Т3), выступающему в роли ключевого регулятора дифференцировки и метаболической активности адипоцитов.
На основании данных экспериментальных исследований, демонстрирующих прямую связь между активностью адипоцитов БЖТ и уровнем Т3 в модели гипер- или гипотиреоза, ученые из Института клинических наук Сингапура предположили, что уровень, метаболическая активность и термогенез в БЖТ у пациентов с заболеваниями щитовидной железы будет меняться в зависимости от статуса заболевания – гипертиреоза или эутиреоза.
Больным диффузным токсическим зобом (болезнь Грейвса; n=21, 5 мужчин; 39.5±2.5 лет) была назначена терапия антитиреоидными препаратами группы тионамидов (карбимазол, тиамазол). На фоне гипертиреоза и при диагностике раннего эутиреоза у пациентов измеряли клинические и биохимические маркеры и общие энергозатраты. Для оценки содержания БЖТ больным проводили инфракрасную термографию надключичной области. По данным ПЭТ/МРТ определяли средний и максимальный стандартизированный уровень захвата 18 F-флуородеоксиглюкозы (mean/maximum standardized uptake value, SUVmean/max) – индикатор метаболической активности адипоцитов, фракцию бурого жира в надключичной ямке (ФБЖН) и локальное повышение температуры в ткани как маркер увеличения ее метаболической активности (mean temperature, Tscv).
Среднее время перехода от гипертиреоза к эутиреозу на фоне антитиреоидной терапии составило 28.6±2.3 недель.
По данным ПЭТ/МРТ наличие БЖТ было выявлено у 8 пациентов с гипертиреозом (ФЖН от 72.3±1.4% до 76.8±1.4%; P 18 F-флуородеоксиглюкозы клетками БЖТ (r=0.58, P=0.01) и скоростью обмена веществ в покое (P
Особенности метаболизма бурой жировой ткани
Энергетический обмен. Ткань потребляет много кислорода, активно окисляет глюкозу и жирные кислоты. При этом энергетический обмен низкий. АТФ образуется только в реакциях субстратного фосфорилирования (2 реакции гликолиза, 1 реакции ЦТК). Причина — разобщение в митохондриях белком термогенином (РБ-1) процессов окисления и фосфорилирования, низкая активность АТФ синтетазы, отсутствие дыхательного контроля со стороны АДФ. В бурой жировой ткани вся образующаяся при окислении энергия рассеивается в виде тепла (термогенез).
Термогенез в бурой жировая ткань активируется при переохлаждении СНС, а также при излишке липидов в крови, под действием лептина. Благодаря этому повышается температура тела и снижается концентрация липидов в крови. Отсутствие бурой жировой ткани у взрослых людей является причиной 10% всех случаев ожирения.
1. Исследовать содержание перекисных радикалов и гидроперекисей липидов в сыворотке кр
2. Исследовать содержание МДА в сыворотке крови больного и здорового человека.
В заключениесвоей работы студенты должны выбрать правильный диагноз (гиперхиломикронемия) из приведенного перечня вероятных ответов, исходя из анализа собственных исследованиий сыворотки больного и приведенных в методическом указании лабораторных исследований, физикального обследования и жалоб больного. Затем
кратко записывают цепочку событий с участием ПОЛ в развитии панкреонекроза: гипер–ХМ-мияэмболиянарушение кровотокагипоксияактивац.ПОЛповреждение мембран
активация фагоцитозаотложение пенистых клеток и гипертрофия органа панкреатит
9. ЭТАЛОНЫ ОТВЕТОВ НА ЗАДАЧИ, тестовые вопросы, используемые при контроле знаний на занятии (можно в виде приложения)
10. ХАРАКТЕР И ОБЪЕМ ВОЗМОЖНОЙ УЧЕБНО-ИССЛЕДОВАТЕЛЬСКОЙ РАБОТЫ ПО ТЕМЕ
(Указать конкретно характер и форму УИРС: подготовка реферативных выступлений, проведение самостоятельных исследований, имитационная игра, оформление истории болезни с использованием монографической литературы и др. формы)
11. ЛИТЕРАТУРА ДЛЯ ПОДГОТОВКИ К ЗАНЯТИЮ ПРЕПОДАВАТЕЛЯМ:
Основная
Березов Т.Т., Коровкин Б.Ф. Биологическая химия. – 3-е изд., перераб. и доп. –М.: Медицина. – 2004. – 704с.
Николаев А.Я. Биологическая химия. – 3-е изд., перераб. и доп. –М.: Медицинское информационное агентство. – 2004. – 566с.: ил.
Биохимия: Учебник / Под ред. Е.С. Северина. – М.: ГЭОТАР-МЕД. 2005. – 784 с.: ил. – (Серия «XXIвек»)
Дополнительная
Климов А.Н., Никуличева Н.Г. Липиды, липопротеиды, атеросклероз. — СПб. «Питер». – 1995. – С. 33-47.
Балаболкин М.И. Диабетология. – М.: Медицина. – 2000. – С. 143-148.
Марри Р., Греннер Д., Мейес П., Родуэлл В. Биохимия человека: в 2-х томах. Т. (?). Пер. с англ.: — М.: Мир, – 1993. – 384 С. С: 225-231.
12. Методическая разработка подготовленадоц. Трубачевым С.Д.
обсужденана заседании кафедры 3 сентября 2007
За сжигание жира тоже отвечает жировая ткань
Бурая жировая ткань
Строение
Клетки белой и бурой жировой ткани значительно отличаются друг от друга. Клетка бурой жировой ткани имеет по всей цитоплазме много мелких жировых капель, окружающих большое количество митохондрий, обильно снабжена окончаниями симпатических нервов и кровеносными сосудами. В эмбриогенезе она развивается из миобластов миотома сомитов, т.е. из предшественников скелетной мускулатуры, а не из предшественников соединительной ткани, подобно белым адипоцитам.
Читайте также: Ткань для медицинских масок как называется
Бурая жировая ткань у младенцев составляет около 5% от массы их тела. Она расположена подкожно, между лопатками, в виде небольших отложений вокруг мышц и кровеносных сосудов шеи, в щеках (комочки Биша), а также в большом количестве – в подмышечных впадинах. Более глубокие отложения находятся в переднем средостении, вдоль позвоночника, вокруг почек, надпочечников, аорты, между лопатками, около почек и щитовидной железы. Между лопатками, на грудной клетке и на плечах у младенцев бурые адипоциты граничат с белыми, но нет четкой грани между клетками, среди бурых адипоцитов на «территории» бурой жировой ткани располагаются в некотором количестве белые адипоциты.
По мере взросления количество клеток бурой жировой ткани в указанных зонах снижается, но совсем не исчезает. По частоте встречаемости в организме одна клетка бурого жира, в среднем, приходится на 100-200 белых адипоцитов.
Функции
Функцией бурой жировой ткани является оперативное обеспечение организма тепловой энергией ( адаптивный термогенез ). Основной механизм термогенеза определяется тем, что в митохондриях клеток бурых адипоцитов процессы окисления и фосфорилирования не сопряжены (разобщены) и при окислении поступающих сюда жирных кислот происходит, в основном, выделение тепла, а не синтез АТФ.
Разобщение окисления и фосфорилирования обеспечивают специальные белки – термогенины (разобщающие белки, uncoupling proteins, UCP). Существуют несколько изоформ разобщающих белков: UCP-1 находится преимущественно в бурой жировой ткани, UCP-2 – в бурой и белой, UCP-3 – в cкелетных мышцах, UCP-4 и UCP-5 – преимущественно в мозге.
Адаптивный термогенез – это повышение теплопродукции в ответ:
- на изменение окружающей среды (снижение температуры) и на переохлаждение организма. При этом активируется симпатическая нервная система и стимулируется липолиз в бурых адипоцитах,
- на стимуляцию катехоламинами (через β3-адренорецепторы) – повышение транскрипции гена UCP-1, что приводит к увеличению энергозатрат,
- на избыток потребления пищи или изменение ее состава (поступление жиров),
- на секрецию лептина (усиление транскрипции гена UCP-1),
- на какие-то патологические воздействия.
У человека адаптивный термогенез ярко выражен у новорожденных, но при взрослении становится менее востребованным, заменяясь на иные способы сохранения тепла.
У взрослых отсутствие (резкое снижение) бурой жировой ткани встречается в 10% всех случаев ожирения.
Бурой жировой ткани много у животных, которые впадают в зимний сон и зимнюю спячку, температура тела у них поддерживается за счет адаптивного термогенеза.
Небольшие количества бурой жировой ткани у здоровых взрослых людей осуществляют постпрандиальный (возникающий после еды) термогенез , что в определенной степени препятствует отложению жира на фоне избытка пищи. При ожирении клетки бурой жировой ткани практически отсутствуют и, соответственно, процесс постпрандиального термогенеза не происходит.
Бежевая жировая ткань
Установлено, что помимо белой и бурой жировых тканей имеется промежуточная разновидность – бежевая жировая ткань. Эта ткань является промежуточной между белой и бурой жировыми тканями. Бежевые адипоциты первоначально имеют крайне низкое количество термогенина, но, как и классические бурые адипоциты, реагируют на определенную стимуляцию (понижение температуры, мышечная работа) началом высокой экспрессии термогенина и превращением в активные теплообразующие адипоциты.
При работе скелетных мышц в них повышается содержание особого белка — фактора транскрипции PGC-1a (peroxisome proliferator-activated receptor Gamma Coactivator 1-alpha), этот белок также экспрессируется в бурой жировой ткани , сердце и почках. Он стимулирует увеличение количества другого белка FNDC5 (fibronectin type III domain-containing protein 5).
У белка FNDC5 имеется две функции:
- Даже в очень низких концентрациях он увеличивает синтез мРНК белка термогенина в 70-100 раз.
- При его расщеплении образуется секретируемый мышцами полипептид, который был назван ирисином . Этот гормон является посредником между физическими нагрузками и различными тканями организма, в том числе белой и бурой жировыми тканями.
Вырабатываясь в работающих мышцах, ирисин транспортируется по крови в белые адипоциты, меняет акценты транскрипции и преобразует белую жировую ткань в бежевую.
Ирисин вырабатывается мышечной тканью не только при физической работе, но и при действии на организм холода. Так, при дрожании от холода в течение 10-15 минут вырабатывается такое же количество ирисина, как и при часовой езде на велосипеде со средней интенсивностью.
Название ирисин дано по имени древнегреческой богини Ириды, которая являлась богиней радуги и вестницей богов Зевса и Геры.
Интересно, что белок FNDC5 также стимулирует синтез нейротрофического фактора BDNF (Brain-Derived Neurotrophic Factor) в зубчатой извилине гиппокампа головного мозга, т.е. в той зоне которая участвует в процессах обучения и памяти. BDNF, в свою очередь, способствует образованию новых нейронов из стволовых клеток и новых синапсов, которые закрепляют новые знания и консолидируют память.
Получены данные взаимосвязи мозговой и физической активности, на основании которых предполагается, что посильные упражнения на выносливость для пожилых людей помогут им сохранить умственное здоровье.
Бурая жировая ткань особенности метаболизма механизм термогенеза биохимия

Бурая жировая ткань впервые была описана несколько сот лет назад. Эта ткань локализована близ жизненно важных органов и играет главную роль в производстве тепла, необходимого для поддержания постоянной температуры, в особенности у мелких животных, живущих в условиях холода и/или впадающих в зимнюю спячку, а также у новорожденных, в том числе и человека.
Читайте также: Кукла из ткани очень легко
Нейротрансмиттер норадреналин инициирует термогенез и увеличивает термогенную активность ткани при хроническом холодовом стрессе, стимулируя её гиперплазию и гипертрофию. Норадреналин многократно ускоряет как пролиферацию клеток, так и их дифференцировку. В бурых преадипоцитах норадреналин стимулирует синтез ДНК и белка по сАМР-зависимому пути. В постконфлуентных клетках норадреналин стимулирует экспрессию гена белка-разобщителя. Была показана линейная корреляция между экспрессией гена и повышением сАМР. Таким образом, процессы; пролиферации, дифференцировки и термогенеза, опосредованы одним и тем же мессенджером – сАМР и инициируются через β-адренорецепторы. Роль α1-адренорецепторов в указанных процессах оказалась минорной и сводилась к синергическому усилению сигнала, индуцированного через β-рецепторы и сАМР. В этом случае ионы Са2+ усиливали стимулирующее действие сАМР на термогенез клеток и экспрессию ряда генов [1,2].
Единственным указанием на возможное участие ионов Са2+ в активации пролиферации бурых преадипоцитов являлась установленная ранее корреляция между эффектами нейропептидов на [Са2+]i в свежевыделенных преадипоцитах и модуляцией пролиферации культивируемых клеток пептидами при тех же концентрациях. Однако оказалось, что [Са2+]i, инициируемый норадреналином и пептидами в преадипоцитах и зрелых клетках бурого жира отличается по ряду параметров (кинетика, амплитуда и т.д.). Более того, создавалось впечатление, что Са2+-сигналы, инициируемые в преадипоцитах разными агонистами, обладают разными физиологическими функциями в этих клетках и связаны, по-видимому, со стимуляцией противоположно направленных процессов. Поэтому мы считаем весьма важным на первом этапе установить, причины существенных отличий Са2+-ответов на норадреналин в преадипоцитах и зрелых клетках бурого жира. Это позволит в последующем использовать полученные знания для контролируемого управления за развитием клеток и ткани.
К важным свойствам бурой жировой ткани относится её способность наращивать свою массу в случае хронического холодового стресса. Запуск этого процесса, также как и термогенеза, осуществляется в основном норадреналином. Бурая жировая ткань представляет собой хорошую модель для исследования механизмов гормонального контроля за развитием ткани и клеток 2.
Материалы и методы исследования
Эксперименты проводили на суспензии свежевыделенных бурых преадипоцитов. Мышей-самцов линии NMRI в возрасте 3-5 недель содержали в отсутствие холодового стресса (температура в помещении 20-23оС). Забор ткани производили из затылочной, межлопаточной и пазушной областей. Бурую жировую ткань помещали в стандартный солевой буфер, содержащий (в мМ): NaCl – 123, KCl – 5, CaCl – 1,3, глюкозы – 5, HEPES – 100, BSA – 1,5%, pH – 7,4. Ткань бурого жира переносили в пробирки с раствором коллагеназы для разрушения межклеточного матрикса. Раствор коллагеназы (0,7-0,8 мг/мл изоляционного буфера) готовили непосредственно перед опытом из расчета 2–2,5 мл раствора на одну мышь. Измельченную ткань инкубировали в растворе коллагеназой на водяной бане при 28оС в течение 20-30 мин, периодически встряхивая на шейкере, и затем охлаждали на льду в течение 15-30 мин. Суспензию клеток фильтровали на нейлоновом фильтре (размер пор 250 мкм) и центрифугировали течение 10 мин при 1200 g. Слой белого жира удаляли и супернатант отбирали шприцем с длинной иглой. Полученный осадок ресуспендировали в 9 мл среды DMEM с добавлением 10 мМ HEPES и BSA (0,5 мг/мл), рН 7,4, отфильтровывали на нейлоновом фильтре (размер пор 25 микрон) и центрифугировали течение 10 мин при 1200 g. Осадок ресуспендировали в среде DMEM без альбумина. Зрелые адипоциты содержат жировые капли и не осаждаются при данных параметрах центрифугирования.
Измерения [Ca2+]i проводили спектрофлуориметрически с помощью внутриклеточных ион-селективных флуоресцентных зондов Fura-2/AM. В данной работе мы регистрировали кинетики [Са2+]i при аппликации агонистов адренорецепторов в течение 20 минут и далее учитывали и анализировали величину [Са2+]i на 20-й минуте. Статистическую обработку результатов экспериментов проводили с использованием программы Origin 7,0.
Результаты исследования и их обсуждение
1. Роль α1— и β-адренорецепторов в формировании Са2+-сигнала в преадипоцитах бурого жира.
В зрелых бурых адипоцитах норадреналин (НА) и α 1 -селективный агонист циразолин вызывали очень схожие Ca2+-ответы, в то время как влияние изопротеренола на [Ca2+]i было на два порядка меньше, чем влияние НА и циразолина. В свежевыделенных бурых преадипоцитах эффективность указанных агонистов была совершенно иной. Агонисты α 1 -адренорецепторов: циразолин, фенилефрин (не показано) и оксиметазолин (агонист α1А-рецепторов, которые составляют 80-90 % от α 1 -адренорецепторов в бурой жировой ткани) вызывали в 2-3 раза меньшее увеличение [Ca2+]i по сравнению с НА, тогда как изопротеренол и НА инициировали в них практически идентичные Са2+-ответы (рис. 1). Агонист α2-адренорецепторов клонидин не имел эффекта в бурых преадипоцитах.
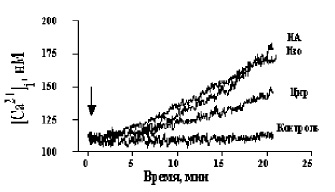
Рис. 1. Кинетика Са2+-ответов в бурых преадипоцитах, инициированных 3 mМ циразолином (Цир), 3 mМ изопротеренолом (Изо) и 3 mМ норадреналином (НА), контроль
При действии адренергических агонистов наблюдался аддитивный Са2+-ответ, стимулированный через β- и α1-адренорецепторы; [Ca2+]i при аппликации НА была примерно равна арифметической сумме эффектов изопротеренола и оксиметазолина. Совместная аппликация β- и α1-агонистов при максимальной концентрации 10 μМ последних дала в результате Са2+-ответы, аналогичные эффектам НА. Для того, чтобы подтвердить относительный вклад α1- и β-адренорецепторов в НА- иницированный Ca2+-ответ, были исследованы эффекты α1/2- и β-селективных антагонистов- фентоламина и надолола, соответственно. Надолол на 90 % подавлял НА- иницированный Ca2+-ответ, в то время как фентоламин уменьшал Са2+-ответ только на 25 % (рис.2). Таким образом, оба типа адренорецепторов (α1- и β-), вносят вклад в формирование [Ca2+]i, однако главная роль в формировании Са2+-ответа в бурых преадипоцитах принадлежит β-адренорецепторам 1.
Читайте также: Обивочная ткань для мебели остатки
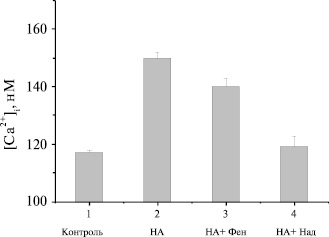
Рис. 2. Влияние селективных антагонистов α1- и β-адренорецепторов на НА-иницированный Са2+-ответ в бурых преадипоцитах: 1 – контроль; 2 – 6 mМ норадреналина (НА); 3 – НА + 10 mМ фентоламина; 4 – НА + 10 mМ надолола. (n=5). Уровень [Ca2+]I измеряли через 20 мин после добавления антагонистов
2. Действие ингибиторов и активаторов сАМР-зависимого пути на Са2+-сигнал в преадипоцитах.
Активация β-адренорецепторов клеток бурого жира приводит к опосредованной Gs-белками стимуляции аденилатциклазы (АЦ) и к образованию вторичного мессенджера сАМР. Увеличение внутриклеточного уровня сАМР можно достичь либо добавлением к клеткам проникающего через плазматическую мембрану аналога сАМР-BrcAMP, либо добавлением форсколина- непосредственного активатора АЦ. На рис. 3 представлены кинетики Са2+-ответов бурых преадипоцитов на норадреналин, BrcAMP и форсколин. Как видно из рисунка, увеличение [Са2+]i в ответ на форсколин и BrcAMP (кривые 3 и 4) заметно выше, чем при аппликации НА, что еще раз подчеркивает ведущую роль β-адренорецепторов в формировании [Са2+]i.
Добавление 500 μМ BrcAMP вызывает максимальное увеличение [Са2+]i, что следует из колоколообразной кривой дозозависимости (рис. 4).
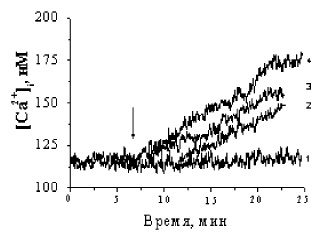
Рис 3. Изменение Са2+-ответа в свежевыделенных преадипоцитах под влиянием норадреналина, BrcAMP и форсколина: 1 – контроль; 2 – 1 μМ норадреналин; 3 – 10 μМ форсколин; 4 – 0,5 μМ BrcAMP
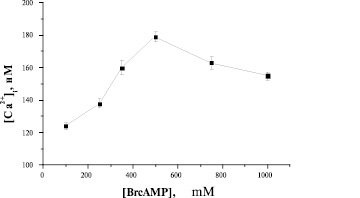
Рис. 4. Зависимость Са2+-ответа свежевыделенных преадипоцитов от концентрации BrcAMP
В настоящем исследовании показано, что воздействия, приводящие к увеличению сАМР в преадипоцитах, вызывают повышение [Са2+]i. Известно, что уровень сАМР в клетке определяется балансом между двумя процессами: процессом синтеза, опосредованным активацией аденилатциклазы, и процессом деградации, опосредованным активацией фосфодиэстеразы циклических нуклеотидов. Неспецифическим ингибитором фосфодиэстераз циклических нуклеотидов является соединение 3-изобутил-1-метилксантин (IBMX). Оптимальная концентрация IBMX для преадипоцитов бурого жира была выбрана в результате титрования вышеупомянутого ингибитора в широком диапазоне концентраций. Как следует из рисунка 5, 30 μМ является самой эффективной концентрацией. Таким образом, ингибирование фосфодиэстераз в присутствии IBMX вызывает увеличение уровня сАМР, что в свою очередь увеличивает кальциевый ответ на НА 5.
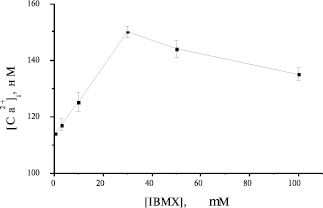
Рис. 5. Зависимость Са2+-ответа на НА от концентрации IBMX в преадипоцитах
Показано, что в адипоцитах бурого жира наиболее широко представлены фосфодиэстеразы 3- и 4-типов. Селективными ингибиторами фосфодиэстераз 3- и 4-типов являются, такие соединения как ОРС-3911 и Ro-20-1724, соответственно.
Ранее было показано, что ингибиторы фосфодиэстераз 3- и 4-типов синергично воздействовали на увеличение внутриклеточного уровня сАМР. Подобное явление наблюдалось также на культивируемых зрелых бурых адипоцитах. Совместное применение этих ингибиторов в данном исследовании на бурых преадипоцитах вызывало меньшее усиление Са2+-ответа на НА, чем вызывали эти ингибиторы по отдельности (рис. 6). На наш взгляд это полностью согласуется с колоколообразными кривыми дозозависимости эффекта BrcAMP (рис. 3) и IBMX (рис. 4) на величину Са2+-ответа в бурых преадипоцитах.
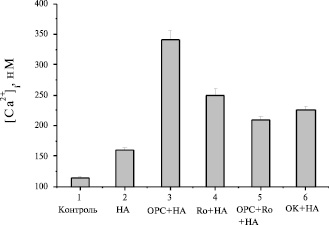
Рис. 6. Влияние ингибиторов фосфодиэстераз и фосфатазы на инициированный НА Ca2+-ответ клеток: 1 – контроль; 2 – 6µМ НА; 3 – 0,5 µМ OPC-3911+НА; 4 – 15 µМ Ro 20-1724+НА; 5 – OPC+Ro+НА; 6 – 30 µМ OK+НА
Известно, что активность Са2+-каналов зрелых клеток модулируется процессами фосфорилирования и дефосфорилирования, осуществляемыми протеинкиназой А и серин- треонин специфической фосфатазой. В данном исследовании установлено, что специфический ингибитор протеинкиназы А (Н-89) подавлял Са2+-ответы на изопротеренол дозозависимым образом (не показано) и форсколин. Максимальное подавление Са2+-ответа на форсколин ингибитором ПКА ([H-89]= 1 µM) составляло 55 % (рис. 7). Добавление ингибитора фосфатазы, окадаиковой кислоты, существенно увеличивало (рис. 6) Са2+-ответ на НА. Эти факты говорят в пользу того, что ПКА, фосфатаза и фосфодиэстераза участвуют в процессе формирования Са2+-ответа на норадреналин в бурых преадипоцитах.
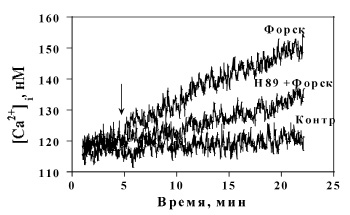
Рис. 7. Влияние ингибитора протеинкиназы А H-89 (1 μМ) на кинетику Ca2+-ответа, инициированного в бурых преадипоцитах форсколином (10 µM)
Таким образом, в данной работе показано, что Са2+-ответ в свежевыделенных бурых преадипоцитах инициируется главным образом через b-адренорецепторы и далее опосредуется через сАМР/протеинкиназа А-зависимый путь. Это является отличительной чертой бурых преадипоцитов. В зрелых адипоцитах, Са2+-ответы инициируются через α1-адренорецепторы.
Второй отличительной чертой Са2+-ответа в бурых преадипоцитах является колоколообразная зависимость его величины от концентрации агонистов адренорецепторов или сАМР; в зрелых клетках эти зависимости имели гиперболический или сигмоидальный вид. Согласно классическим представлениям колоколообразная дозозависимость в ответ на действие гормона или нейротрансмиттера указывает на его двойственное действие: он является активатором при низкой концентрации и ингибитором при высокой. Ранее были описаны подобные колоколообразные дозозависимости аккумуляции сАМР в зрелых бурых адипоцитах при аппликации норадреналина и изопротеренола. Ингибирующее действие высоких концентраций (1-10 μМ) НА и изопротеренола в этом случае было обусловлено активацией ими Са2+-сигнала. Возросший уровень [Ca2+]i приводил к активации СаМ/Са2+-зависимой фосфодиэстеразы и расщеплению сАМР. Колоколообразный характер кривых может быть связан с наличием отрицательной обратной связи, так например, показано на кардиомиоцитах, что при адренергической стимуляции L-типа Са2+-каналов увеличение [Ca2+]i приводит к подавлению аккумуляции сАМР, связанное в одном случае с Са2+- индуцированным ингибированием аденилатциклазы, в другом- с активацией СаМ-зависимой фосфодиэстеразы. Более того, с помощью моделирования показано, что наличие двух участков фосфорилирования Са2+-каналов в большей степени соответствует экспериментальным данным о двойственном эффекте изопротеренола на кардиомиоцитах 6.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом