
Жировая ткань относится к соединительным тканям со специальными свойствами. Развивается из мезенхимы. После рождения обновление идет за счет адвентициальных клеток.
Различают два вида жировой ткани: белую и бурую. Бурая жировая ткань характерна лишь для раннего детского возраста. У взрослых может находится в средостении, вдоль аорты. Белая жировая ткань встречается в подкожной жировой клетчатке, в сальниках, в строме внутренних органов, орбитах.
Белая жировая ткань состоит из адипоцитов (липоцитов), содержащих одну крупную каплю жира. Липоциты имеют округлую форму, в центре крупная капля жира, а вокруг узкий ободок цитоплазмы, содержащий митохондрии, комплекс Гольджи, ЭПС и палочковидное ядро.
Клетки бурой жировой ткани содержат мелкие капельки жировых включений, много митохондрий с пластинчатыми кристами. Митохондрии здесь отличатся мелкими размерами, плотным матриксом, расширенным межмембранным пространством. Такие митохондрии способны активно захватывать воду и набухать, при этом при помощи белка термогенина усиливается разобщение окислительного фосфорилирования и происходит выделение тепла. В центре клетки находится округлое ядро, содержащее эухроматин. Бурый цвет ткани придают железосодержащие пигменты — цитохромы митохондрий. Клетки бурой жировой ткани окружены многочисленными капиллярами.
На данной фотографии (рисунок 20) представлен фрагмент адипоцита бурой жировой ткани, содержащий участок цитоплазмы и часть ядра. Видно, что ядро расположено в центре клетки, имеет округлую форму, содержит эухроматин. В кариолемме заметны поры. Митохондрии с пластинчатыми кристми многочисленны. Липидные включения заполнены гомогенным содержимым.
Бурая жировая ткань участвует в терморегуляции.
Белая жировая ткань — обеспечивает поглощение из крови, синтез и накопление нейтральных липидов. Выполняет трофическую функцию, связанную с обеспечением энергетического запаса к резерва воды в организме.

Рисунок 21 — Остеоцит (увеличение в 10 000 раз):
ядро; 2 — цитоплазма; 3 — плазмолемма;
4 — отросток остеоцита; 5 — лакуна
Остеоцит
Остеоциты — это зрелые, высокодифференцированные клетки костной ткани. Имеют отросчатую форму, темное компактное ядро и слабоба-зофилъную цитоплазму. Некоторые остеоциты имеют развитые мембранные структуры, другие находятся на различных стадиях деструкции.
Остеоциты располагаются в костных полостях — лакунах. Их тонкие отростки проходят в костных канальцах, пронизывающих основное вещество. При помощи этих канальцев происходит обмен веществ между остеоци-тами и кровью. Остеоциты не делятся, но участвуют в процессах метаболизма, обновления межклеточных структур и поддержания ионного баланса. Функция остеоцитов сводится к участию в обменно-транспортных процессах и регуляции минерального состава костной ткани.
На электронограмме (рисунок 21) представлены структуры костной ткани: клетка и межклеточное вещество:
1. Остеоцит лежит в ячейке (лакуне). Кроме тела остеоцита в лакуне находится аморфный компонент межклеточного вещества, который сооб щается с жидкостью в костных канальцах. За пределами ячейки располо жено сильно минерализованное электронно-плотное межклеточное веще ство. Клетка отросчатой формы. Отростки лежат в костных канальцах. Видны два отростка.
Ядро остеоцита повторяет форму тела клетки. В ядре не видны ядрышки, преобладает гетерохроматин (активность считывания информации с ДНК, а значит и синтеза — низкая).
Цитоплазма остеоцита скудная. Вокруг ядра различимы единичные цистерны и пузырьки.
2. Из-за сильной минерализации межклеточное вещество не пропус кает электронов и выглядит абсолютно черным. Поскольку остеоцит не синтезирует межклеточное вещество, то неминерализованной костной тка ни на данной электронограмме нет.

Рисунок 22 — Фрагмент поперечно-полосатого мышечного волокна
S — саркомер; А — анизотропный диск; I — изотропный диск;
Z — телофрагма; Н — светлая полоса в центре А-диска
в середине которой проходит М-линия (мезофрагма)
Фрагмент поперечно-полосатого мышечного волокна
На электронограмме (рисунок 22) представлен фрагмент миосимпла-ста. Мышечное волокно — это структурно-функциональная единица поперечно-полосатой скелетной мышечной ткани, которая развивается из миотомов сомитов мезодермы.
Каждое волокно покрыто сарколеммой, состоящей из двух слоев: внутреннего — плазмолеммы и внешнего — базальной мембраны, в которую вплетаются ретикулярные волокна. Многочисленные ядра занимают периферическое положение. В саркоплазме содержатся включения миог-лобина и гликогена, свободные рибосомы в виде полисом. Присутствуют лизосомы, много митохондрий, хорошо развита агранулярной ЭПС (депо Са++), клеточный центр отсутствует. Триада — это комплекс из одной инвагинации плазмолеммы (Т-трубочка) и двух цистерн агранулярной ЭПС (L-трубочки). Обеспечивает проведение возбуждения от плазмолеммы к мембранам ЭПС.
Основной объем саркоплазмы занят сократительным аппаратом — миофибриллами, которые являются органеллами специального назначения. Миофибриллы состоят из уложенных параллельными рядами миофила-ментов. Миофиламенты — это нити сократительных белков. Тонкие фила-менты содержат актин, тропомиозин, тропонин. Толстые филаменты состоят из миозина. Упорядоченное расположение миофиламентов придает миофибрилле поперечную исчерченность (видны правильно чередующиеся темные и светлые диски).
Саркомер — это структурно-функциональная единица миофибриллы, участок между телофрагмами (рисунок 23.1).

Рисунок 23.1 — Саркомер поперечно-полосатого мышечного волокна
1 — мезофрагма; 2 — толстые миозиновые филаменты;
3 — тонкие актиновые миофиламенты; 4 — Z-телофрагма;
5 — часть 1-диска; 6 — М-линия; 7 — А-диск; 8 — саркомер
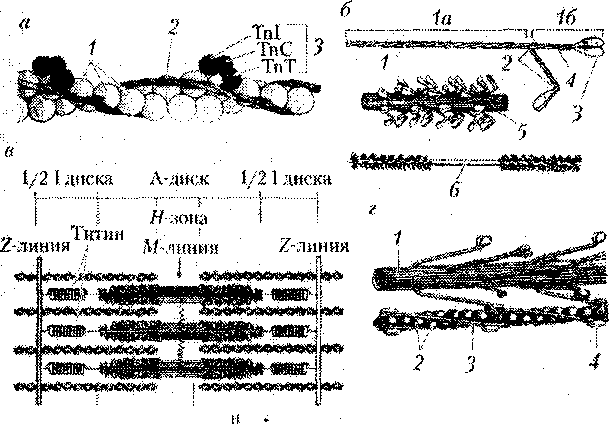
Рисунок 23.2 — Схема строения миофиламентов
Саркомер поперечно-полосатого мышечного волокна
На электронограмме (рисунки 23.1, 23.2) представлен саркомер — структурно-функциональная единица миофибриллы поперечно-полосатой мышечной ткани.
Саркомер — это часть миофибриллы между телофрагмами (Z-линиями). Формула саркомера 1/2 I-диска + А-диск +1/2 I-диска. Линия сшивки соседних саркомеров (Z-линия) состоит из белков алъфа-актинина, десмина, виментина.
Миофибриллы состоят из уложенных параллельными рядами мио-филаментов. Миофиламенты — это нити сократительных белков. Тонкие филаменты — актин, тропомиозин, тропонин. Толстые филаменты — миозин. Упорядоченное расположение миофиламентов придает миофибрилле поперечную исчерченность, т. е. видны правильно чередующиеся темные и светлые диски.
В поляризованном свете темные диски обнаруживают двойное лучепреломление (анизотропные, А-диски). В середине А-диска находится светлая полоска Н-полоска. Там находятся только толстые миозиновые нити, которые прикрепляются в центре А-диска к М-линии (мезофрагма)
Светлые диски называют изотропными (I-диски). 1-диски состоят только из тонких филаментов. В центре диска видна телофрагма (Z-линия). Это место прикрепления тонких филаментов.
При сокращении тонкие актиновые нити глубоко заходят между мио-зиновыми и продвигаются к средней линии. При этом ширина I-диска и Н-полоски уменьшается, А-диска не изменяется.

Рисунок 24 — Различия в структуре и конфигурации
Читайте также: Чем отличается ткань поплин от перкали
вставочных дисков сердечной мышцы
А — вставочный диск в миокарде предсердий;
Б — вставочный диск в миокарде желудочков;
В — слоистые структуры типа десмосом вставочных дисков желудочков
Метаболическая активность и термогенез в бурой жировой ткани – чувствительный маркер тиреоидного статуса
У пациентов с гипертиреозом наблюдалось повышение содержания бурой жировой ткани в надключичной области, а изменение уровня свободного трийодтиронина при раннем эутиреозе корректировало с метаболической активностью адипоцитов и скоростью обмена веществ.
Бурая жировая ткань (БЖТ) – один из двух типов жировой ткани млекопитающих, которая хорошо развита у новорожденных детей и животных, впадающих в спячку. Адипоциты БЖТ содержат много липидных капель (в отличие от одной капли в клетках белой жировой ткани), богаты митохондриями и кровеносными сосудами и иннервируются волокнами симпатической нервной системы. Морфологические характеристики ткани обусловлены способностью адипоцитов БЖТ к несократительному, т.е. не связанному с мышечной активностью термогенезу.
В процессе эмбрионального развития БЖТ появляется на 5-м месяце беременности, и у новорожденных детей ее масса составляет до 5% массы тела. У взрослых содержание бурого жира снижается, однако ткань остается метаболически активной и участвует в процессах энергетического обмена, окисления липидов и инсулинорезистентности. В экспериментальных исследованиях продемонстрирована чувствительность БЖТ к гормонам щитовидной железы, в частности трийодтиронину (Т3), выступающему в роли ключевого регулятора дифференцировки и метаболической активности адипоцитов.
На основании данных экспериментальных исследований, демонстрирующих прямую связь между активностью адипоцитов БЖТ и уровнем Т3 в модели гипер- или гипотиреоза, ученые из Института клинических наук Сингапура предположили, что уровень, метаболическая активность и термогенез в БЖТ у пациентов с заболеваниями щитовидной железы будет меняться в зависимости от статуса заболевания – гипертиреоза или эутиреоза.
Больным диффузным токсическим зобом (болезнь Грейвса; n=21, 5 мужчин; 39.5±2.5 лет) была назначена терапия антитиреоидными препаратами группы тионамидов (карбимазол, тиамазол). На фоне гипертиреоза и при диагностике раннего эутиреоза у пациентов измеряли клинические и биохимические маркеры и общие энергозатраты. Для оценки содержания БЖТ больным проводили инфракрасную термографию надключичной области. По данным ПЭТ/МРТ определяли средний и максимальный стандартизированный уровень захвата 18 F-флуородеоксиглюкозы (mean/maximum standardized uptake value, SUVmean/max) – индикатор метаболической активности адипоцитов, фракцию бурого жира в надключичной ямке (ФБЖН) и локальное повышение температуры в ткани как маркер увеличения ее метаболической активности (mean temperature, Tscv).
Среднее время перехода от гипертиреоза к эутиреозу на фоне антитиреоидной терапии составило 28.6±2.3 недель.
По данным ПЭТ/МРТ наличие БЖТ было выявлено у 8 пациентов с гипертиреозом (ФЖН от 72.3±1.4% до 76.8±1.4%; P 18 F-флуородеоксиглюкозы клетками БЖТ (r=0.58, P=0.01) и скоростью обмена веществ в покое (P
Бурая жировая ткань цвет ткани определяют цитохромы митохондрий
Челябинский областной клинический онкологический диспансер
Кафедра пластической и эстетической хирургии, косметологии и клеточных технологий ГБОУ ВПО «Российский национальный исследовательский медицинский университет им. Н.И. Пирогова» Минздравсоцразвития России
Российский научный центр хирургии им. акад. Б.В. Петровского РАМН, Москва
МАУЗ ОЗП «Городская клиническая больница», Челябинск, Россия
Биологическая характеристика жировой ткани
Журнал: Пластическая хирургия и эстетическая медицина. 2019;(2): 33-42
Васильев В. С., Мантурова Н. Е., Васильев С. А., Терюшкова Ж. И. Биологическая характеристика жировой ткани. Пластическая хирургия и эстетическая медицина. 2019;(2):33-42.
Vasil’ev V S, Manturova N E, Vasil’ev S A, Teryushkova Zh I. Biological features of adipose tissue. Plastic Surgery and Aesthetic Medicine. 2019;(2):33-42. (In Russ.).
https://doi.org/10.17116/plast.hirurgia201902133
Челябинский областной клинический онкологический диспансер



Жировая ткань представляет собой биологически активную субстанцию. Помимо депонирования энергетических запасов и участия в формировании контуров тела, определяющих внешний вид человека, жировая ткань является ключевым компонентом системы терморегуляции, оказывает регулирующее влияние на иммунную систему, секретирует большое количество биологически активных веществ. Жировая ткань является динамически функционирующим эндокринным органом, высвобождающим ряд гормонов, напрямую взаимодействующих с гипоталамусом. Стволовые клетки жировой ткани обладают высоким дифференцировочным потенциалом. Они могут дифференцироваться в клетки хрящевой, нервной, костной, мышечной, кардиальной, сосудистой, эндокринной, печеночной, гемопоэтической тканей. Эти свойства стволовых клеток жировой ткани открывают широкие возможности их клинического применения не только по косметическим, но и по реконструктивным показаниям. Доступность жировой ткани для забора из организма человека позволяет рассматривать ее в качестве перспективного субстрата для регенеративной медицины и тканевой инженерии.
Челябинский областной клинический онкологический диспансер
Кафедра пластической и эстетической хирургии, косметологии и клеточных технологий ГБОУ ВПО «Российский национальный исследовательский медицинский университет им. Н.И. Пирогова» Минздравсоцразвития России
Российский научный центр хирургии им. акад. Б.В. Петровского РАМН, Москва
МАУЗ ОЗП «Городская клиническая больница», Челябинск, Россия
Введение
Жировая ткань является биологической субстанцией, вызывающей всевозрастающий научный интерес. В течение продолжительного времени функция жировой ткани сводилась к сохранению энергетических запасов организма, получаемых с пищей. Однако оказалось, что жировая ткань секретирует молекулы, напрямую взаимодействующие с головным мозгом, и является компонентом иммунной системы. Эта ткань формирует форму человеческого организма, определяя сексуальную привлекательность, тем самым выполняя еще и социальную функцию. Большинство исследований жировой ткани было проведено на животных, что не позволяет экстраполировать эти данные на человека. Доступность жировой ткани для забора из организма человека открывает широкие возможности ее применения в качестве субстрата для клеточной терапии по различным показаниям.
Различают две разновидности жировой ткани человека: коричневую жировую ткань (brown adipose tissue — BAT) — КЖТ и белую жировую ткань (white adipose tissue — WAT) — БЖТ. Эти две разновидности выполняют различные функции, но по содержанию запасов итрацеллюлярного триглицерида относятся к жировой ткани [1, 2]. КЖТ участвует в механизме выработки энергии для организма, в то время как БЖТ имеет несколько функций: обеспечение термосбережения; определение формы тела, что особенно важно для сексуальной привлекательности женщин; сбережение энергии; амортизация при механических ударах; выполнение эндокринной функции; заполнение свободных пространств организма; облегчение скольжения мышц при их движении. КЖТ и БЖТ обычно не имеют четких границ, а существуют в виде смешанной субстанции [1, 3]. У женщин и мужчин с нормальной комплекцией жировая ткань составляет 22 и 15% от массы тела соответственно. Жировая ткань широко распространена по всему организму, при этом существуют различия в ее распределении у мужчин и женщин. Количество жировой ткани в организме может изменяться в зависимости от особенностей питания и энергозатрат. При поступлении в организм избыточной энергии жировая ткань может образовывать вокруг кровеносных сосудов недифференцированные клетки. На начальной стадии адипогенеза клетки не содержат жировых включений. Затем в стадии жировой пролиферации происходит аккумуляция жира, и преадипоциты дифференцируются в адипоциты с формированием островков жировой ткани. Если адипоцит теряет свою массу (в результате голодания и потери массы тела), клетка становится морфологически неравномерной, с увеличенными митохондриями в цитоплазме. Эти потерявшие жировое наполнение адипоциты (постадипоциты) окружены плотным коллагеновым матриксом, который может вызвать фиброз жировой ткани. Дольки жировой ткани окружаются соединительной тканью с формированием капсулы. Коллагеновая фрагментация продолжается до тех пор, пока каждый адипоцит не будет окружен коллагеновым матриксом (scaffold). Соединительнотканные перегородки внедряются в жировую ткань и разделяют ее на несколько более мелких фрагментов [1, 3].
Читайте также: Как удалить пятно жира от ткани
Жировая ткань имеет два типа нервных волокон: симпатические и чувствительные. Доказано, что симпатическая иннервация подавляет увеличение количества жировых клеток. Это может служить подтверждением связи некоторых синдромов ожирения со сниженной симпатической иннервацией [1, 3]. У мышей и крыс не обнаружено парасимпатической иннервации жировой ткани [4]. Прямая нейронная связь между паравентрикулярными ядрами гипоталамуса и жировой тканью установлена только в придатке яичка [5]. Такая же прямая гипоталамическая связь существует с печенью, что может обеспечивать контроль за метаболическими процессами в организме. Если учесть, что гипоталамус имеет множественные нейронные связи с другими отделами головного мозга, можно понять возможность непредсказуемых биологических взаимодействий. В межлопаточной области хомяков обнаружены прямые нейронные пути между центральной нервной системой и КЖТ. Нейроны средней преоптической области, гипоталамические ядра, вентромедиальные гипоталамические ядра, супрахиазматические и латеральные гипоталамические ядра соединяются с межлопаточной КЖТ через пути спинномозговых волокон, а также со стволом головного мозга, средним и передним мозгом [2].
В зависимости от анатомической локализации жировая ткань демонстрирует различные свойства. Основная масса ткани локализуется в двух отделах: подкожном слое и сальнике. Этот массив ткани легко увеличивается или уменьшается в зависимости от питания. Половые различия также влияют на количество и распределение жировой ткани. В то же время некоторые фрагменты жировой ткани, такие как периокулярные, ладонные, подошвенные, не вовлечены в метаболический процесс организма и их масса не изменяется даже в результате строгого поста. [1, 3]. Самые маленькие адипоциты, которые располагаются в брыжейке, наименее чувствительны к адреналину. Жировая ткань брыжейки имеет более богатые иннервацию и кровоснабжение. Кровоснабжение жировой ткани меняется в зависимости от индекса массы тела (ИМТ). Сердечный выброс в жировую ткань варьирует от 3—7% у худых людей, до 15—30% — при выраженном ожирении. Такое увеличение сердечного выброса может вызывать гемодинамические эффекты с возможным развитием кардиомегалии и выраженной сердечной недостаточности. Процентное содержание макрофагов в ткани также зависит от ИМТ: с увеличением массы тела увеличивается количество макрофагов, которые фагоцитируют жировые фрагменты погибших адипоцитов.
Адипоциты БЖТ содержат одну большую каплю жира в каждой клетке. Эта капля не имеет четкой дифференцировки, но у нее есть однослойная мембрана, отделяющая ее от цитоплазмы. Клетки бывают круглой или полигональной формы, и их размер варьирует от 25 до 200 мкм. Они содержат много органелл, однако трудноразличимы на фоне крупного жирового фрагмента, который выдавливает внутриклеточные структуры вместе с ядром к периферии, где они находятся в тонкой прослойке цитоплазмы под плазмолеммой. В процессе гистологической обработки жировой фрагмент исчезает, оставляя пустое пространство, которое под микроскопом визуализируется как кольцо. Большое скопление пиноцитотических везикул расположено возле плазматической мембраны. Для прокраски адипоцитов используют Sudan III или Scarlet Red staining. БЖТ содержит множество сосудов, обычно каждый адипоцит контактирует с кровеносным сосудом. Жировая ткань представлена зрелыми адипоцитами, преадипоцитами, постадипоцитами, мезенхимальными стволовыми клетками, клетками эндотелия, перицитами, мастовскими клетками, макрофагами, фибробластами, циркулирующими кровяными клетками, ретикулоцитами, нервными клетками. При этом зрелые адипоциты составляют ½ всей клеточной популяции.
КЖТ относят к жировой ткани, поскольку ее клетки содержат в цитоплазме триглицеридные депозиты. Коричневый цвет этой ткани обусловлен высоким содержанием цитохромной оксидазы в митохондриях, которая в избыточном количестве находится в цитоплазме. Различают две разновидности клеток КЖТ. Первая популяция эмбриологически образуется из того же источника, что и мышечные клетки, и составляет основу всех классических депозитов КЖТ. Вторая популяция формируется из клеток БЖТ [6] и распределяется среди белых жировых клеток, поэтому именуется как «коричневая в белом» («beige or brite»). Анатомическое распределение КЖТ и БЖТ различно. В течение первых 10 лет человеческой жизни КЖТ можно обнаружить практически в тех же местах, где находится БЖТ, например в межлопаточной области и на передней брюшной стенке. Однако с течением времени КЖТ исчезает из периферических отделов и концентрируется во внутренних отделах организма: в средостении, вокруг почек, надпочечников, аорты, сохраняясь там вплоть до восьмой декады жизни. Результаты недавних исследований, включающих 3604 пациента и основанных на исследовании 18F-FDG-PET/CT, показали, что объем и активность КЖТ более выражены у женщин. Мужской организм с возрастом теряет объем и активность КЖТ. Предполагается, что это связано с гормональными различиями, а именно — тестостероновой ингибицией экспрессии UCP1 mRNA в дозозависимом порядке. Коричневые адипоциты имеют полигональную или эллипсовидную форму, их диаметр варьирует от 15 до 50 мк. Большое количество жировых капель различных размеров располагается в цитоплазме, что придает адипоцитам мультиокулярый вид [2]. КЖТ продуцирует тепло (термогенезис), которое распространяется по всему организму за счет циркуляции крови. Регуляция термогенеза в основном осуществляется гипоталамусом. Поступающие от гипоталамуса сигналы вызывают высвобождение норэпинефрина, который индуцирует метаболизм жировых кислот в митохондриях коричневых адипоцитов. Выработка тепла происходит за счет функционирования высвобожденного протеина 1 (UCP1) во внутренней мембране митохондрий. Этот протеин в литературе также называют «термогенин». Он присутствует только в клетках КЖТ, являясь их маркером. UCP1 активируется в присутствии свободных жировых кислот, высвобождаемых из триглицеридов в результате воздействия неэпинефрин-активированных β-адренергических рецепторов, и подавляет генерацию АТФ в процессе оксидирования жира. В результате свободная энергия выделяется в виде тепла. Механизм выработки тепла до сих пор изучается. UCP1 скорее всего является переносчиком триглицеридов, а не протонов. Свободные протоны выбрасываются в цитоплазму из митохондрий, где они соединяются с жировыми кислотами, переводя их в растворимое состояние. Эти растворимые жировые кислоты возвращаются в митохондрии. При этом (согласно концепции расщепления) протоны не используются для продукции АТФ, а тепло генерируется за счет высвобождения протонов. Таким образом, жировые кислоты вновь переходят в нерастворимое состояние и транспортируются в цитоплазму с помощью UCP1. Остаточные продукты оксигенированного жира, вероятно, подавляют активность UCP1. КЖТ иннервируется симпатическими нервными волокнами. Выработка тепла зависит от особенностей воздействия секретируемого из нервной ткани норэпинефрина на рецепторы КЖТ. При активации β3-рецепторов происходит активация термогенеза, а при активации β2-рецепторов — его ингибиция. Хотя очевидно, что механизм терморегуляторного баланса существует, однако он нуждается в дальнейшем изучении. Сигналы от кожных терморецепторов по нервным путям поступают в ростральный отдел гипоталамуса, который является предмозговым центром системы КЖТ-терморегуляции [1, 3]. Помимо термогенерирующей функции, коричневые адипоциты секретируют ряд молекулярных элементов, а именно компоненты экстрацеллюлярного матрикса (ЭЦМ) (collagen IV, laminin, heparan sulphate, proteoglycan, fibronectin), аутокринные молекулы (adipsin, FGF-2, IGF-I, prostaglandins E2, prostaglandin Fa, adenosine), паракринные молекулы (NGF, VEGF-A, VEGF-B, VEGF-C, nitric oxide, angiotensinogen), эндокринные молекулы (fatty acids, leptin, adiponectin).
Читайте также: Краткий ответ соединительная ткань
ЭЦМ жировой ткани представлен коллагеновыми, ретикулярными, эластическими волокнами, сосудами, стромальной и лимфатической системой. ЭЦМ ткани очень важен для выживания зрелых адипоцитов, включающих большие жировые капли. Масса этих жировых капель намного превышает массу остальных структур клетки, поэтому механическая поддержка ЭЦМ предохраняет клетки от разрушения [7]. Кроме того, сама архитектура ЭЦМ перераспределяет векторы давления окружающих тканевых структур, что также снижает опасность разрушения [8]. ЭЦМ можно разделить на две части: базальную мембрану и межклеточную субстанцию. Каждый адипоцит имеет свою собственную базальную мембрану, состоящую из сети коллагеновых волокон 4-го типа, пластинки, гепарансульфат полигликана (heparan sulphate proteoglycan), перликана и энтактина. Основным компонентом межклеточной субстанции является коллаген 6-го типа [7]. Детальное рассмотрение этого протеина необходимо для лучшего понимания специфики жировой ткани.
Коллаген 6-го типа обладает интересными свойствами. Этот протеин связывается с коллагеном 4-го типа базальной мембраны и взаимодействует с протеогликанами и фибронектином. Коллаген 6-го типа образует структурную интеграцию ЭЦМ. Трехмерная форма (3D) этого коллагена состоит из тетрамеров, которые имеют множество ответвлений, мягкие корзиноподобные волокна, объединяющие различные структуры, такие как сосуды, нервы и другие коллагеновые волокна соединительной ткани, связанные или не связанные с базальной мембраной. Пространственная конфигурация коллагена 6-го типа различается в зависимости от тканевой локализации. Он может секретироваться из фибробластов, мышечных клеток, макрофагов, адипоцитов, а также из опухолевых клеток. В некоторых клетках коллаген 6-го типа усиливает миграцию и инвазию (макрофагоподобные клетки) и способствует выживанию, однако эти эффекты также могут вызывать пролиферацию опухолевых клеток. Коллаген повышает в непосредственной близости от себя количество цитокинов, лигандов, факторов роста и способен самостоятельно посылать сигналы окружающим клеткам. Мышечная слабость, возникающая при дефиците коллагена 6-го типа, подчеркивает его важное значение. Изучение свойств коллагена позволяет понять функционирование ЭЦМ жировой ткани. Плотность жировой ткани у свиньи составляет 4,1 (1,2) кг/м 3 , что примерно в 4 раза ниже плотности дермы (около 17 кг/м 3 ). Плотность ткани дает представление об упругонаследственности, что применительно к жировой ткани является важной информацией для определенных процессов тканевой инженерии, таких как возможности манипуляции с тканью, вакуумной аспирации и трансплантации. Плотность жировой ткани в основном обеспечивается трехмерной микроархитектурой коллагена 4-го типа [8].
Жировая ткань не только выполняет депонирующую функцию, но также является эндокринным органом [9]. Это ткань состоит из адипоцитов, преадипоцитов, фибробластов, стромально-васкулярных клеток, мастовских клеток, и все эти клетки секретируют биологические молекулы и могут вырабатывать как однотипные, так и дифференцированные вещества. При этом один секретируемый продукт может вырабатываться только строго определенным типом клеток, в то время как другой — тремя или четырьмя типами. Постоянная миграция макрофагов и мастовских клеток приводит к изменению общего количества клеток, что затрудняет оценку секретирующей функции жировой ткани. На существующий в БЖТ клеточный трафик влияет целый ряд параметров. Секретируемые жировой тканью продукты могут быть классифицированы в зависимости от их специфики следующим образом: компоненты ЭЦМ (collagen I, III, VI, V, laminin, heparan sulphate, proteoglycan, perlecan, entactin), аутокринные (glycerol, NEFA, monoglyceride, eicosanoids, oleoyl-esttrone, prostagalandin E2, I2, lipoprotein lipase, acylation-stimulating protein, fasting-induced adipose factor, cholestery1 ester transfer protein, tumor necrosis factor a, interleukin-6, complement components, angiotensinogen, VEGF, metalloproteinase inhibitors, matrix metalloproteinases, secreted protein acidic), паракринные (NEFA, monoglyceride, eicosanoids, oleoy1-estrone, prostaglandin E2,12, acylation-stimulating protein, fasting-induced adipose factor, cholesteryl ester transfer protein, tumor necrosis factor α, interleukin-6, IL-1β, IL-8, IL-10, IL-18, IL-17D, transforming growth factor-β, monocyte chemoattractant protein 1, macrophage migration inhibiting factor, complement components, haptoglobin, serum amyloid A3, plasmogen activator inhibitor-1, angeotensinigen, pigment epithelian-derived factor, adrenomedullin, VEGF, apelin, NGF, fibroblast growth factors, metalloproteinase inhibitors, matrix metalloproteniases, secreted protein acid), эндокринные (NEFA, leptin, resistin, omentin, estrogen, oleoy1-oestrogen, androgens, cortisol, cortisone, adiponectin, vistafin, vaspin, fasting-induced adipose factor, retinol binding protein, interleukin-6, IL-1β, IL-8, IL-10, IL-18, IL-17D, transforming growth factor-β, monocyte chemoattractant protein 1, macrophage migration inhibiting factor, complement components, haptoglobine, serum amyloid A3, plasminogen activator inhibitor-1, pigment epithelian-derived factor, adrenomedullin, VEGF, apelin
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом