
Бурые водоросли помимо хлорофилла имеют в хроматофорах бурый пигмент — фукоксантин, маскирующий остальные пигменты. Это многоклеточные водоросли, большинство которых растет на глубине 5-15 метров, однако некоторые растут на глубине от 40 до 100 метров. По размерам являются наиболее крупными из всех водорослей. Типичные представители — фукус, саргассум и ламинария сахарная, изображенная ниже.

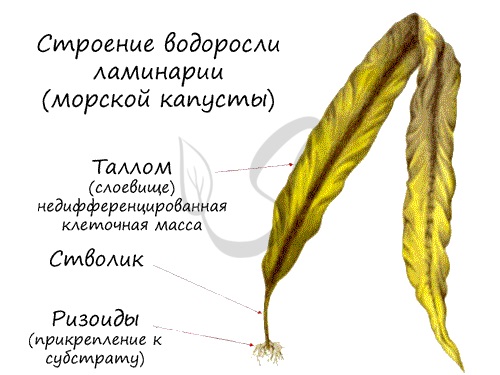
Строение ламинарии
Ламинария сахарная может достигать в длину до 60 м. Тело водоросли подразделяется на таллом (слоевище), стволик, ризоиды. Внешне напоминает лист на черешке. Вода всасывается всей поверхностью слоевища (таллома), ризоиды выполняют исключительно функцию прикрепления к субстрату.
Хроматофор имеет округлую форму, содержит помимо хлорофилла, каротина, ксантофилла бурый пигмент — фукоксантин. Запасное питательное вещество — ламинарин, откладывается в цитоплазме.
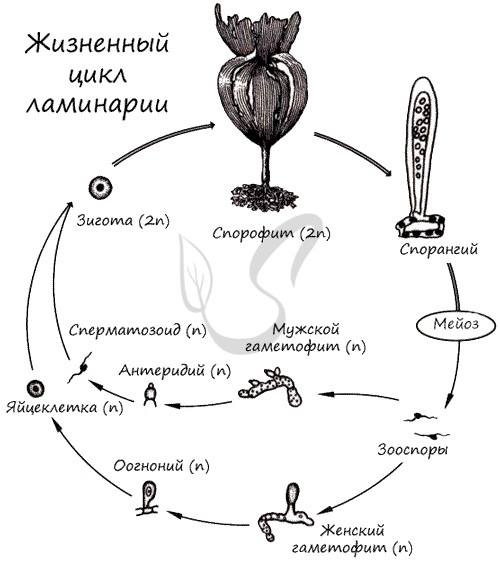
Жизненный цикл
Жизненный цикл представлен чередованием спорофита (2n, бесполое поколение) — имеет вид взрослого растения ламинарии, и гаметофита — гаплоидной фазы(n), представленной небольшими разветвленными нитями. В результате мейоза в зооспорангиях на спорофите (2n) образуются зооспоры (n). Зооспоры на почве прорастают в женские и мужские гаметофиты, на которых расположены соответственно оогонии с яйцеклетками (n) и антеридии со сперматозоидами (n). В результате оплодотворения образуется зигота (2n), из которой начинает расти взрослая особь — спорофит. Цикл замыкается.
Значение бурых водорослей
Бурые водоросли являются основным источником органического вещества в прибрежной зоне (являются звеном в цепи питания). Служат для морских животных местом размножения, укрытия и питания.
Не самое лучшее значение бурых водорослей состоит в том, что они очень любят участвовать в обрастании морских судов, ухудшая тем самым их характеристики.

© Беллевич Юрий Сергеевич 2018-2021
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Строение водорослей
Водоросли относятся к низшим растениям, наиболее примитивным: у них отсутствует разделение организма на стебель, корень и листья. Спешу заметить, что термин «низшие растения» — отжившее понятие, использовавшееся в ботанике до второй половины XX века.
Современная биология не считает дифференциацию тканей определяющим различием, сейчас существенным считают фундаментальные различия в строение клеток, обмене веществ. Тем не менее, во многих устаревших пособиях этот термин используется, и я обязан предупредить вас о нем.
Наука о водорослях называется альгология (от лат. alga — морская трава, водоросль и греч. λόγος — учение).
Среди водорослей есть одноклеточные и многоклеточные, некоторые водоросли достигают в длину 100-200 метров. Способ питания водорослей автотрофный: они синтезируют органические вещества в процессе фотосинтеза. Солнечный свет, проходя через толщу воды, рассеивается, что делает фотосинтез с увеличением глубины все труднее и труднее. Поэтому кроме хлорофилла они часто имеют и другие пигменты.
Клетки водорослей характеризуются наличием клеточной стенки (из целлюлозы и гликопротеинов — от греч. glykys сладкий (углеводы) + греч. prōtos — первый, важнейший (белок)) Органоиды располагаются в цитоплазме (син. — внеядерной протоплазме), где также располагается(-ются) один или несколько хроматофоров. Размножение происходит бесполым, вегетативным или половым путем.
Тело водорослей представлено слоевищем (син. — талломом) — недифференцированным скоплением клеток. С помощью ризоидов (от др.-греч. ῥίζα — корень и εἶδος — вид) водоросли прикрепляются к субстрату (камням, коралловым полипам), функцию всасывания ризоиды не выполняют. У водорослей отсутствуют настоящие ткани, механических тканей нет, так как таллом водоросли поддерживается (парит) в толще воды. Нет проводящих тканей: каждая клетка имеет доступ к воде напрямую, так что в клетку из окружающей воды поступает кислород, а в воду удаляется углекислый газ.
Хроматофор (от греч. chroma — цвет и phoros — несущий) — органелла в клетке водоросли, аналогичная хлоропласту и осуществляющая фотосинтез. Отличается от хлоропласта упрощенным строением, меньшим размером и иным составом хлорофилла. Внешне отличаются между собой по форме, хроматофор может быть: чашевидный, спиралевидный, в виде незамкнутых колец, цилиндрические, лентовидные, дисковидные. В хроматофорах находятся пигменты, которые придают окраску растению.
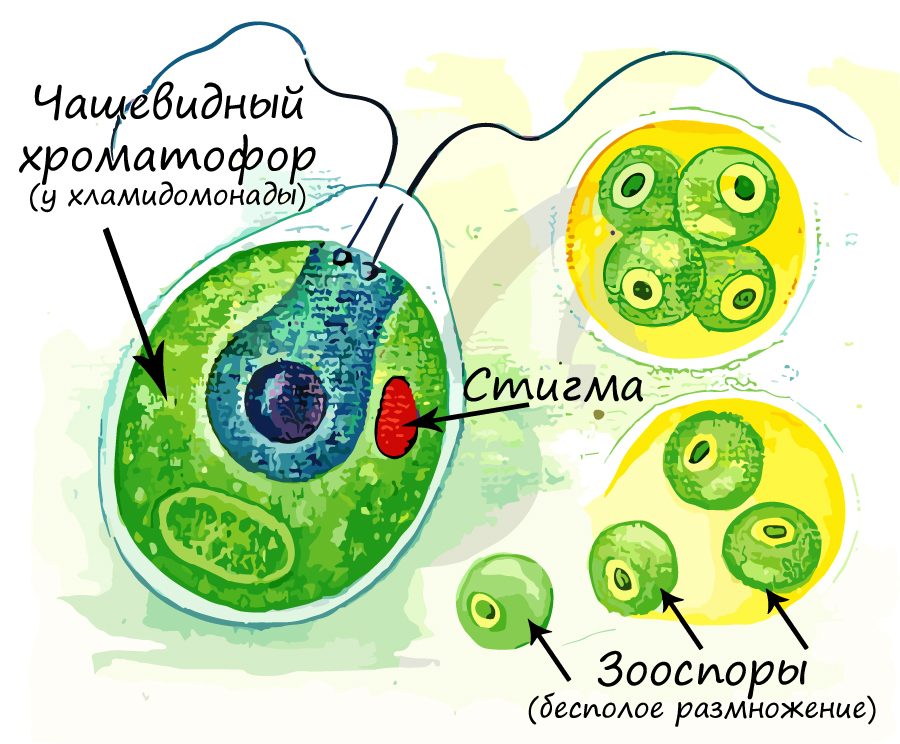
Система вакуолей в клетках водорослей развита отлично, в подвижных клетках водорослей можно обнаружить пульсирующие (сократительные) вакуоли. Их основная функция — поддержание постоянного осмотического давления внутри клетки. Вообразите: в глубине океана находится клетка водоросли, в которую постоянно поступает много воды. Если бы не было таких сократительных вакуолей, то клетка просто лопнула бы, но их работа обеспечивает удаление избытка воды.
Также у многих подвижных водорослей в клетках присутствует светочувствительный глазок (стигма), что обуславливает их чувствительность к свету — фототаксис. Подвижные водоросли стремятся занять как можно более освещенное место, чтобы активно шел процесс фотосинтеза.
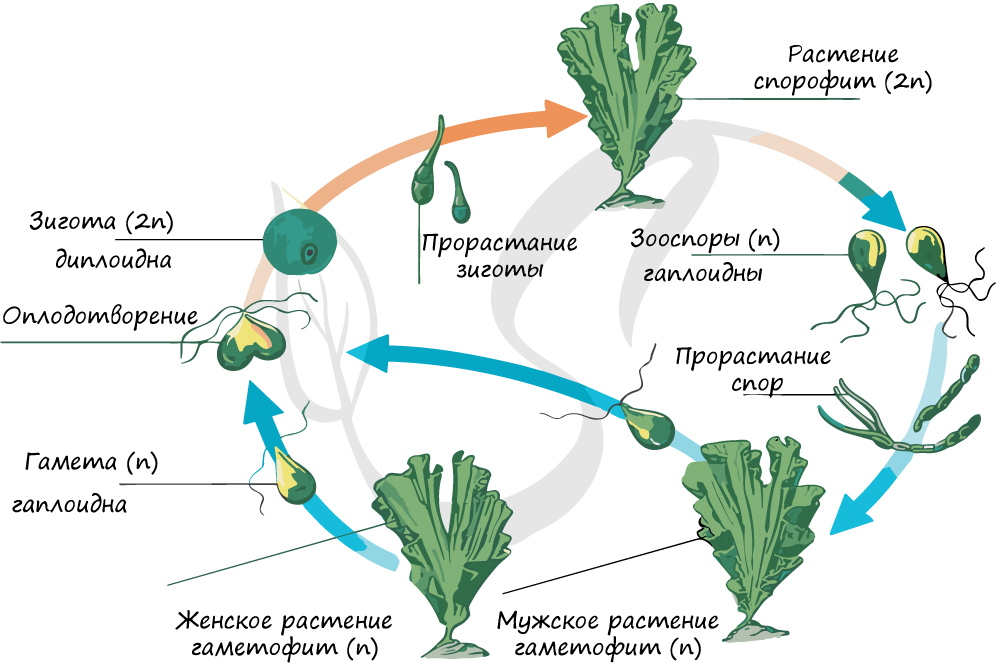
Жизненный цикл водорослей
Жизненные циклы водорослей разнообразны, обусловлены рядом экологических факторов. Мы разберем жизненный цикл на примере зеленой водоросли ульвы (морского салата).
Для начала отметим, что в целом жизненный цикл водорослей представляет собой чередование двух фаз: гаплоидной (гаметофита) и диплоидной (спорофита). Гаплоидной фазой называется фаза, при которой клеточные ядра содержат непарный (половинный) набор хромосом. К гаплоидной фазе всегда принадлежат гаметы: сперматозоиды, спермии (отличающиеся от сперматозоидов отсутствием жгутика), яйцеклетки.
При слиянии двух гамет: яйцеклетки (n) и спермия (n) образуется зигота (2n) из которой развивается спорофит (2n), таким образом, в спорофите восстанавливается диплоидный набор хромосом. В зооспорангии на спорофите в результате мейоза образуются зооспоры (n), которые делятся митозом, порастают и образуют мужские и женские гаметофиты (n). Клетки гаметофитов делятся митозом, образуются гаметы (n), которые сливаются в зиготу (2n), цикл замыкается.
Типы половых процессов
У водорослей выделяют несколько типов полового процесса:
- Изогамия — копулирующие элементы (гаметы) не отличаются друг от друга, подвижны
- Анизогамия — от греч. anisos неравный и gamos брак (гетерогамия) — при таком типе копулирующие элементы различаются по размерам, форме, величине, поведению
- Оогамия — от др. греч. ᾠόν яйцо и γάμος брак — копулирующие элементы резко отличаются друг от друга: крупная женская гамета без жгутиков обычно с мужской мелкой подвижной гаметой. Допустимо считать оогамию в некотором смысле подтипом анизогамии.
Читайте также: Одежда из вышитой ткани
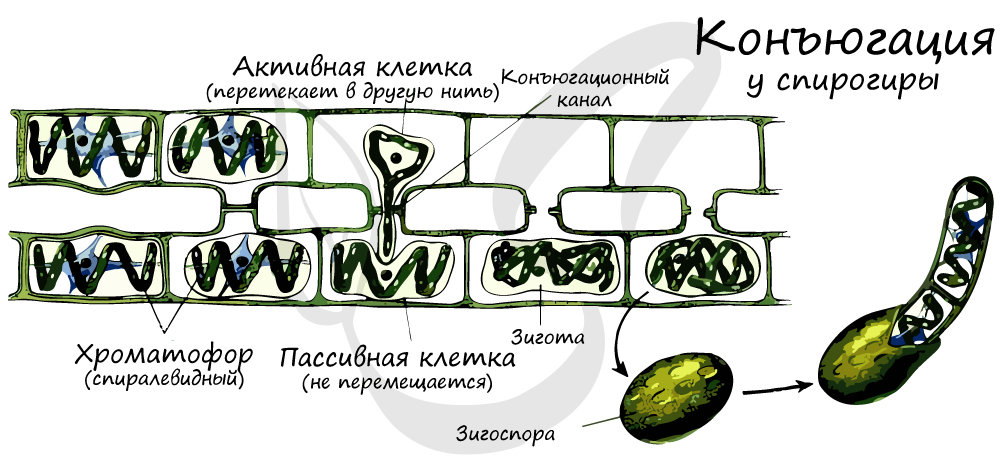
Особо стоит выделить тип полового процесса — конъюгацию. Конъюгация отличается тем, что сливаются не гаметы, а обычные вегетативные клетки, лишенные жгутиков. Клетки соединяются друг с другом с помощью боковых выростов, формируется копуляционный (конъюгационный) канал, по которому содержимое из одной клетки перетекает в другую — образуется зигоспора. В дальнейшем из зигоспоры развивается новая водоросль.
Отметим, что зооспора представляет собой подвижную клетку, которая способна двигаться в воде с помощью жгутиков. Образуется она в зооспорангии. Зооспора участвует в бесполом размножении у многих водорослей и простейших грибов. У некоторых водорослей имеются апланоспоры (гр. aplanes неподвижный + spora семя) — неподвижные безжгутиковые споры. Зооспоры и апланоспоры выходят в окружающую среду, разрывая стенки спорангия, в котором они находятся.
Значение водорослей
В Мировом океане водоросли составляют основную часть биомассы. Именно они являются главными продуцентами (производителями) органического вещества, преобразуя в ходе фотосинтеза энергию солнечного света в энергию химических связей. Значение водорослей для человека трудно переоценить: содержащиеся в них вещества необходимы для нормального роста и развития животных и человека (к примеру, морская капуста (ламинария) отличается большим содержанием йода.)

© Беллевич Юрий Сергеевич 2018-2021
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Бурые водоросли
- Аскозейровые (Ascoseirales)
- Хордариевые (Chordariales)
- Кутлериевые (Cutleriales)
- Диктиосифоновые (Dictyosiphonales)
- Десмарестиевые (Desmarestiales)
- Диктиотовые (Dictyotales)
- Хордариевые (Chordariales)
- (Discosporangiales)
- Эктокарповые (Ectocarpales)
- Фукусовые (Fucales)
- (Ishigeales)
- Ламинариевые (Laminariales)
- (Nemodermatales)
- (Onslowiales)
- (Ralfsiales)
- Сцитосифоновые (Scytosiphonales)
- (Scytothamnales)
- Сфацеляриевые (Sphacelariales)
- Спорохновые (Sporochnales )
- Тилоптеридовые (Tilopteridales)
- (Syringodermatales)
Бурые водоросли (лат. Phaeophyceae ) — отдел автотрофных хромистов. В жизненном цикле всех представителей присутствуют многоклеточные стадии. Преимущественно морские формы, лишь восемь видов перешли к существованию в пресных водоёмах.
Бурые водоросли включают 1500 видов, которые объединены в 265 родов, из которых достаточно известны Ламинария ( Laminaria ), Саргасс ( Sargassum ), Цистозейра ( Cystoseira ).
Бурые водоросли в хроматофорах содержат бурый пигмент фукоксантин (C40H56O6). Этот пигмент маскирует остальные пигменты.
В отличие от других водорослей, для бурых водорослей характерны многоклеточные волоски с базальной зоной роста.
Некоторые бурые водоросли, например, вакамэ, употребляются в пищу.
Строение
Среди бурых водорослей отсутствуют одноклеточные и колониальные формы, все особи многоклеточные. Встречаются как однолетние, так и многолетние виды, возраст которых может достигать 15-18 лет.
У бурых водорослей талломы могут быть микроскопическими или достигать нескольких десятков метров (например, у Macrocystis, Nereocystis). Форма талломов самая разнообразная: стелющиеся или вертикально стоящие нити, корочки, пластинки (простые или рассечённые), мешки, ветвящиеся кустики. Прикрепление талломов осуществляется с помощью ризоидов или подошвы. Для удержания в вертикальном положении у ряда бурых водорослей образуются воздушные пузыри, заполненные газом.
Наиболее сложно устроены талломы ламинариевых и фукусовых. Их слоевища имеют признаки тканевой дифференцировки со специализацией клеток. В их талломе можно различить: кору, состоящую из нескольких слоёв интенсивно окрашенных клеток; сердцевину, состоящую из бесцветных клеток, часто собранных в нити. У ламинариевых в сердцевине образуются ситовидные трубки и трубчатые нити. Сердцевина выполняет не только транспортную функцию, но и механическую, так как в ней находятся нити с толстыми продольными стенками. Между корой и сердцевиной у многих бурых водорослей может находиться промежуточный слой из крупных бесцветных клеток.
Рост таллома у бурых водорослей чаще всего интеркалярный и апикальный, реже базальный. Интеркалярный рост может быть диффузный или имеется зона роста. У крупных представителей интеркалярная меристема расположена в месте перехода «черешка» в «листовую пластинку». Крупные водоросли также имеют меристематическую зону на поверхности таллома, так называемую меристодерму (своеобразный аналог камбия высших растений).
Необычный тип меристемы, который встречается только у некоторых бурых водорослей, — трихоталлическая меристема, развитие клеток которой происходит в основании настоящих волосков. Настоящие волоски размещаются на поверхности меристодермы рассеянно или пучками и часто погружены своим основанием в особые углубления — криптосомы.
Жгутики
Жгутиковые стадии в жизненном цикле бурых водорослей представлены только гаметами и зооспорами. Два неравных жгутика прикреплены сбоку (сперматозоиды Dictyota имеют только один жгутик). Обычно длинный перистый жгутик направлен у бурых водорослей вперёд, а гладкий — вбок и назад, но у сперматозоидов ламинариевых, спорохналиевых и десмарестиевых, наоборот, длинный перистый жгутик направлен назад, а короткий гладкий — вперёд. Помимо трёхчастных мастигонем на длинном жгутике присутствуют чешуйки и шипики; его кончик может быть спирально закручен. В основании гладкого жгутика находится базальное вздутие. У сперматозоидов фукусовых вокруг жгутика имеется своеобразная воронкообразная структура — хоботок, поддерживаемая микротрубочками первого корешка.
Базальные тела жгутиков расположены под углом почти 110 градусов и соединены тремя исчерченными лентами. Типичной конфигурацией для бурых водорослей является наличие четырёх микротрубочковых корешков. Один корешок состоит из 7-5 микротрубочек, направлен к переднему концу клетки, где он загибается и идёт назад; другой корешок состоит из 5-4 микротрубочек и направлен в две стороны от базального тела — к переднему и заднему концу клетки; ещё два корешка — короткие, состоят каждый из одной микротрубочки. В корешковой системе отсутствует ризопласт. У ряда бурых водорослей строение корешковой системы отличается от описанного.
Покровы
Клеточная стенка бурых водорослей толстая, двух- или трёхслойная. Внутренний слой содержит преимущественно волокна из целлюлозы, внешний пектиновый слой содержит альгиновую кислоту, её натриевую соль, фукоидан и другие сульфатированные полисахариды.
У бурых водорослей целлюлоза составляет от 1-10 % от сухого веса таллома. Она встречается в виде микрофибрилл, синтезируемых терминальными комплексами, расположенными в плазмалемме. Терминальные комплексы бурых водорослей представляют собой линейные ряды плотно уложенных частиц. Каждая частица состоит из двух субъединиц. Такой линейный комплекс встречается у трибофициевых, красных и некоторых зелёных водорослей.
Читайте также: Конспект занятия в старшей группе опыты с тканью
Растворимые альгинаты входят в состав матрикса клеточной стенки, иногда на их долю приходится до 40 % сухого веса таллома [источник не указан 764 дня] .
Фуканы (фукоиданы или аскофилланы) — полимеры L-фукозы и сульфатированных сахаров. Их функция до конца не выяснена. Считается, что они играют важную роль в прикреплении зиготы и её прорастании у фукусовых водорослей.
У некоторых диктиотовых, например у Padina, в клеточных стенках откладывается известь в форме арагонита.
Клеточные структуры
В клетках бурых водорослей встречается от одной до многих пластид. Чаще хлоропласты мелкие, дисковидные, париетальные. Их форма может быть звёздчатой, лентовидной или пластинчатой; форма хлоропластов может меняться с возрастом клетки. Оболочка хлоропласта состоит из четырёх мембран; там, где хлоропласт расположен рядом с ядром, наружная мембрана хлоропластной эндоплазматической сети переходит в наружную мембрану ядра. Перипластидное пространство хорошо развито. Ламеллы трёхтилакоидные; имеется опоясывающая ламелла; хлоропластная ДНК собрана в кольцо.
В хлоропластах содержатся хлорофиллы a,с1, с2. Помимо фукоксантина у бурых водорослей присутствуют другие каротиноиды: β-каротин, виолаксантин, зеаксантин, антераксантин, неоксантин, диадиноксантин и диатоксантин.
Хлоропласты вегетативных клеток Ectocarpales имеют один или несколько грушеобразных пиреноидов, в то время как в вегетативных клетках у Ischigeales, Dictyotales, Laminariales, Sphacelariales хлоропласты без пиреноидов. Гаметы и зооспоры этих порядков содержат хлоропласты с рудиментарным пиреноидом.
Глазок состоит из 40-80 липидных глобул, собранных в один слой; расположен в хлоропласте и ориентирован на базальное вздутие. У бурых водорослей он выполняет функцию линзы, фокусирующей свет на жгутиковое вздутие, которое и является собственно фоторецептором. Для фототаксиса бурых водорослей более эффективным является свет с длиной волны 420 и 460 нм, что, возможно, связано с флавинподобными субстанциями в базальном вздутии заднего жгутика. В жгутиковых клетках ламинарии и близкородственных родов отсутствует фоторецепторный аппарат; у них нет ни глазка, ни базального вздутия. Глазок отсутствует у сперматозоидов десмарестиевых.
Основной запасной продукт — хризоламинарин (β-1,3-связанный глюкан). Он расположен в специальных вакуолях в цитоплазме около пиреноида. Встречаются маннит и липиды (в виде капель масла). Маннит — шестиатомный спирт, который помимо резервной функции выполняет осморегуляторную. Его концентрация внутри клетки зависит от изменения солёности воды.
В молодых клетках бурых водорослей содержатся мелкие и многочисленные вакуоли, которые с возрастом становятся крупнее за счёт слияния. Сократительные вакуоли отсутствуют. В цитоплазме расположены физоиды — везикулы, содержащие феофициновые таннины (флоротаннины). Флоротаннины — полимеры флороглюцина, известны только у бурых водорослей. Функция их, возможно, связана с защитой талломов от выедания животными, например такими, как гастроподы. Они, возможно, ингибируют поселение на поверхности таллома эпифитных водорослей и животных. Предполагают также, что они принимают участие в защите от радиационного повреждения и в аккумуляции тяжёлых металлов. Бесцветные флоротаннины на воздухе окисляются с образованием бурого или тёмного пигмента фикофеина, придающего высушенным бурым водорослям их характерную тёмную окраску.
Ядро чаще всего одно, но у некоторых представителей и с возрастом клетки изредка бывают многоядерными. Например, в клетках Durvillea содержится от 2 до 5 ядер. Ядрышко одно.
Плазмодесмы встречаются, вероятно, в поперечных перегородках всех бурых водорослей, даже у тех, у которых перегородка образуется за счёт впячивания мембраны. Через плазмодесмы осуществляется связь цитоплазмы соседних клеток.
Размножение
Вегетативное размножение у ряда бурых водорослей может осуществляться участками таллома, у видов Sphacelaria — выводковыми веточками; у Fucus на подошве имеется группа клеток, способных к дифференцировке в новый таллом.
Бесполое размножение происходит с помощью зооспор, у некоторых — неподвижными тетра- и моноспорами. Споры бесполого размножения формируются в результате мейоза и последующих митозов в одногнёздных спорангиях.
Половой процесс изо-, гетеро- и оогамный. Гаметы образуются в многогнёздных гаметангиях. В каждом гнезде (клетке) такого гаметангия формируется по одной гамете.
Для бурых водорослей известны половые феромоны, исследование которых активно проводится с 80-х годов прошлого века. Половые феромоны — это растворимые вещества, которые координируют активность клеток при половом размножении. Они активны на расстоянии до 0,5-1 мм. Феромоны бурых водорослей могут или стимулировать раскрытие антеридиев, или привлекать мужские гаметы к женским. Они также принимают участие в изоляции видов. Известно по крайней мере десять феромонов у бурых водорослей, первым из которых был открыт эктокарпен.
Полагают, что феромоны продуцирует большинство (если не все), бурых водорослей, имеющих половое размножение.
Жизненный цикл
У бурых водорослей, имеющих половое размножение, можно выделить два основных типа жизненных циклов. Один — гапло-диплобионтный жизненный цикл со спорической редукцией с изо- или гетероморфной сменой форм развития. Споры бесполого размножения формируются на диплоидных спорофитах, в одногнёздных спорангиях при их формировании происходит мейоз. Гаплоидные зооспоры и тетраспоры прорастают в гаплоидный гаметофит, на котором в многогнёздных гаметангиях формируются гаметы. После слияния гамет диплоидная зигота прорастает в диплоидный спорофит.
Другой тип жизненного цикла — диплобионтный с гаметической редукцией; редукционное деление происходит при образовании гамет. У бурых водорослей место мейоза доказано цитологически по наличию в пахитене синаптонемального комплекса.
Экология и значение
Бурые водоросли широко распространены во всех морях нашей планеты, наибольшего развития достигая в морях умеренных и приполярных широт. В тропиках массовое развитие бурых водорослей приурочено к зимним месяцам, когда понижается температура воды. В морях умеренных и приполярных широт бурный рост их талломов начинается весной, и наибольшего развития они достигают в летние месяцы. Наиболее густые заросли бурых водорослей формируются в верхней сублиторали до глубины 15 м, хотя встречаются от литоральной зоны и до глубины 40-120-200 м. На такую глубину, например в западной части Средиземного моря, проходит только 0,6 % света по отношению к поверхности воды. Ламинариевые могут формировать гигантские подводные леса, такие, например, как вдоль тихоокеанского побережья Северной Америки. Прикрепляются бурые водоросли к различным субстратам — скалам, камням, гравию, раковинам моллюсков, другим водорослям. Некоторые небольшие бурые водоросли живут внутри тканей других водорослей как эндофиты.
Читайте также: Букет цветов аппликация из ткани
В пресных водах встречаются только 8 видов, относящихся к родам Heribaudiella, Ectocarpus, Sphacelaria, Pseudobodanella, Lithoderma, Pleurocladia и Porterinema. Возможно, H. fluviatilis — обычный компонент речной флоры, но из-за незнания этой группы часто остаётся в пробах незамеченным.
Роль бурых водорослей в природе чрезвычайно велика. Это один из основных источников органического вещества в прибрежной зоне, особенно в морях умеренных и приполярных широт; их заросли служат местом питания, укрытия и размножения многих животных.
Бурые водоросли используют в пищу, на корм скоту, как удобрения, для производства альгинатов и маннита. Ежегодный сбор Laminaria и близких к ней водорослей достигает 2 млн т. сырой массы, более миллиона тонн даёт производство её марикультуры в Китае.
Альгинаты — нетоксичные соединения, обладающие коллоидными свойствами, поэтому они широко используются в пищевой и фармацевтической промышленности. Альгиновая кислота и её соли способны к 200-300-кратному поглощению воды, образуя гели, для которых характерна высокая кислотноустойчивость. В пищевой промышленности они используются в качестве эмульгаторов, стабилизаторов, желирующих и влагоудерживающих компонентов. Например, сухой порошковый альгинат натрия используют в производстве порошкообразных и брикетированных растворимых продуктов (кофе, чай, сухое молоко, кисели и др.) для их быстрого растворения. Водные растворы альгинатов используют для замораживания мясных и рыбных продуктов. В мире в пищевую промышленность идёт порядка 30 % получаемых альгинатов.
В текстильной и целлюлозно-бумажной промышленности альгинаты используют для загущения красок и усиления прочности их связи с основой. Пропитка тканей некоторыми солями альгиновой кислоты придаёт им водонепроницаемость, кислотоустойчивость и увеличивает механическую прочность. Ряд солей альгиновых кислот используют для получения искусственного шёлка. Во время Второй мировой войны в США и Англии из альгиновой кислоты и её солей производилось большое количество маскировочной ткани и сетей для жилых и промышленных зданий. Альгинаты применяются в металлургии как компонент формовочной земли, в радиоэлектронике — как связующий агент при изготовлении высококачественных ферритов, а также в горнодобывающей, химической и других отраслях промышленности.
В фармацевтической промышленности альгинаты используются для покрытия таблеток, пилюль, в качестве компонентных основ для различных мазей и паст, как гели-носители лекарственных препаратов. В медицине альгинат кальция используют как кровеостанавливающее средство, как сорбент, способствующий выведению радионуклидов (в том числе стронция).
В Северной Америке для получения альгинатов собирают Macrocystis и Nereocystis, на европейском побережье используют виды Laminaria и Ascophyllum. К концу двадцатого столетия ежегодное производство альгинатов в мире достигло 21 500 т: 12 800 т в Европе, 6 700 — в Северной Америке, 1 900 — в Японии и Корее, 100 — в Латинской Америке. В России в 1990 г. было получено всего 32 т пищевого альгината натрия.
Фукоиданы — эффективные антикоагулянты, даже более активные, чем гепарин. Перспективным считается их использование для получения противоопухолевых препаратов и антивирусных соединений. Даже в очень низких концентрациях они могут ингибировать прикрепление вирусов к поверхности клеток. Фукоиданы способны образовывать исключительно прочные и вязкие слизи, что находит применение в получении стабильных эмульсий и суспензий.
Маннит используют как заменитель сахара для больных диабетом. Кроме того, он может быть использован в качестве плазмозаменителя при консервации крови.
Клетки многих бурых водорослей накапливают йод. Его содержание может достигать 0,03 %-0,3 % от свежей массы водорослей, в то время как его содержание в морской воде достигает только 0,000005 % (0,05 мг на литр воды). До 40-х гг. XX в. бурые водоросли использовали для добычи йода.
Энергетический кризис, который охватил в последние годы многие страны мира, привёл к необходимости поиска новых нетрадиционных источников энергии. Так, в США с этой целью изучается возможность разведения водоросли Macrocystis pyrifera с последующей переработкой в метан. Подсчитано, что с площади 400 квадратных километров, занятых этой водорослью, можно получить 620 млн кубических метров метана.
В последние годы бурые водоросли привлекают внимание в связи со способностью выделять в атмосферу органические бромиды (бромоформ, дибромохлорметан и дибромометан). Ежегодный выброс водорослями органических бромидов достигает 10 000 тонн, что сравнимо с образованием этих веществ промышленностью. Существует мнение о связи выделения органических бромидов с разрушением озона в атмосфере Арктики.
Филогения
Ископаемые находки, которые могут иметь отношение к бурым водорослям, датируются поздним ордовиком (около 450 млн лет) и известны как Winnipegia и Tallocystis из среднего силура (425 млн лет). Но эти находки нельзя точно привязать только к бурым водорослям, так как они схожи и с некоторыми современными зелёными и красными водорослями. Ископаемые находки, которые точно можно связать с современными бурыми водорослями, относятся к миоцену (5-25 млн лет). Это Zonarites и Limnophycus, напоминающие современную Dictyota и др. Молекулярные методы определяют возраст бурых водорослей как 155—200 млн лет.
Бурые — монофилетичная группа, но связи внутри неё до конца не понятны. К настоящему времени данные по анализу нуклеотидных последовательностей ряда генов из-за своей малочисленности пока не отражают полную картину в филогении бурых водорослей. Традиционно к наиболее примитивным бурым водорослям относили эктокарпусовые, но анализ последовательностей генов rbcL, psaA, psaB и их комбинации показывает, что они таковыми не являются. В деревьях, полученных в этих исследованиях, эктокарпусовые располагаются на вершине, а в основании — представители порядка Ishigeales, которые рано отделились от общего дерева бурых водорослей.
Не вызывает сомнений отнесение бурых водорослей к охрофитовым. В рамках этого отдела по ряду особенностей их долгое время считали наиболее близкими к золотистым водорослям. В настоящее время эта точка зрения оспаривается. По ультраструктурным, биохимическим особенностям и по сравнению нуклеотидных последовательностей гена 16S рРНК бурые водоросли ближе всего стоят к трибофициевым. После описания нового класса Schizocladiophyceae в ряде исследований показано, что он является сестринской группой для бурых водорослей.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом