
Водоросли относятся к низшим растениям, наиболее примитивным: у них отсутствует разделение организма на стебель, корень и листья. Спешу заметить, что термин «низшие растения» — отжившее понятие, использовавшееся в ботанике до второй половины XX века.
Современная биология не считает дифференциацию тканей определяющим различием, сейчас существенным считают фундаментальные различия в строение клеток, обмене веществ. Тем не менее, во многих устаревших пособиях этот термин используется, и я обязан предупредить вас о нем.
Наука о водорослях называется альгология (от лат. alga — морская трава, водоросль и греч. λόγος — учение).
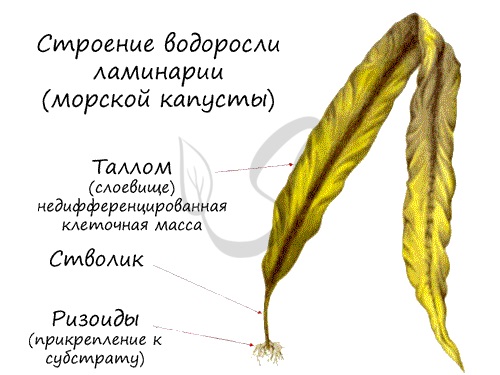
Среди водорослей есть одноклеточные и многоклеточные, некоторые водоросли достигают в длину 100-200 метров. Способ питания водорослей автотрофный: они синтезируют органические вещества в процессе фотосинтеза. Солнечный свет, проходя через толщу воды, рассеивается, что делает фотосинтез с увеличением глубины все труднее и труднее. Поэтому кроме хлорофилла они часто имеют и другие пигменты.
Клетки водорослей характеризуются наличием клеточной стенки (из целлюлозы и гликопротеинов — от греч. glykys сладкий (углеводы) + греч. prōtos — первый, важнейший (белок)) Органоиды располагаются в цитоплазме (син. — внеядерной протоплазме), где также располагается(-ются) один или несколько хроматофоров. Размножение происходит бесполым, вегетативным или половым путем.
Тело водорослей представлено слоевищем (син. — талломом) — недифференцированным скоплением клеток. С помощью ризоидов (от др.-греч. ῥίζα — корень и εἶδος — вид) водоросли прикрепляются к субстрату (камням, коралловым полипам), функцию всасывания ризоиды не выполняют. У водорослей отсутствуют настоящие ткани, механических тканей нет, так как таллом водоросли поддерживается (парит) в толще воды. Нет проводящих тканей: каждая клетка имеет доступ к воде напрямую, так что в клетку из окружающей воды поступает кислород, а в воду удаляется углекислый газ.
Хроматофор (от греч. chroma — цвет и phoros — несущий) — органелла в клетке водоросли, аналогичная хлоропласту и осуществляющая фотосинтез. Отличается от хлоропласта упрощенным строением, меньшим размером и иным составом хлорофилла. Внешне отличаются между собой по форме, хроматофор может быть: чашевидный, спиралевидный, в виде незамкнутых колец, цилиндрические, лентовидные, дисковидные. В хроматофорах находятся пигменты, которые придают окраску растению.
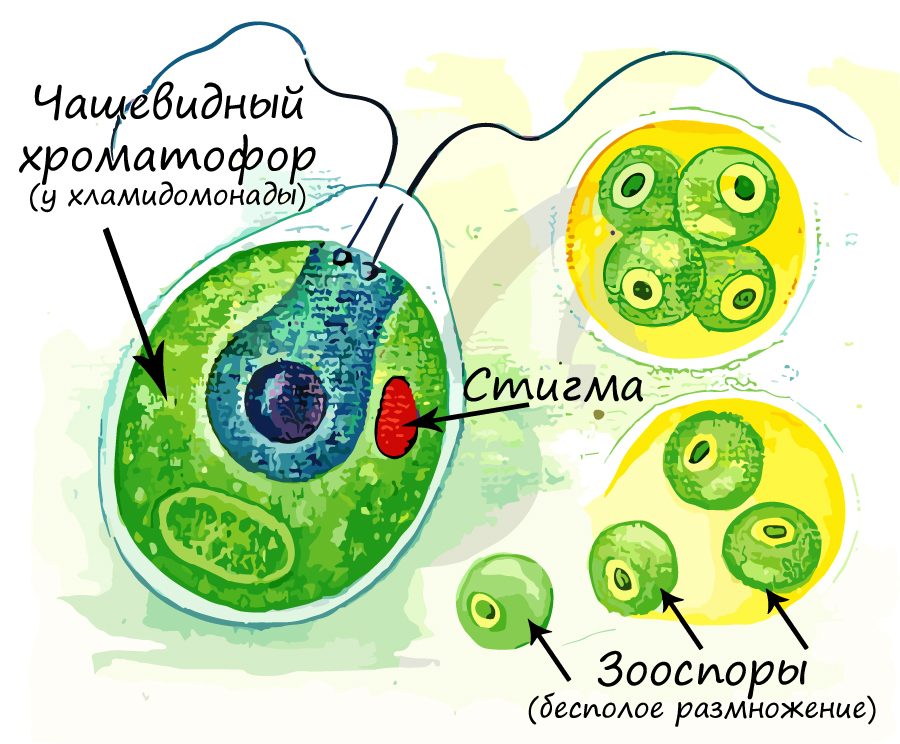
Система вакуолей в клетках водорослей развита отлично, в подвижных клетках водорослей можно обнаружить пульсирующие (сократительные) вакуоли. Их основная функция — поддержание постоянного осмотического давления внутри клетки. Вообразите: в глубине океана находится клетка водоросли, в которую постоянно поступает много воды. Если бы не было таких сократительных вакуолей, то клетка просто лопнула бы, но их работа обеспечивает удаление избытка воды.
Также у многих подвижных водорослей в клетках присутствует светочувствительный глазок (стигма), что обуславливает их чувствительность к свету — фототаксис. Подвижные водоросли стремятся занять как можно более освещенное место, чтобы активно шел процесс фотосинтеза.
Жизненный цикл водорослей
Жизненные циклы водорослей разнообразны, обусловлены рядом экологических факторов. Мы разберем жизненный цикл на примере зеленой водоросли ульвы (морского салата).
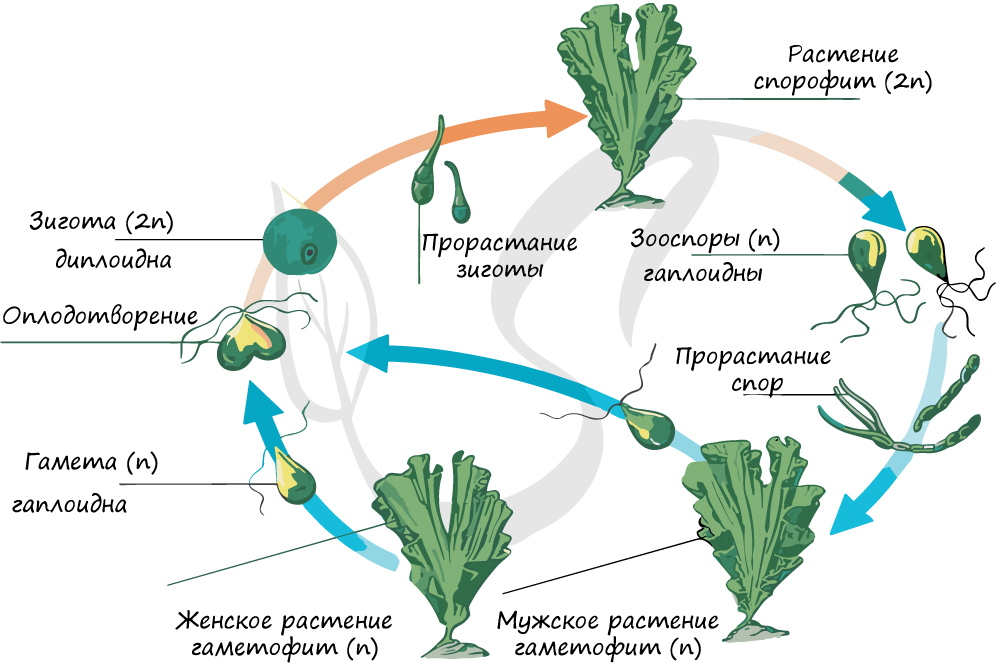
Для начала отметим, что в целом жизненный цикл водорослей представляет собой чередование двух фаз: гаплоидной (гаметофита) и диплоидной (спорофита). Гаплоидной фазой называется фаза, при которой клеточные ядра содержат непарный (половинный) набор хромосом. К гаплоидной фазе всегда принадлежат гаметы: сперматозоиды, спермии (отличающиеся от сперматозоидов отсутствием жгутика), яйцеклетки.
При слиянии двух гамет: яйцеклетки (n) и спермия (n) образуется зигота (2n) из которой развивается спорофит (2n), таким образом, в спорофите восстанавливается диплоидный набор хромосом. В зооспорангии на спорофите в результате мейоза образуются зооспоры (n), которые делятся митозом, порастают и образуют мужские и женские гаметофиты (n). Клетки гаметофитов делятся митозом, образуются гаметы (n), которые сливаются в зиготу (2n), цикл замыкается.
Типы половых процессов
У водорослей выделяют несколько типов полового процесса:
- Изогамия — копулирующие элементы (гаметы) не отличаются друг от друга, подвижны
- Анизогамия — от греч. anisos неравный и gamos брак (гетерогамия) — при таком типе копулирующие элементы различаются по размерам, форме, величине, поведению
- Оогамия — от др. греч. ᾠόν яйцо и γάμος брак — копулирующие элементы резко отличаются друг от друга: крупная женская гамета без жгутиков обычно с мужской мелкой подвижной гаметой. Допустимо считать оогамию в некотором смысле подтипом анизогамии.
Читайте также: Страна мастеров декупаж по ткани
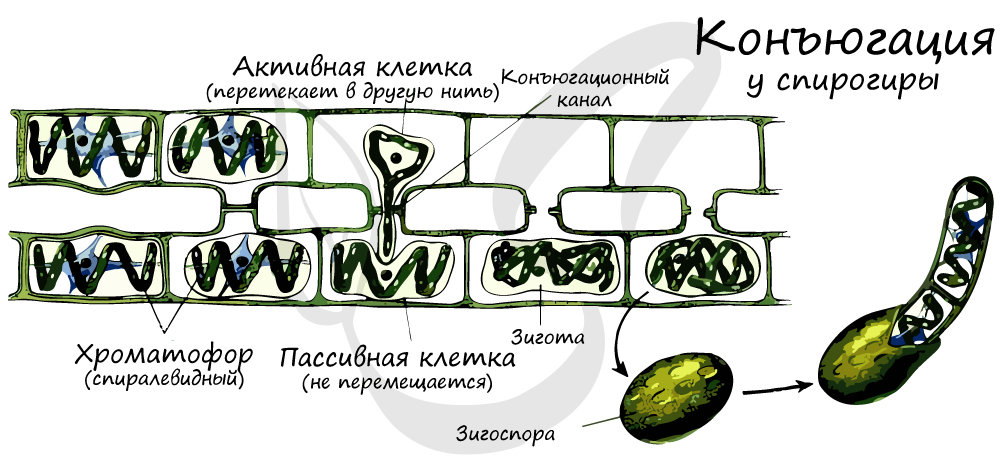
Особо стоит выделить тип полового процесса — конъюгацию. Конъюгация отличается тем, что сливаются не гаметы, а обычные вегетативные клетки, лишенные жгутиков. Клетки соединяются друг с другом с помощью боковых выростов, формируется копуляционный (конъюгационный) канал, по которому содержимое из одной клетки перетекает в другую — образуется зигоспора. В дальнейшем из зигоспоры развивается новая водоросль.
Отметим, что зооспора представляет собой подвижную клетку, которая способна двигаться в воде с помощью жгутиков. Образуется она в зооспорангии. Зооспора участвует в бесполом размножении у многих водорослей и простейших грибов. У некоторых водорослей имеются апланоспоры (гр. aplanes неподвижный + spora семя) — неподвижные безжгутиковые споры. Зооспоры и апланоспоры выходят в окружающую среду, разрывая стенки спорангия, в котором они находятся.
Значение водорослей
В Мировом океане водоросли составляют основную часть биомассы. Именно они являются главными продуцентами (производителями) органического вещества, преобразуя в ходе фотосинтеза энергию солнечного света в энергию химических связей. Значение водорослей для человека трудно переоценить: содержащиеся в них вещества необходимы для нормального роста и развития животных и человека (к примеру, морская капуста (ламинария) отличается большим содержанием йода.)

© Беллевич Юрий Сергеевич 2018-2022
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Водоросли, их отличие от высших растений. Основные типы морфологической структуры

Водоросли, их отличие от высших растений.
Водоросли – это прокариотические и эукариотические талломные споровые организмы, содержащие, как правило, в своих клетках хлорофилл, обладающие оксигенным фотосинтезом, являющиеся аэробами и не достигшие уровня дифференциации, характерной для архегониальных растений.
Возникли раньше (архейская эра,
Возникли позже (силур палеозойской эры).
Наземные и вторичноводные.
Произошли в разное время от разных предков. Распределены среди 4 царств (Сyanobiontes, Euglenobiontes, Chromista, Plantae).
Произошли от водорослевого предка и относятся к 1 царству Plantae.
Различные типы питания: автотрофное, гетеротрофное, миксотрофное (включая голозойное).
Преимущественно автотрофное питание, реже вторично гетеротрофное, либо авто-гетеротрофное.
Различный уровень организации (одноклеточный, колониальный, многоклеточный).
Только многоклеточный уровень организации.
Талломные (слоевищные) организмы.
Листостебельные, реже талломные, организмы.
Разнообразие морфоструктуры таллома (монадный, амебоидный, пальмеллоидный, коккоидный, нитчатый, разнонитчатый, паренхиматозный, псевдопаренхиматозный, харофитный, сифональный, сифонокладальный) (см. ниже).
У талломных только паренхиматозный (тканевой) таллом.
Ткани только у водорослей с паренхиматозным талломом (часть зеленых, бурых, красных, желтозеленых, золотистых).
Из тканей только паренхима, реже (бурые) покровная, образовательная, проводящая (“ситовидные трубки”), механическая.
У большинства отсутствуют механические структуры. Есть только у бурых в сердцевине). Вода – плотнее воздуха.
Имеются механические ткани.
У подавляющего большинства нет проводящих тканей.
Нет устьиц, газообмен всей поверхностью тела.
У большинства нет покровных тканей.
Всасывание минеральных веществ всей поверхностью тела. Ризоиды только для прикрепления.
Всасывание воды и минеральных веществ с помощью корневых волосков у большинства.
Покровы клеток разнообразны (цитоплазматическая мембрана, пелликула, перипласт, тека, панцирь, домик, клеточная оболочка).
Клетки имеют клеточную оболочку.
Клеточная стенка имеет разнообразный состав: целлюлоза, каллоза, муреин, пектин, углекислая известь, альгиновая кислота, фикоколлоиды. Лигнина нет.
Клеточная оболочка из целлюлозы, гемицеллюлозы, пектина. Могут откладываться лигнин, суберин.
Прокариоты и эукариоты. В клетке одно или несколько ядер.
Хроматофоры (хлоропласты) разнообразны по форме, числу, строению, набору пигментов, местоположению в клетке. Лейкопласты редко. У прокариотических водорослей нет хроматофоров.
Читайте также: Матрац из пористой резины обшитый тканью с электроподогревом
Линзовидные хлоропласты. Есть хромопласты, лейкопласты.
Тилакоиды в хроматофорах по одному, в ламеллах и реже в гранах (зеленые, харовые).
Тилакоиды только в гранах.
Каротиноиды (в том числе специфические – фукоксантин, сифонеин, сифоноксантин, вашериаксантин),
фикобилины (фикоцианин, аллофикоцианин, фикоэритрин).
Хроматическая адаптация в водоемах.
Пиреноид – это белковая структура, пространственно и функционально связанная с хроматофором, представляющая собой скопление фермента фиксации углекислоты при фотосинтезе.
Пиреноид есть только у некоторых мохообразных.
Разнообразные запасные вещества: крахмал, хризоламинарин (лейкозин), парамилон, ламинарин, багрянковый крахмал, сахариды, многоатомные спирты, цианофициновые гранулы, цианофициновый крахмал, белки, липиды (масла, жиры), волютин, гликоген.
Часто крахмал, масло, белки.
Кристы митохондрий разные: пластинчатые, трубчатые, дисковидные.
Кристы митохондрий только пластинчатые (ламеллярные).
Центриоли есть у немногих (хлорококковые зеленые, желтозеленые, бурые).
Жгутики у подвижных стадий (зооспор, гамет) разные: изоморфные, изоконтные, гетероморфные, гетероконтные.
Жгутики есть только у сперматозоидов (только изоморфные и изоконтные).
– вегетативное (деление клетки, фрагментация колонии и многоклеточного таллома, столоны, выводковые почки, гормогонии, акинеты, гонидии, кокки, планококки, клубеньки),
– собственно бесполое (подвижные и неподвижные споры),
– половое (хологамия, гаметогамия, конъюгация, автогамия). Изогамия, гетерогамия, оогамия истинная или примитивная.
Вегетативное (выводковые почки, вегетативные органы).
Собственно бесполое только неподвижными спорами.
Половое только гаметогамия – истинная оогамия, сифоногамия.
Споры гаплоидные и диплоидные.
Все споры одинаковые по размеру.
Есть одинаковые споры, но есть и разные споры – микроспоры и мегаспоры.
Гаметангии у большинства одноклеточные. Только у харовых исключение.
Гаметангии только многоклеточные.
Для копуляции гамет нужна капельно-жидкая влага.
Для копуляции гамет капельно-жидкая влага нужна только споровым. У семенных мужские гаметы доставляют гаусториальная или пыльцевая трубки.
Разнообразные (все из возможных) жизненные циклы и типы мейоза.
Есть циклы без полового процесса и мейоза.
Только один тип цикла и мейоза – гапло-диплобионтный со спорическим (промежуточным) мейозом.
В гапло-диплобионтном цикле есть
1) изоморфное чередование поколений,
2) гетероморфное с преобладанием гаметофита,
3) гетероморфное с преобладанием спорофита.
Только гетероморфное чередование поколений:
1) гетероморфное с преобладанием гаметофита (только у мохообразных),
3) гетероморфное с преобладанием спорофита (у всех других высших растений).
Уровни организации водорослей
По уровню организации водоросли делятся на одноклеточные, колониальные и многоклеточные.
Одноклеточные водоросли – представлены одиночными клетками (талломами) и присутствуют среди всех отделов, за исключением бурых и харовых водорослей. Например, Euglena, Chlamydomonas.
Колониальные водоросли представлены в таких отделах как синезеленые, золотистые, диатомовые, желтозеленые, красные и зеленые. Колония – более или менее сложное объединение талломов, которые при делении остаются соединенными, как правило, посредством слизи, выростов покровов тела, редко при помощи плазмодесм. Например, Microcystis, Scenedesmus, Volvox, Melosira. Ценобий – это колония, в которой число талломов определяется на ранних стадиях развития и не меняется до следующей репродуктивной фазы. Талломы в составе ценобия только растут, но не делятся. Например, Scenedesmus, Volvox.
Многоклеточные водоросли имеют таллом из многих клеток. Такие водоросли есть среди многих отделов, за исключением эвгленовых, криптофитовых и диатомовых водорослей. Например, Ulothrix, Laminaria.
Морфологическая дифференциация таллома водорослей
Огромное разнообразие внешней формы водорослей может быть сведено к нескольким основным структурам, или ступеням морфологической дифференциации. Эти структуры повторяются в разных систематических группах, что указывает на некоторый параллелизм в эволюции их форм.
Монадная структура. Наиболее характерным признаком является наличие жгутиков, с помощью которых организмы активно передвигаются в воде. У простейших одноклеточных и колониальных форм монадная структура наблюдается в течение всей вегетативной жизни индивидуума, а у более высокоорганизованных бывает лишь у репродуктивных клеток. Монадный тип структуры оказался эволюционно перспективным. На его основе развились другие, более сложные структуры, связанные с утратой подвижности в вегетативном состоянии. Например, Volvox, Euglena, Chlamydomonas.
Амебоидная, или ризоподиальная структура. Наиболее важные признаки этого типа структуры – отсутствие твердой оболочки и способность к амебоидному движению с помощью псевдоподий. Амебоидный тип таллома, по мнению многих ученых, является эволюционным тупиком, возникшим в результате приспособления монадных форм к особым условиям существования в биотопах, богатых органикой. Например, Rhizochloris.
Читайте также: Это нарушение целостности кости с повреждением окружающих тканей
Гемимонадная (пальмеллоидная, капсальная, тетраспоральная, трансгрессивная) структура. Характерным признаком этого типа структуры является объединение неподвижных клеток в слизистую колонию. Клетки в общей слизи колонии независимы друг от друга и часто имеют органеллы, свойственные монадным организмам (сократительные вакуоли, стигмы, жгутики или их производные). Клеткам гемимонадного типа, как и монадным, свойственно полярное строение. Если эта структура встречается в жизненном цикле водоросли как временное явление, ее называют пальмеллевидным состоянием. Гемимонадный тип таллома стал важным этапом на пути морфологической эволюции водорослей в направлении от подвижных монадных к типично растительным неподвижным формам. Например, Apiocystis.
Коккоидная структура. Неподвижный за счет жгутиков и псевдоподий таллом, не изменяющий форму тела, покрытый плотной клеточной оболочкой (целлюлоза, муреин в составе стенки) или панцирем. Например, Chlorella, Microcystis, Pinnularia. Клетки различной формы (не всегда шаровидные), одиночные или соединенные в колонии. В эволюционном плане этот тип структуры рассматривается как исходный для возникновения многоклеточных талломов. У диатомовых водорослей, имеющих на панцире шов, коккоидные талломы подвижные, но за счет особого механизма, связанного с циркуляцией слизи или воды в шве.
Нитчатая, или трихальная структура. Представляет талломы, состоящие из клеток, расположенных в форме нити. Нити могут быть простые (улотрикс) или разветвленные (бульбохете, эдокладиум), одно- или многорядные, свободноживущие или прикрепленные, одиночные или объединенные в разного типа соединения. Нитям присуще важнейшее свойство растительных организмов – неограниченный рост в течение вегетативной фазы жизненного цикла. Рост называют диффузным, если способностью делиться обладают все клетки нити. Деление клеток может происходит только на ограниченных участках слоевища, называемых зонами роста, или меристемальными зонами. В зависимости от положения зон роста различают интеркалярный, базальный и апикальный рост. При интеркалярном росте меристемальная зона находится в средней части нити, при базальном – у основания нити, при апикальном – рост осуществляется делением верхушечных клеток. Нижняя клетка нередко превращается в бесцветный ризоид или стопу, лишенный хлоропластов. Нитчатый тип таллома послужил отправным пунктом для развития других более сложных типов талломов. Например, Ulothrix, Spirogyra.
Разнонитчатая, или гетеротрихальная структура. Таллом данной структуры состоит из стелющейся по субстрату горизонтальной части и отходящей от нее вертикальной (трентеполия, стигеоклониум). Стелющиеся по субстрату нити выполняют главным образом функцию прикрепления, вертикальные, приподнимающиеся над субстратом нити – ассимиляционную функцию. Может наблюдаться редукция горизонтальной (Драпарнальдия) или вертикальной (Coleochaete) части таллома, реже и горизонтальной, и вертикальной частей (Desmococcus).
Паренхиматозная, или тканевая структура. Клетки первичной нити способны к делению в разных плоскостях. В пределах этого типа структуры можно наблюдать постепенное усложнение таллома от простых недифференцированных пластинок с диффузным ростом до сложноорганизованных слоевищ, с «тканями», выполняющими различные функции. Например, Ulva, Porphyra, Laminaria.
Псевдопаренхиматозная, или ложнотканевая структура. Характерным признаком этого типа структуры является образование достаточно крупных слоевищ в результате переплетения и срастания нитей разветвленного нитчатого (иногда сифонального) таллома. Образуются ложные ткани. Тупиковая ветвь в морфологической эволюции водорослей. Например, Batrachospermum, Nemalion.
Харофитная структура. Имеется только у харовых водорослей. Характеризуется крупными многоклеточными талломами линейно-членистого строения, которые состоят из главных побегов с ветвями («стеблей»), боковых ветвей ограниченного роста («листьев») и ризоидов. Например, Chara, Nitella.
Сифональная структура. Отличительная черта такой структуры – отсутствие внутри таллома клеточных перегородок при наличии большого числа ядер. Талломы могут быть сложно расчленены и иметь довольно крупные размеры. Например, Codium, Caulerpa, Vaucheria.
Сифонокладальная структура. Представлена многоядерными клетками, объединенными в многоклеточные талломы разной формы. Берет свое начало от сифональной структуры. Имеет место сегрегативное деление, характерной особенностью которого является разобщенность процессов кариокинеза и цитокинеза. Например, Cladophora, Valonia.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом