
Когда выше мы давали определение клетки, то в качестве одного из ее свойств мы называли способность к делению с образованием копии себе подобной. Хотя это остается справедливым для клеток одноклеточных организмов, уровень специализации, которого достигли клетки у многоклеточных, требует расширения базовых понятий.
Термин дифференцировка описывает процесс, посредством которого клетка приобретает новый фенотип, или же дает начало предшественникам клеток, которые по фенотипу отличаются от родительской.
Под развитием организма подразумевают процесс, в результате которого исходная клетка (оплодотворенный ооцит в случае млекопитающих) путем последовательных делений превращается в организм, состоящий из клеток различных типов. Оплодотворенная зигота обладает тотипотентностью, т. е. из нее способны образовываться все клетки организма.
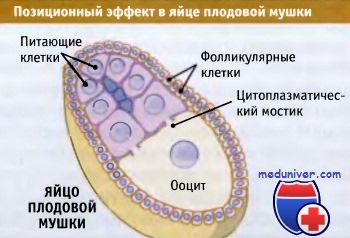
Питающие клетки окружают ооцит и участвуют в переносе веществ,
что создает асимметричность в распределении белков.
Процесс развития можно рассматривать как наложение неких дополнительных ограничений на свойства образующихся клеток. В результате этих ограничений клетки могут развиваться, образуя лишь определенный тип клеток.
Поскольку существуют многоклеточные организмы, состоящие из специализированных тканей, необходимо различать два типа клеток. К соматическим относятся все клетки организма, за исключением герминативных. Соматические клетки предназначены для выполнения в организме определенных функций и не участвуют в процессе воспроизводства организма. У млекопитающих они являются диплоидными клетками (т. е. содержат один материнский и один отцовский набор генетического материала).
Следующее поколение организма формируется при участии специализированных герминативных клеток. Эти клетки существуют для каждого пола отдельно и являются гаплоидными: сперматозоиды и ооциты содержат по одному набору генетического материала.
В период развития и существования взрослого организма некоторые клетки сохраняют способность к воспроизводству любых клеток различных тканей, в то время как другие образуют лишь родственные клетки или не образуют никаких. Клетки, обладающие широкими возможностями к регенерации, называются стволовыми. Например, клетки иммунной системы произошли от иммунных стволовых клеток. Основной вопрос биологии состоит в том, существуют стволовые клетки только при развитии организма или же они сохраняются и в зрелом возрасте.
При развитии ткани происходит серия процессов дифференцировки клеток, при которых они продолжают изменять свой фенотип до тех пор, пока не образуются все необходимые типы. По завершении этого процесса некоторые клетки могут утратить способность к делению и превратиться в терминально дифференцированные. Клетки также могут стареть и умирать. Многие соматические клетки являются терминально дифференцированными. К их числу относятся и половые клетки, хотя они обладают уникальной способностью образовывать зиготу, которая способна к делению.
Терминально дифференцированные клетки могут проявлять настолько необычные свойства, что возникает вопрос, применимо ли к ним определение самого понятия клетка. У млекопитающих дифференцировка эритроцитов приводит к потере ядра. Клетка представляет собой не более чем мембрану, которая внутри содержит раствор гемоглобина и форма которой поддерживается сетью волокон актина и спектрина. Поскольку эритроцит произошел из полностью функциональной клетки, мы рассматриваем его как клетку, хотя и утратившую почти все характерные признаки.
Широко обсуждается вопрос, происходят ли при дифференцировке клеток постоянные изменения их генетической программы или же ограничения на развитие фенотипа носят исключительно эпигенетическую природу? Возможность клонирования организмов при помещении ядра соматической клетки в ооцит дает однозначный ответ на этот вопрос: почти каждая клетка организма сохраняет генетическую информацию, необходимую для поддержания процесса развития. (Исключение составляют клетки с измененным генетическим материалом, например клетки, продуцирующие антитела, или утратившие генетическую информацию (эритроциты).)
Читайте также: Гладкая мышечная ткань гистология презентация
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Дифференцировка тканей у человека
Нарушения тканевой дифференцировки Б. Фишер-Вазельс (1927) разделил на 3 основные группы 1) врожденные пороки развития, гамартин и гамартомы, 2) метаплазии и 3) дисплазии Аналогичная классификация, хотя и с иной терминологией, была предложена Е. Finckh в 1982 г, выделившим аберрантную, вариантную и девиантную формы нарушения дифференцировкн в условиях патологии.
К первой форме относятся врожденные уродства, гетеротопии, гетероплазии, гамартомы. Гетеротопиями именуют смещение в перинатальном периоде какого-либо органа или его части с дальнейшим развитием в необычных местах, например ткани щитовидной железы в корне языка, коры надпочечника внутри почки и т. д. Гетероплазии служат проявлением первичного нарушения дифференцировкн в развивающейся ткани слизистая оболочка желудка в меккелевом дивертикуле, островки хряща в гипопластической почке. Гетероплазию необходимо отличать от метаплазии, так как последняя возникает у взрослых организмов в участках зрелой высокодифференцированной ткани. Гамартомы или хористомы представляют собой опухолеподобные узлы или доброкачественные опухоли, развивающиеся из гетеротопированной ткани.
В редких случаях в эктопированиых тканях развиваются истинные злокачественные опухоли аденокарцинома, инсулома из ткани поджелудочной железы в стенке желудка нли кишечника, фиброаденома, а затем и рак из ткаии молочной железы, эктопированной в области вульвы и др.
Ко второй форме нарушений дифференцировки относят метаплазии, заключающиеся в замещении одного типа дифференцированных (зрелых) клеточных элементов другим вследствие хронического воспаления, нарушений питания, нарушений функции.
При оценке метапластических изменений следует учитывать не только гистотипические и структурные характеристики, но и цитологические особенности тканевых элементов.
Имеется широкий спектр признаков, по которым можно установить качественные различия между отдельными типами дифференцированных форм это различия во внутриклеточных органеллах, продуктах клеточного выделения, эндо- и экзокринной секреции, ферментном обмене, митотической активности, специфической чувствительности к гормонам, токсинам, лекарственным веществам, в различном характере межклеточных контактов, двигательной и сократительной активности и т. п.
Третья группа нарушений дифференцировки характеризуется появлением клеток с патологическими характеристиками и обозначается как дисплазия.
Указанные варианты нарушений тканевой дифференцировки могут развиваться независимо друг от друга. Чрезвычайно важно, что диспластические изменения нередко возникают на фоне метаплазии (дисплазия на фоке кишеч ной метаплазии слизистой оболочки желудка, дисплазия шейки матки при эпидермизации желез н пролиферации резервных клеток и т. д., в аденомах, в частности желудка, кишечника, молочной железы, предстательной железы).
По-видимому, в широком общепатологическом смысле дисплазия, как нарушение дифференцировки ткани должна включать в себя не только весь спектр предраковых изменений, но также и предынвазивный и инвазивный рак.
Дисплазия, будучи тесно связанной с хроническим воспалением или раздражением, включая механические, химические, эндокринные воздействия, нередко возникает на фоне гиперпластических, дисрегенераторных и метапластических процессов. При этом морфологический дифференциальный диагноз этих состояний, как указывалось, бывает крайне затруднителен. Между тем правильная оценка наличия дисплазии и степень ее развития имеет не только теоретическое, но н важное клиническое значение, определяя в известной мере и степень риска последующей малигнизации.
Читайте также: В кроветворении участвует ткань
По-видимому, следует отказаться от использвания терминов «атипическая эпителиальная гиперплазия», «аденоматозная гиперплазия», «атипическая регенерация» и «атипическая метаплазия» даже как синонимов диспластических изменений, так как эти термины создают возможность неверной клинической оценки процесса.
В настоящее время важное место в онкологии занимает теория «опухолевого поля», объясняющая закономерности мультицентрического роста опухолей и, соответственно, очагов диспластических изменений и внутриэпителиального рака. Опухолевое поле является основой формирования множественных зачатков новообразований, находящихся на разных стадиях канцерогенеза и часто происходящих из разных клеточных клонов. При слиянии этих зачатков в единый очаг нередко возникают диморфные и полиморфные гистологические типы опухолей.
Вместе с тем нельзя не подчеркнуть, что результаты исследований последних лет убеждают в неоднозначности морфологической характеристики опухолевого поля оно может включать в себя небольшой участок ткани, вплоть до микроочага, состоящего, однако, из мультицентричных зачатков (чаще в мезенхимальиой ткани), а в эпителиальной ткаии локализуется на обширных участках, нередко захватывая целый орган или несколько органов (например, молочные железы, желудок, бронхиальное дерево).
Предраковые (диспластические) изменения приводят к инвазивному раку в результате прерывистости процесса опухолевой прогрессии, в ходе которой возникают качественные сдвиги в состоянии клеток Указанные изменения, по Л. Фулдсу, являются «несовершенным раком», так как не обладают всеми признаками злокачественного новообразования. Малигнизация представляет собой одни из последних этапов опухолевого процесса Когда и на каком этапе предраковых состояний возникает необратимая опухолевая прогрессия, до сих пор остается неясным. Однако некоторые гистологические признаки катаплазии (анаплазии), как это было отмечено, наблюдаются уже при дисплазии I степени. Нет сомнений, что идеи Л Фулдса верны, однако в клинической онкологии приложимы не ко всем вариантам и локализациям новообразований.
Факты постоянного и постепенного нарастания предраковых (диспластических) изменений в известной мере противоречат возможности внезапной скачкообразно возникающей малнгинзации. Канцерогенные агенты, как теперь стало известно, воздействуют иа клеточные мембраны и одним из первых признаков трансформации нормальных клеток в опухолевые является их обособление, утрата контактного торможения, что способствует постепенному нарастанию потенций к инфильтативному росту и метастазированию.
Дифференцировка тканей у человека
Известно, что одни клетки непрерывно делятся, например стволовые клетки костного мозга, клетки зернистого слоя эпидермиса, эпителиальные клетки слизистой кишечника; другие, включая гладкомышечные, могут не делиться в течение нескольких лет, а некоторые клетки, например нейроны и поперечнополосатые мышечные волокна, вообще не способны делиться (если не считать внутриутробный период).
В некоторых тканях дефицит клеточной массы устраняется за счет быстрого деления оставшихся клеток. Так, у некоторых животных после хирургического удаления 7/8 печени ее масса восстанавливается почти до исходного уровня за счет деления клеток оставшейся 1/8 части. Таким свойством обладают многие железистые клетки и большинство клеток костного мозга, подкожной клетчатки, кишечного эпителия и других тканей, за исключением высокодифференцированных мышечных и нервных клеток.
Пока мало известно, каким образом в организме поддерживается необходимое число клеток разных типов. Тем не менее, экспериментальные данные говорят о существовании трех механизмов регуляции клеточного роста.
Во-первых, деление многих видов клеток находится под контролем факторов роста, вырабатываемых другими клетками. Некоторые из этих факторов поступают к клеткам из крови, другие — из близлежащих тканей. Так, эпителиальные клетки некоторых желез, например поджелудочной, не могут делиться без фактора роста, вырабатываемого подлежащей соединительной тканью.
Читайте также: Кукольный театр репка своими руками из ткани
Во-вторых, большинство нормальных клеток перестают делиться при недостатке места для новых клеток. Это можно наблюдать в клеточных культурах, в которых клетки делятся, пока не начнут контактировать друг с другом, затем они прекращают деление.
В-третьих, многие тканевые культуры перестают расти, если в культуральную жидкость попадает даже небольшое количество вырабатываемых ими веществ. Все эти механизмы контроля клеточного роста можно рассматривать как варианты механизма отрицательной обратной связи.
Регуляция размера клеток. Размер клетки зависит в основном от количества функционирующей ДНК. Так, при отсутствии репликации ДНК клетка растет, пока не достигнет определенного объема, после этого ее рост прекращается. Если с помощью колхицина заблокировать процесс образования веретена деления, то можно остановить митоз, хотя репликация ДНК при этом будет продолжаться. Это приведет к тому, что количество ДНК в ядре значительно превысит норму, и объем клетки увеличится. Предполагается, что избыточный рост клеток в данном случае обусловлен повышенной продукцией РНК и белка.
Дифференциация клеток в тканях
Одной из характеристик роста и деления клеток является их дифференцировка, под которой понимают изменение их физических и функциональных свойств в ходе эмбриогенеза с целью образования специализированных органов и тканей организма. Рассмотрим интересный эксперимент, помогающий объяснить этот процесс.
Если из яйцеклетки лягушки с помощью специальной методики вынуть ядро и вместо него поместить ядро клетки слизистой кишечника, то из такой яйцеклетки может вырасти нормальная лягушка. Этот эксперимент показывает, что даже такие высокодифференцированные клетки, как клетки слизистой кишечника, содержат всю необходимую генетическую информацию для развития нормального организма лягушки.
Из эксперимента ясно, что дифференцировка идет не за счет потери генов, а благодаря селективной репрессии оперонов. Действительно, на электронных микрофотографиях можно увидеть, что некоторые сегменты ДНК, «упакованные» вокруг гистонов, конденсированы настолько сильно, что уже не могут быть расплетены и использованы в качестве матрицы для транскрипции РНК. Этому явлению можно дать такое объяснение: на определенной стадии дифференцировки клеточный геном начинает синтезировать белки-регуляторы, которые необратимо репрессируют определенные группы генов, поэтому эти гены навсегда остаются инактивированными. Как бы то ни было, зрелые клетки человеческого организма способны синтезировать всего 8000-10000 разных белков, хотя если бы функционировали все гены, эта цифра составила бы около 30000.
Эксперименты на эмбрионах показывают, что некоторые клетки способны осуществлять контроль над дифференцировкой соседних клеток. Так, хордомезодерму называют первичным организатором эмбриона, поскольку вокруг нее начинают дифференцироваться все остальные ткани эмбриона. Превращаясь в ходе дифференцировки в сегментированную, состоящую из сомитов дорсальную мезодерму, хордомезодерма становится индуктором для окружающих тканей, запускающим формирование из них практически всех органов.
В качестве другого примера индукции можно привести развитие хрусталика. Когда глазной пузырек соприкасается с головной эктодермой, она начинает утолщаться, постепенно превращаясь в хрусталиковую плакоду, а та, в свою очередь, образует впячивание, из которого в результате и формируется хрусталик. Таким образом, развитие эмбриона в значительной степени обусловлено индукцией, суть которой заключается в том, что одна часть эмбриона вызывает дифференцировку другой, а та — дифференцировку остальных частей.
Итак, хотя дифференцировка клеток в целом все еще остается для нас загадкой, многие регуляторные механизмы, которые лежат в ее основе, нам уже известны.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом