
Кровообращение выполняет одну из важнейших функций переноса кислорода от легких к тканям, а углекислого газа — от тканей к легким. Потребление кислорода клетками тканей может изменяться в значительных пределах, например при переходе от состояния покоя к физической нагрузке и наоборот. В связи с этим кровь должна обладать большими резервами, необходимыми для увеличения ее способности переносить кислород от легких к тканям, а углекислый газ в обратном направлении.
Транспорт кислорода.
При 37 С растворимость 02 в жидкости составляет 0,225 мл • л-1 • кПа-1 (0,03 мл/л/мм рт. ст.). В условиях нормального парциального давления кислорода в альвеолярном воздухе, т. е. 13,3 кПа или 100 мм рт.ст., 1 л плазмы крови может переносить только 3 мл 02, что недостаточно для жизнедеятельности организма в целом. В покое в организме человека за минуту потребляется примерно 250 мл кислорода. Чтобы тканям получить такое количество кислорода в физически растворенном состоянии, сердце должно перекачивать за минуту огромное количество крови. В эволюции живых существ проблема транспорта кислорода была более эффективно решена за счет обратимой химической реакции с гемоглобином эритроцитов. Кислород переносится кровью от легких к тканям организма молекулами гемоглобина, которые содержатся в эритроцитах.
Гемоглобин способен захватывать кислород из альвеолярного воздуха (соединение называется ок-сигемоглобином) и освобождать необходимое количество кислорода в тканях. Особенностью химической реакции кислорода с гемоглобином является то, что количество связанного кислорода ограничено количеством молекул гемоглобина в эритроцитах крови. Молекула гемоглобина имеет 4 места связывания с кислородом, которые взаимодействуют таким образом, что зависимость между парциальным давлением кислорода и количеством переносимого кислорода с кровью имеет S-образную форму, которая носит название кривой насыщения или диссоциации оксигемоглобина (рис. 10.18). При парциальном давлении кислорода 10 мм рт. ст. насыщение гемоглобина кислородом составляет примерно 10 %, а при Р02 30 мм рт. ст. — 50—60 %. При дальнейшем увеличении парциального давления кислорода от 40 мм рт. ст. до 60 мм рт. ст. происходит уменьшение крутизны кривой диссоциации оксигемоглобина и процент его насыщения кислородом возрастает в диапазоне от 70—75 до 90 % соответственно. Затем кривая диссоциации оксигемоглобина начинает занимать практически горизонтальное положение, поскольку увеличение парциального давления кислорода с 60 до 80 мм рт. ст. вызывает прирост насыщения гемоглобина кислородом на 6 %. В диапазоне от 80 до 100 мм рт. ст. процент образования оксигемоглобина составляет порядка 2. В результате кривая диссоциации оксигемоглобина переходит в горизонтальную линию и процент насыщения гемоглобина кислородом достигает предела, т. е. 100. Насыщение гемоглобина кислородом под влиянием Р02 характеризует своеобразный молекулярный «аппетит» этого соединения к кислороду.
Значительная крутизна кривой насыщения гемоглобина кислородом в диапазоне парциального давления от 20 до 40 мм рт. ст. способствует тому, что в ткани организма значительное количество кислорода может диффундировать из крови в условиях фадиента его парциального давления между кровью и клетками тканей (не менее 20 мм рт. ст.). Незначительный процент насыщения гемоглобина кислородом в диапазоне его парциального давления от 80 до 100 мм рт. ст. способствует тому, что человек без риска снижения насыщения артериальной крови кислородом может перемещаться в диапазоне высот над уровнем моря до 2000 м.
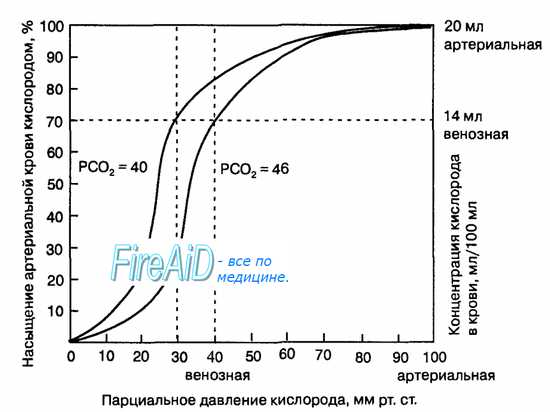
Рис. 10.18. Кривая диссоциации оксигемоглобина. Пределы колебания кривой при РС02 = 40 мм рт. ст. (артериальная кровь) и РС02 = 46 мм рт. ст. (венозная кровь) показывают изменение сродства гемоглобина к кислороду (эффект Ходена).
Общие запасы кислорода в организме обусловлены его количеством, находящимся в связанном состоянии с ионами Fe2+ в составе органических молекул гемоглобина эритроцитов и миоглобина мышечных клеток.
Один грамм гемоглобина связывает 1,34 мл 02. Поэтому в норме при концентрации гемоглобина 150 г/л каждые 100 мл крови могут переносить 20,0 мл 02.
Количество 02, которое может связаться с гемоглобином эритроцитов крови при насыщении 100 % его количества, называется кислородной емкостью гемоглобина. Другим показателем дыхательной функции крови является содержание 02 в крови (кислородная емкость крови), которое отражает его истинное количество, как связанного с гемоглобином, так и физически растворенного в плазме. Поскольку в норме артериальная кровь насыщена кислородом на 97 %, то в 100 мл артериальной крови содержится примерно 19,4 мл 02.
Диффузия со2 из ткани в кровь
Диффузия газов через альвеолярную мембрану происходит между альвеолярным воздухом и венозной, а также артериальной кровью легочных капилляров. В табл. 10.2 приведены стандартные величины напряжения дыхательных газов в артериальной и венозной крови легочных капилляров.

Таблица 10.2. Напряжение дыхательных газов в артериальной и венозной крови легочных капилляров
Градиенты парциального давления кислорода и углекислого газа обусловливают процесс пассивной диффузии через альвеолярную мембрану кислорода из альвеол в венозную кровь (градиент 60 мм рт. ст.), а углекислого газа — из венозной крови в альвеолы (градиент 6 мм рт. ст.). Парциальное давление азота по обе стороны альвеолярной мембраны остается постоянным, поскольку этот газ не потребляется и не продуцируется тканями организма. При этом сумма парциального давления всех газов, растворенных в тканях организма, меньше, чем величина атмосферного давления, благодаря чему газы в тканях не находятся в газообразной форме. Если величина атмосферного давления будет меньше, чем парциальное давление газов в тканях и в крови, то газы начинают выделяться из крови в виде пузырьков, вызывая тяжелые нарушения в кровоснабжении тканей организма (кессонная болезнь).
Читайте также: Какие нити располагаются вдоль полотна ткани нити утка нити основы
Скорость диффузии 02 и С02 в легких
Скорость диффузии (M/t) кислорода и углекислого газа через альвеолярную мембрану количественно характеризуется законом диффузии Фика. Согласно этому закону газообмен (M/t) в легких прямо пропорционален градиенту (ДР) концентрации 02 и С02 по обе стороны от альвеолярной мембраны, площади ее поверхности (S), коэффициентам (к) растворимости 02 и С02 в биологических средах альвеолярной мембраны и обратно пропорционален толщине альвеолярной мембраны (L), а также молекулярной массе газов (М). Формула этой зависимости имеет следующий вид:
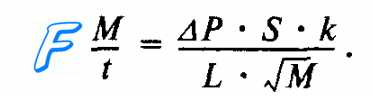
Структура легких образует максимальное по величине поле для диффузии газов через альвеолярную стенку, которая имеет минимальную толщину (рис. 10.16). Так, количество альвеол в одном легком человека приблизительно равно 300 млн. Суммарная площадь альвеолярной мембраны, через которую происходит обмен газов между альвеолярным воздухом и венозной кровью, имеет огромные размеры (порядка 100 м2), а толщина альвеолярной мембраны составляет лишь — 0,3—2,0 мкм.
В обычных условиях диффузия газов через альвеолярную мембрану происходит в течение очень короткого отрезка времени (не более 3/4 с), пока кровь проходит через капилляры легких. Даже при физической работе, когда эритроциты проходят капилляры легкого в среднем за 1/4 с, указанные выше структурные особенности альвеолярной мембраны создают оптимальные условия для формирования равновесия парциальных давлений 02 и С02 между альвеолярным воздухом и кровью капилляров легких (рис. 10.17). В уравнении Фика константы диффузии (к) пропорциональны растворимости газа в альвеолярной мембране. Углекислый газ имеет примерно в 20 раз большую растворимость в альвеолярной мембране, чем кислород. Поэтому, несмотря на существенное различие в градиентах парциальных давлений 02 и С02 по обе стороны от альвеолярной мембраны, диффузия этих газов совершается за очень короткий отрезок времени движения эритроцитов крови через легочные капилляры.
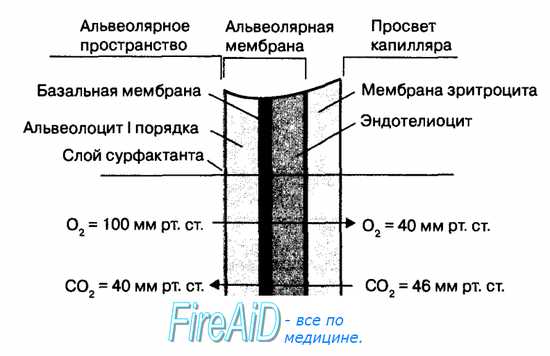
Рис. 10.16. Диффузия газов через альвеолярную мембрану. Диффузия газов в легких осуществляется по градиентам концентрации 02 и С02 между альвеолярным пространством и кровью капилляров легких, которые разделены альвеолярной мембраной. При этом диффузия тем эффективнее, чем тоньше альвеолярная мембрана и области контакта альвеолоцитов и эндотелиоцитов. Поэтому альвеолярная мембрана образована уплощенными частями альвеолоцитов I порядка (0,2 мкм) и эндотелиоцитов капилляров легких (0, 2 мкм), между которыми находится тонкая общая базальная мембрана (0,1 мкм) этих клеток. В состав мембраны входит также мономолекулярный слой сурфактант а. Мембрана эритроцитов является препятствием для диффузии газов в легких.
Газообмен через альвеолярную мембрану количественно оценивается диффузионной способностью легких, которая измеряется количеством газа (мл), проходящего через эту мембрану за 1 мин при разнице давления газа по обе стороны мембраны в 1 мм рт. ст.
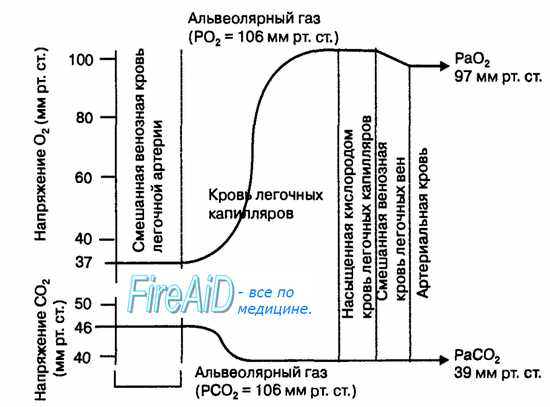
Рис. 10.17. Градиенты парциального давления дыхательных газов в смешанной венозной крови легочной артерии, альвеолярном воздухе и артериальной крови. Равновесие парциальных давлений углекислого газа и кислорода между альвеолярным воздухом и кровью легочных капилляров достигается в течение короткого времени (1/4—3/4 с) движения плазмы крови и эритроцитов в капиллярах легких.
Наибольшее сопротивление диффузии 02 в легких создают альвеолярная мембрана и мембрана эритроцитов, в меньшей степени — плазма крови в капиллярах. У взрослого человека в покое диффузионная способность легких 02 равна 20—25 мл • мин-1 • мм рт. ст.-1. С02, как полярная молекула (0=С=0), диффундирует через указанные мембраны чрезвычайно быстро, благодаря высокой растворимости этого газа в альвеолярной мембране Диффузионная способность легких С02 равна 400—450 мл•мин-1• мм рт. ст.-1.
Диффузия со2 из ткани в кровь
После поступления свежего воздуха в альвеолы начинается следующий этап дыхательного процесса: диффузия кислорода из альвеол в кровь и диффузия двуокиси углерода в обратном направлении — из крови в альвеолы. Процесс диффузии представляет собой беспорядочное движение молекул, прокладывающих себе дорогу через дыхательную мембрану и жидкости во всех направлениях. Однако в физиологии дыхания нас интересуют не только основные механизмы диффузии, но и ее скорость, что представляет собой намного более сложную проблему и потребует более глубоких знаний в области физики диффузии и обмена газов.
Физические основы диффузии и парциальные давления газов
Все газы, представляющие интерес для физиологии дыхания, являются простыми молекулами, которые свободно перемещаются в смеси. Этот процесс называют диффузией. Это справедливо и для газов, растворенных в жидкостях и тканях тела.
Читайте также: Рапс ткань что это
Для процесса диффузии необходимо наличие источника энергии. Энергия производится кинетическим движением самих молекул. При температуре выше абсолютного нуля молекулы находятся в постоянном движении. Это значит, что свободные молекулы, не связанные с другими молекулами, двигаются линейно на высокой скорости до встречи с другими молекулами. После столкновения их движение получит новое направление — до следующего столкновения. Таким образом, молекулы находятся в быстром и случайном движении среди себе подобных.
а) Диффузия газа одном направлении. Влияние градиента концентрации. Если в емкости или в растворе концентрация одного газа в одной зоне высокая, а в другой — низкая (для облегчения понимания просим вас изучить рисунок ниже), то суммарная диффузия газа будет направлена от зоны с высокой концентрацией в зону с низкой концентрацией: на рисунке в зоне А находится больше молекул, способных двигаться в направлении зоны Б, чем молекул, которые могут переместиться в обратном направлении, поэтому диффузия в каждом из направлений пропорциональна концентрации молекул, что на рисунке демонстрирует длина стрелок.
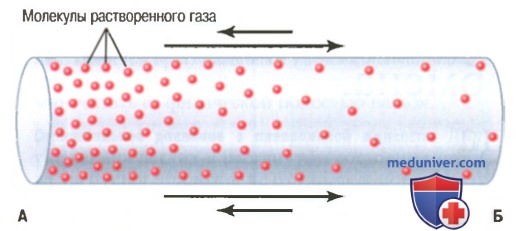
Диффузия кислорода из одной зоны (А) в другую (Б). Разница в длине стрелок представляет величину конечной диффузии
б) Давление газов в газовой смеси. Парциальные давления отдельных газов. Давление создается множественными ударами движущихся молекул о поверхность, поэтому давление газа на поверхности дыхательных ходов и альвеол пропорционально суммарной силе ударов о поверхность всех молекул данного газа в данный момент, т.е. давление газа прямо пропорционально концентрации молекул газа.
В физиологии дыхания мы имеем дело со смесями газов, состоящих главным образом из кислорода, азота и двуокиси углерода. Скорость диффузии каждого из них прямо пропорциональна давлению, создаваемому только этим газом, и это давление называют парциальным давлением данного газа. Далее приводим объяснение концепции парциального давления.
Воздух состоит примерно из 79% азота и 21% кислорода. Общее давление этой смеси на уровне моря равно 760 мм рт. ст. Из приведенного ранее объяснения молекулярных основ возникновения давления ясно, что доля каждого газа в давлении их смеси находится в прямой пропорции с его концентрацией, поэтому 79% из 760 мм рт. ст. давления воздуха создается азотом (600 мм рт. ст.) и 21% — кислородом (160 мм рт. ст.). Таким образом, парциальное давление азота в смеси составляет 600 мм рт. ст., парциальное давление кислорода — 160 мм рт.ст., а общее давление (760 мм рт. ст.) является суммой отдельных парциальных давлений. Парциальное давление отдельных газов обозначают PCO2, PO2, PN2, PH2O, PHe и т.д.
Видео физиология газообмена в легких и транспорта газов кровью — профессор, д.м.н. П.Е. Умрюхин
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Диффузия со2 из ткани в кровь
а) Транспорт кислорода артериальной кровью. Около 98% объема крови, входящей в левое предсердие после прохождения легких, прошло через легочные капилляры и было оксигенировано до PO2, равного приблизительно 104 мм рт. ст. Оставшиеся 2% объема крови ушли из аорты в бронхиальное кровообращение, которое снабжает в основном глубокие ткани легких, и не вошли в контакт с альвеолярным воздухом. Эту часть кровотока в легких называют «шунтирующей», т.к. она направляется мимо газообменных участков легких.
На выходе из легких напряжение кислорода шунтированной крови находится примерно на уровне нормальной системной венозной крови — около 40 мм рт. ст. При смешивании этой крови в легочных венах с оксигенированной кровью из альвеолярных капилляров эта так называемая венозная примесь крови приведет к снижению PO2, входящей в левую половину сердца и выбрасываемой оттуда в аорту крови, примерно до 95 мм рт. ст. Эти изменения PO2 в крови в разных точках системы кровообращения показаны на рисунке ниже.
![]() Изменения PO2 в капиллярной крови легких, системной артериальной крови и системной капиллярной крови, демонстрирующие эффект «венозной примеси» (смесь с легочной шунтовой кровью)
Изменения PO2 в капиллярной крови легких, системной артериальной крови и системной капиллярной крови, демонстрирующие эффект «венозной примеси» (смесь с легочной шунтовой кровью) ![]() Диффузия кислорода из тканевого капилляра к клеткам (PO2 в интерстициальной жидкости — 40 мм рт. ст., в клетках ткани — 23 мм рт. ст.)
Диффузия кислорода из тканевого капилляра к клеткам (PO2 в интерстициальной жидкости — 40 мм рт. ст., в клетках ткани — 23 мм рт. ст.)
б) Диффузия кислорода из периферических капилляров в тканевую жидкость. При достижении артериальной кровью периферических тканей PO2 в ней находится на уровне 95 мм рт. ст. На рисунке выше видно, что в окружающей тканевые клетки интерстициальной жидкости PO2 достигает только 40 мм рт. ст. Таким образом, имеется большой градиент давления, под влиянием которого кислород начинает быструю диффузию из капиллярной крови в ткани — настолько быструю, что PO2 в капиллярах почти сравняется с PO2 (40 мм рт. ст.) в интер-стиции, поэтому PO2 в крови, покидающей капилляры и поступающей в системные вены, составляет около 40 мм рт. ст.
1. Влияние скорости кровотока на PO2 интерстициальной жидкости. Если кровоток через какую-нибудь ткань повышается, в ткань попадает большее количество кислорода, и PO2 в ткани, соответственно, повышается. Обратите внимание, что повышение кровотока до 400% относительно кровотока в покое повышает PO2 от 40 мм рт. ст. до 66 мм рт. ст.. Однако высшей границей для подъема PO2 даже при максимальном кровотоке является 95 мм рт. ст., т.к. это является парциальным давлением кислорода в артериальной крови. Соответственно, при понижении кровотока в ткани снижается и PO2 в ткани.
Читайте также: Лучшие ткани в америке
![]()
Влияние кровотока и скорости потребления кислорода на PO2 в ткани
2. Влияние интенсивности тканевого метаболизма на PO2 интерстициальной жидкости. Если клеткам для метаболизма потребуется больше кислорода, чем обычно, то PO2 в интерстициальной жидкости снижается. Это влияние также видно на рисунке выше, где при повышении потребления кислорода клетками прослеживается снижение PO2 в интерстициальной жидкости и повышение его при снижении потребления.
В итоге величина PO2 ткани определяется равновесием между:
(1) скоростью транспорта кислорода кровью в ткани;
(2) скоростью потребления кислорода тканями.
в) Диффузия кислорода из периферических капилляров в клетки ткани. Клетки непрерывно потребляют кислород. Следовательно, PO2 в клетках периферических тканей постоянно ниже, чем PO2 в периферических капиллярах, поэтому нормальное PO2 внутри клеток может оказаться как низким (5 мм рт. ст.), так и довольно высоким (40 мм рт. ст.) и в среднем (при прямом измерении у низших животных) составляет 23 мм рт. ст. Обычно для полного обеспечения кислородом клеточных процессов вполне достаточно PO2, равного 1-3 мм рт. ст.
г) Диффузия двуокиси углерода из клеток периферических тканей в капилляры и из легочных капилляров в альвеолы. Кислород в клетках практически весь превращается в двуокись углерода, в результате в клетке повышается PCO2. Из-за повышения PCO2 двуокись углерода диффундирует из клеток в тканевые капилляры и переносится кровью в легкие. Там она диффундирует из легочных капилляров в альвеолы и выводится из легких.
Таким образом, в каждой точке транспортного конвейера направление диффузии двуокиси углерода прямо противоположно направлению диффузии кислорода. Но в процессе диффузии двуокиси углерода и кислорода имеется одно большое различие: скорость диффузии двуокиси углерода примерно в 20 раз выше, чем скорость диффузии кислорода, поэтому градиенты давления, необходимые для диффузии двуокиси углерода, в каждой точке намного меньше таковых для диффузии кислорода. Парциальное давление CO2 в разных точках имеет приблизительно следующие значения.
1. PCO2 внутри клетки равно 46 мм рт. ст., PCO2 в интерстициальной ткани — 45 мм рт. ст. Таким образом, градиент составляет только 1 мм рт. ст. (для облегчения понимания просим вас изучить рисунок ниже)
![]()
Связывание двуокиси углерода кровью в тканевых капиллярах (PCO2 в клетках ткани — 46 мм рт. ст., в интерстициальной жид кости — 45 мм рт ст.)
2. PCO2 в артериальной крови, поступающей в ткани, равно 40 мм рт. ст., PCO2 в венозной крови, покидающей ткани, — 45 мм рт. ст. Таким образом, PCO2 крови в тканевых капиллярах практически уравновешивается с PCO2 в интерстициальной ткани (45 мм рт. ст.).
3. PCO2 крови, поступающей в артериальный конец легочных капилляров, равно 45 мм рт. ст.;
PCO2 в альвеолярном воздухе — 40 мм рт. ст. Таким образом, для обеспечения всей необходимой диффузии двуокиси углерода из легочных капилляров в альвеолы достаточно градиента парциального давления двуокиси углерода, равного 5 мм рт. ст. Далее (для облегчения понимания просим вас изучить рисунок ниже) PCO2 в крови легочных капилляров падает и становится практически равным PCO2 в альвеолярном воздухе (40 мм рт. ст.) прежде, чем эта кровь пройдет через первую треть капилляра. Такую же картину мы наблюдали ранее при рассмотрении процесса диффузии кислорода, но только в противоположном направлении.
![]() Диффузия двуокиси углерода из капиллярной крови в альвеолу
Диффузия двуокиси углерода из капиллярной крови в альвеолу ![]() Влияние кровотока и скорости метаболизма на PCO2 в периферической ткани
Влияние кровотока и скорости метаболизма на PCO2 в периферической ткани
д) Влияние интенсивности метаболизма ткани и тканевого кровотока на PCO2 интерстициальной ткани. Капиллярный кровоток в ткани и метаболизм ткани оказывают на PCO2 действие, противоположное их влиянию на PO2 в ткани. На рисунке выше показано, что:
1. Снижение кровотока от нормального уровня до 1/4 нормы приводит к повышению PCO2 в периферической ткани от нормы 45 мм рт. ст. до 60 мм рт. ст., а повышение кровотока в 6 раз относительно нормы (точка В) снижает PCO2 в интерстициальной ткани от 45 мм рт. ст. до 41 мм рт. ст., что почти равно PCO2 во входящей в тканевые капилляры артериальной крови (40 мм рт. ст.).
2. Повышение интенсивности метаболизма ткани в 10 раз намного повышает PCO2 в интерстициальной жидкости при всех уровнях кровотока, а снижение метаболизма до 1/4 от нормального уровня вызывает падение PCO2 в интерстициальной жидкости примерно до 41 мм рт. ст., что близко к уровню PCO2 в артериальной крови (40 мм рт. ст.).
Видео физиология газообмена в легких и транспорта газов кровью — профессор, д.м.н. П.Е. Умрюхин
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021