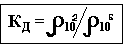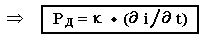Цель работы: изучить зависимость импеданса от частоты на примерах эквивалентных электрических схем и живой ткани.
Литература: Ремизов А.Н. Медицинская и биологическая физика. -М.: Высшая школа. 1999.С. 315-322.
Приборы и материалы: вольтметр, амперметр, генератор звуковой частоты, экспериментальная установка.
Метод измерения импеданса (полного электрического сопротивления) позволяет изучать структуру живого вещества, не повреждая его. Измерение электрического сопротивления используют:
Важной характеристикой состояния органов и тканей является кровоснабжение, которое также можно исследовать с помощью определения полного электрического сопротивления тканей. Ткани живых органов состоят из клеток, омываемых тканевой жидкостью. Цитоплазма клеток и тканевая жидкость представляют собой электролиты, разделенные плохо проводящей клеточной мембраной — диэлектриком. Такие системы в электрическом отношении подобны конденсаторам (С). Тканевые жидкости содержат ионы электролитов, своего рода свободные переносчики зарядов. Поэтому они обладают активным (омическим) сопротивлением (R).
При исследовании зависимости импеданса живых тканей от частоты переменного тока в широком диапазоне частот была обнаружена следующая зависимость: высокое значение (до 3000 Ом) при низких частотах с уменьшением (до 200 Ом) при высоких частотах. Подобная зависимость свойственна всем живым тканям. Зона дисперсии импеданса обычно варьирует в интервале 10² — 10 8 Гц. У многих объектов минимальное сопротивление наблюдается при частоте 10 8 Гц. Дисперсия импеданса живых тканей является результатом того, что при низких частотах, как и при постоянном токе, сопротивление связано с поляризацией, по мере увеличения частоты поляризационные явления сказываются меньше. Дисперсия импеданса свойственна только живым тканям, это подтверждает динамика кривых, показывающих зависимость импеданса (Z) растительной ткани от частоты: в норме (рис.а), после нагревания (рис.б) и при полном отмирании ткани (рис. в).
Закономерность изменения импеданса живой ткани с частотой можно объяснить следующим образом: сопротивление живых клеток является суммарным и складывается из омического сопротивления, которое не зависит от частоты тока, и емкостного сопротивления, которое значительно уменьшается по мере увеличения частоты. Это приводит к уменьшению полного сопротивления всей системы. Следовательно, можно считать, что явление дисперсии импеданса клеток и тканей есть результат уменьшения их емкостного сопротивления с увеличением частоты.
Кроме дисперсии электропроводности живой ткани, были отмечены и другие особенности:
а) сопротивление переменному току ниже, чем постоянному;
б) сопротивление не зависит от величины тока, если величина не превышает физиологическую норму;
в) сопротивление на данной частоте постоянно, если не изменяется их физиологическое состояние;
г) сопротивление изменяется при изменении физиологического состояния объекта, при отмирании ткани оно уменьшается.
Для изучения закономерностей прохождения переменного тока через биологические ткани используют эквивалентные схемы, т.е. такие комбинации соединения омического сопротивления и емкости, которые в первом приближении могут моделировать электрические свойства клеток.
Подберем эквивалентную электрическую схему живой ткани, для этого рассмотрим примеры несложных электрических схем с конденсатором (С) и резистором (R). Пусть схема состоит из последовательно соединенных омического сопротивления и конденсатора (схема 1), импеданс (Z1) такой схемы определяется выражением:
(1.1)
При малых частотах импеданс будет большим, т.к. емкостное сопротивление при этом резко увеличивается. При параллельном соединении омического сопротивления и конденсатора (схема 2), импеданс (Z2) такой схемы определяется выражением:
(1.2)
В этом случае при больших частотах импеданс системы стремится к нулю, так как при этом емкостное сопротивление становится минимальным.
Сравнивая графики, изображенные на рисунках для схем 1 и 2, с зависимостью для живой ткани, легко заметить их несходство. Схема 3 лучше других повторяет свойства живой ткани. Импеданс такой схемы определяется выражением:
(1.3)
По построенной релаксационной кривой Z(v), исходя из модели (1.3), можно определить величины сопротивлений и ёмкости эквивалентной электрической схемы живой ткани:
В данной работе определим импеданс с помощью амперметра и вольтметра. Схема установки представлена на рисунке. Импеданс эквивалентной схемы определяем по закону Ома: Z = U/ I, где Z — импеданс эквивалентной схемы (Ом), I — сила тока (А), U — напряжение (В). Изменяя частоту сигнала, подаваемого с помощью звукового генератора на эквивалентную схему, исследуем зависимость импеданса от частоты.
V. Особенности живой ткани как проводника переменного тока. Дисперсия электропроводности и ее количественная оценка.
Живая ткань как проводник переменного тока имеет следующие особенности:
Полное сопротивление живой ткани Z зависит от ее вида, физиологического состояния (кровенаполнения) и от частоты тока.
Сопротивление живой ткани переменному току меньше чем постоянному.
С увеличением частоты импеданс живой ткани не линейно уменьшается до определенного значения, а затем остается постоянным.
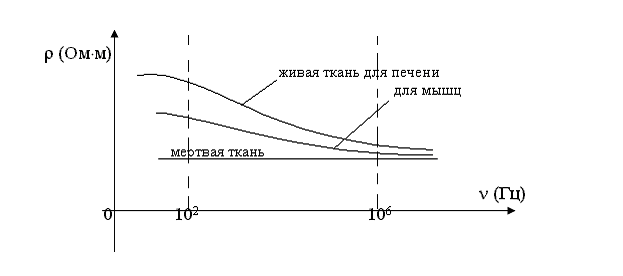
При прохождении переменного тока через живые ткани наблюдается дисперсия электропроводности – это явление зависимости удельного сопротивления живой ткани от частоты переменного тока. Это явление связывают с наличием в живых тканях поляризации, которая с увеличением меньше влияет на удельное сопротивление.
Рассмотрим дисперсионные кривые.
— частота, — удельное сопротивление живой ткани.
Интервал частот = 10 2 10 6 – один из интервалов на котором дисперсия ярко выражена. Таких интервалов существует несколько.
Дисперсия присуща только живым тканям и отсутствует у мертвых тканей.
Наиболее выражена на частотах до 1 МГц.
На практике используется для оценки физиологического состояния и жизнеспособности ткани.
Для количественной оценки дисперсии используют коэффициент дисперсии «Кд» – это безразмерная величина, равная отношению удельного сопротивления на частоте = 10 2 Гц к удельному сопротивлению при = 10 6 Гц.
Ее физиологического состояния
Эволюционной стадии развития организма (для печени млекопитающих КД = 9 – 10 относительных единиц, для печени лягушки КД = 2 – 3 относительных единицы).
При умирании ткани КД стремится к 1.
VI. Виды действия переменного тока на живые ткани. Законы раздражающего действия переменного тока и их характеристика.
Переменный ток оказывает на организм лечебное и поражающее действие. Его действие определяется следующими факторами:
Видом ткани и ее физиологическим состоянием;
Тепловое (определятся плотностью и временем действия). Лежит в основе терапии.
Специфическое физиологическое действие. Лежит в основе терапии.
Раздражающее – это действие проявляется только для возбудимых тканей (нервной, мышечной и железистой). Виды этого действия зависят от физиологического состояния клетки, от характеристик тока. Раздражающее действие лежит в основе диагностического применения и подчиняется трем законам.
Наибольшая скорость изменения тока у прямоугольных импульсов, Следовательно они обладают наибольшим РД. Такие токи можно получить размыканием (замыканием) ключа электрического тока.
Тут вы можете оставить комментарий к выбранному абзацу или сообщить об ошибке.
ЭЛЕКТРОПРОВОДНОСТЬ БИОЛОГИЧЕСКИХ СИСТЕМ
ЭЛЕКТРОПРОВОДНОСТЬ БИОЛОГИЧЕСКИХ СИСТЕМ — количественная характеристика способности живых объектов (тканей) проводить электрический ток. Электропроводность обратно пропорциональна величине электрического сопротивления системы.
Измерение электропроводимости биологических систем используют для получения информации о функциональном состоянии биол, тканей, для выявления воспалительных процессов, изменения проницаемости клеточных мембран и стенок сосудов при патологии или действии на организм физических, химических и других факторов (см. Проницаемость). Измерение электропроводимости биологических систем лежит в основе многих методов оценки кровенаполнения сосудов органов и тканей (см. Реография).
При подаче разности потенциалов (U) через объект течет электрический ток силой (I), величина которой пропорциональна электропроводности (G): I=GU. Величина электропроводности зависит от количества свободных электрических зарядов и их подвижности. Чем больше количество зарядов и их подвижность, тем больше электропроводность. В клетке основными свободными зарядами являются ионы калия и органические анноны, а снаружи клетки, в межклеточных пространствах и тканевых жидкостях — ионы натрия и хлора. В биологических жидкостях (кровь, спинномозговая жидкость, моча и т. д.) электропроводность пропорциональна содержанию в них свободных ионов. Связанные заряды (ионогенные группы белков, липидов, углеводов), перемещение которых ограничено, и крупные ионы с малой подвижностью не оказывают существенного влияния на величину электропроводимости биологических систем.
Электропроводность или сопротивление клеток, тканей, органов и целых организмов измеряют при пропускании через них постоянного или переменного синусоидальной формы тока, частота которого может изменяться от долей герца до 10 10 гц. При измерениях на переменном токе с круговой частотой ω (ω = 2πf, где f — частота в гц) общее сопротивление системы, или импеданс (см.), зависит от наличия границ раздела в системе, на которых может происходить накопление зарядов — поляризация (см.). Свойства границ раздела (в биологическом объекте это главным образом различные мембраны) могут быть описаны, если ввести понятие емкости С, сопротивление которой Хс (реактивное сопротивление в отличие от R — активного сопротивления) зависит от частоты, на которой производится измерение: Xс = 1/(ωC).
Общее сопротивление (Z) равно сумме сопротивлений реактивного Хc и активного R, если R и С соединены последовательно; при параллельном соединении R и С общее сопротивление рассчитывается по формуле:
Для измерения электрических характеристик биол. объекта применяют металлические или угольные электроды (см.), которые прикладывают к объекту с помощью жидкостного контакта — тонкого слоя хорошо проводящей жидкости, чаще всего — физиологического раствора. В ряде случаев, напр. при измерении электропроводности цитоплазматических мембран, один из электродов вводится внутрь клетки, а другой подводится к клетке снаружи (см. Микроэлектродный метод исследования). При измерении электропроводимости биологических систем на постоянном и переменном токах низкой частоты важно учитывать величину поляризации электродов, поскольку за счет электродной поляризации истинные электрические параметры биол, объекта могут значительно отличаться от измеренных. Величина поляризации электродов определяется плотностью тока, которая зависит от приложенной к системе разности потенциалов, сопротивления системы, площади измерительных электродов. Чем больше площадь электродов, тем меньше плотность тока и тем меньше искажения, вносимые в измерения электродами. Поэтому для уменьшения плотности тока используют электроды с большой эффективной поверхностью, в частности платиновые электроды, покрытые платиновой чернью (их губчатое покрытие увеличивает эффективную поверхность электродов в 100 — 1000 раз). Возможно применение и так называемых неполяризующихся электродов (например, каломельных, хлорсеребряных).
Для облегчения интерпретации получаемых результатов биологическую систему (ткани, суспензии клеток) часто представляют в виде модели — электрической схемы, состоящей из набора активных сопротивлений и емкостей, являющихся как бы эквивалентами биологических структур клеток или тканей, участвующих в проведении электрического тока.
Измерение электропроводимости биологических систем на постоянном токе из-за высокой степени поляризации мембран и электродов крайне затруднено. На низких частотах переменного тока большая часть тока протекает по межклеточным промежуткам. При увеличении частоты электрического тока реактивное сопротивление емкости падает, поляризационные явления уменьшаются. Зависимость сопротивления и емкости объекта от частоты получила название дисперсии (см.). На высоких частотах общее сопротивление системы зависит только от активных сопротивлений межклеточных пространств и цитоплазмы.
В медицине и биологии электропроводимость биологических систем чаще всего исследуют в области так называемой β-дисперсии, которая наблюдается в диапазоне частот 10 2 —10 8 гц и определяется поляризацией границ раздела и неоднородностью структуры объекта. Измерения электропроводимости биологических систем показали, что по мере повышения частоты электропроводность возрастает, достигая предельной величины. При переживании и отмирании ткани возрастает электропроводимость биологических систем на низких частотах. Это связано с тем, что при отмирании ткани растет проницаемость мембран для ионов, и они уже не являются границей, на которой может происходить поляризация. Основываясь на способности живой ткани к поляризации, Б. Н. Тарусов предложил в качестве критерия оценки жизнеспособности ткани использовать коэффициент К, численно равный отношению Rн/Rв где Rн и Rв — сопротивления ткани, измеренные соответственно на низкой и высокой частотах; при отмирании ткани он стремится к единице. Выбор частот для расчета К определяется диапазоном β-дисперсии: низкой частоте соответствует частота начала β-дисперсии, высокой — частота, при которой электропроводимость биологических систем достигает максимальной величины. Например, для мышечной ткани — это 10 3 и 10 6 гц, клеток крови и жировой ткани — 10 4 и 10 7 гц, кожи — 10 2 и 10 4 гц и т. д. На высоких частотах, когда активное и общее сопротивления не зависят от частоты, возможно исследование относительных изменений числа ионов в биол. системе, связанных с нарушением водно-солевого обмена.
Сопоставляя Данные, полученные при измерении на низких и высоких частотах, можно вычислить объем и ионную проводимость межклеточных пространств и цитоплазмы клеток, проницаемость мембран для ионов, емкостные характеристики мембраны. Если измерения проводятся в системе, где межклеточные пространства занимают достаточно большой объем (более 20—30%), например при измерениях электропроводности крови, то для вычисления параметров дисперсной фазы (эритроцитов) используют специальные формулы. Частоты, на которых наблюдается дисперсия, зависят от величины клеток и объема межклеточных пространств. Так, дисперсия электропроводимости биологических систем для клеток крови начинается на частотах порядка нескольких десятков килогерц, для мышечной ткани — несколько килогерц, жировой — сотен килогерц. При исследовании электрических характеристик плазматических мембран клеток дисперсия обнаруживается на частотах порядка нескольких десятков герц. Электрические характеристики тканей и органов на низких частотах зависят от неоднородности расположения клеток и межклеточных пространств и соотношения их объемов. Этот факт используется в реографии и реоэнцефалографии (см.) при исследовании изменений кровенаполнения ткани и эластических свойств стенок сосудов. Измерение электропроводимости биологических систем на низких частотах позволяет оценить изменения объема межклеточных пространств, в частности при развитии воспаления (см.). Так, на первых стадиях воспалительного процесса структура клеток изменяется незначительно, и импеданс клеток сохраняет свою величину. По мере набухания клеток и уменьшения объема межклеточных пространств происходит увеличение общего сопротивления системы. На более поздних стадиях развития воспаления импеданс системы уменьшается за счет возрастания проницаемости мембран для различных ионов.
Таким образом, измерение электропроводимости биологических систем или импеданса, особенно в широком диапазоне частот, может быть использовано при исследовании проницаемости клеточных мембран и других границ раздела в клетках, тканях, органах, а стандартизация (измерение удельных величин) дает возможность сравнивать данные, полученные разными исследователями. Возбуждение, изменение интенсивности метаболизма и других функций клеток приводят к изменению электропроводимости биологических систем. Методы измерения электропроводимости биологических систем используют для исследования влияния на биологические объекты различных факторов: работы (увеличение интенсивности метаболизма приводит к увеличению проницаемости мембран); психогенных (изменяется проницаемость кожи за счет работы потовых желез); физических (радиация, ультрафиолетовое излучение, температура и др.) и химических (кислоты, щелочи, спирты и др.), обычно сопровождаемых ростом проницаемости. Изменение проницаемости мембран часто зависит от дозы или концентрации действующего вещества. Так, соли меди в малых концентрациях уменьшают проницаемость мембран мышечных клеток кожи лягушки, а в концентрациях более 10 -3 М — увеличивают. Исследование электрических свойств возбудимых тканей способствовало изучению механизма проведения возбуждения по нерву п мышце. На основании измерений активного сопротивления, емкости и их дисперсии была вычислена статическая емкость клеточной мембраны (около 1 мкф/см 2 ) и впервые определена толщина ее липидного бислоя. Было найдено, что удельное сопротивление аксоплазмы и миоплазмы всего в 2—3 раза выше сопротивления внеклеточной жидкости, тогда как сопротивление мембраны выше в десятки тысяч раз. Эти данные послужили основанием для возникновения представления о «кабельной» структуре волокна. Установлены временные соотношения между изменением проницаемости мембраны для ионов и развитием потенциала действия — «импедансный спайк» (см. Биоэлектрические потенциалы, Нервный импульс). Исследование электропроводимости биологических систем может быть использовано для оценки состояния тканей при их консервации, а также эффективности действия биологически активных веществ на модельные системы. В ряде случаев проницаемость биол. мембран для ионов сопряжена с их проницаемостью для незаряженных частиц— сахаров, аминокислот и других соединений. Поэтому измерение электропроводимости биологических систем может оказаться полезным при изучении проницаемости мембран и для неэлектролитов. Исследование электпроводимости биологических систем может найти применение и в биотехнологии для оценки оптимальности среды и условий культивирования клеток.
Библиогр.: Андреев В. С. Кондуктометрические методы и приборы в биологии и медицине, М., 1973; Биофизика, под ред. Б. Н. Тарусова и О. Р. Колье, с. 186, М., 1968; Гречин В. Б. и Боровикова В. Н. Медленные неэлектрические процессы в оценке функционального состояния мозга человека, с. 22, Л, 1982; Гуревич М. И. и др. Импедансная реоплетизмография, Киев, 1982; Егоров Ю. В. и Кузнецова Г. Д. Мозг как объемный проводник, М., 1976; Слынько П. П. Основы низкочастотной кондуктометрии в биологии, М., 1972; Хассет Дж. Введение в психофизиологию, пер. с англ., с. 53, М., 1981; Электроника и кибернетика в биологии и медицине, пер. с англ., под ред. П. К. Анохина, с. 71, М., 1963; Schwan Н. P. Electrical properties of tissue and cell suspensions, Advanc, biol. med. Phys., v. 5, p. 147, 1957.