
Гексозомонофосфатный путь превращения глюкозы в тканях
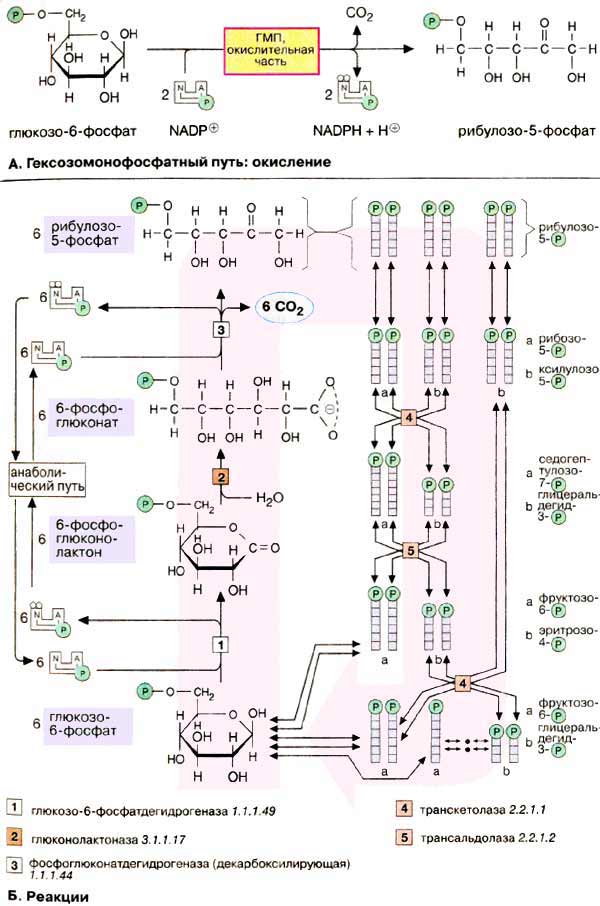
Гексозомонофосфатный путь [ГМП (HMW), часто называемый также пентозофосфатным путем ] является окислительным обменом веществ в цитоплазме, в котором, как и в гликолизе, исходным субстратом служит глюкозо-6-фосфат . ГМП поставляет два важных исходных соединения для анаболических процессов: НАДФН + Н + (NADPH + Н + ), необходимый для биосинтеза жирных кислот и изопреноидов (см. с. 170), и рибозо-5-фосфат , предшественник в биосинтезе нуклеотидов (см. с. 190).
А. Гексозомонофосфатный путь: окисление
В процессе окисления глюкозо-6-фосфат превращается в рибулозо-5-фосфат. При этом образуются 1 молекула CO 2 и 2 НАДФН + Н + . Значительно более сложная часть пути — восстановительная ( Б ) — в зависимости от обмена веществ либо превращает часть образованного пентозофосфата снова в гексозофосфат, либо включает его в гликолиз для деградации. В большинстве клеток за счет ГМП разрушается не более 10% глюкозо-6-фосфата.
Окислительная часть ГМП начинается с окисления глюкозо-6-фосфата глюкозо-6-фосфатдегидрогеназой [ 1 ]. При этом образуется НАДФН + Н + и 6-фосфоглюколактон — внутримолекулярный сложный эфир (лактон) 6-фосфоглюконата. Специфическая гидролаза (фермент [ 2 ]) расщепляет сложноэфирную связь и оставляет свободной карбоксильную группу 6-фосфоглюконата . Последний фермент окислительной части, фосфоглюконатдегидрогеназа ( 3 ), отщепляет карбоксильную группу 6-фосфоглюконата в виде CO 2 с одновременным окислением гидроксильной группы при С-3 до кетогруппы. Наряду со второй молекулой НАДФН + Н + при этом образуется кетопентоза, рибулозо-5-фосфат , которая под действием изомеразы превращается в рибозо-5-фосфат, исходное соединение для нуклеотидного синтеза (на схеме сверху).
Восстановительная часть ГМП показана здесь только схематически. Полная схема реакции представлена на с. 396.
Функция восстановительной ветви состоит в том, чтобы производство НАДФН + Н + и пентозофосфатов соответствовало метаболическим потребностям клеток. Обычно потребность в НАДФН + Н + намного выше, чем в пентозофосфатах. В этих условиях 6 молекул рибулозо-5-фосфата под действием трансальдолаз и транскетолаз образуют 5 молекул фруктозо-6-фосфата, которые изомеризуются в 5 молекул глюкозо-6-фосфата. Глюкозо-6-фосфат вновь участвует в окислительной части ГМП в процессе получения НАДФН + Н + . Неоднократное повторение этих реакций позволяет окислить глюкозо-6-фосфат до 6 молекул CO 2 . При этом образуется 12 молекул НАДФН + Н + , а пентозофосфат не образуется.
При взаимном превращении фосфатов сахаров в восстановительной части ГМП особенно важны два фермента. Трансальдолаза [ 5 ] переносит С 3 -звенья от седогептулозо-7-фосфата, кетосахара с 7 атомами углерода, на альдегидную группу глицеральдегид-3-фосфата. Аналогичным образом транскетолаза [ 4 ] катализирует перенос С 2 -фрагмента с одного фосфата сахара на другой.
Реакции восстановительной части ГМП обратимы, т.е. гексозофосфаты могут непосредственно превращаться в пентозофосфаты. Это превращение может происходить при высокой потребности клетки в пентозофосфатах, например на стадии репликации ДНК и S-фазе клеточного цикла (см. с. 380).
Читайте также: Итальянский джерси ткань что это такое
Если наряду с НАДФН + Н + клетке требуется энергия в форме АТФ, продукты восстановительной части ГМП (фруктозо-6-фосфат и глицеральдегид-3-фосфат) включаются в гликолиз и далее в цитратный цикл и дыхательную цепь с образованием CO 2 и воды. На этом пути из 6 молей глюкозо-6-фосфата образуется 12 молей НАДФН + Н + и примерно 150 молей АТФ.
ГЕКСОЗОМОНОФОСФАТНЫЙ ШУНТ
ГЕКСОЗОМОНОФОСФАТНЫЙ ШУНТ – ферментативный процесс прямого аэробного окисления фосфорилированной глюкозы до CO2 и H2O, протекающий в цитоплазме живых клеток и сопровождающийся накоплением важного кофермента – восстановленного никотинамидадениндинуклеотидфосфата (НАДФ-Н)- и образованием производных пентоз, откуда он и получил другие названия – пентозофосфатный или пентозный путь или цикл. Он известен еще и как фосфоглюконатный цикл. Свое название этот путь получил потому, что при его реализации глюкозо-6-фосфат (Г6Ф) выключается из гликолиза. Этот шунт был исследован в 1950-е Ф.Дикенсом, Ф.Липманом, Э.Рэкером и Б.Хорекером после открытия (О.Варбург, 1931) глюкозо-6-фосфатдегидрогеназы (КФ1.1.1.49). Гексозомонофосфатный шунт состоит из окислительного декарбоксилирования (3 первых реакции) Г6Ф (от гексозы отщепляется первый атом углерода) и неокислительных превращений пентозофосфатов (5 последующих реакций) с образованием исходного Г6Ф:
6 Г6Ф + 12НАДФ + = 6CO2 + 12НАДФ-Н + 12H + + 5 Г6Ф + H3PO4
Полная последовательность его такова:
1) НАДФ + + Г6Ф = 6-фосфоглюконолактон + НАДФ-Н + Н +
2) 6-фосфоглюконолактон + Н2О = 6-фосфоглюконат
3) НАДФ + + 6-фосфоглюконат = рибулозо-5-фосфат + СО2 + НАДФ-Н + Н +
4) рибулозо-5-фосфат = рибозо-5-фосфат
5) рибулозо-5-фосфат = ксилозо-5-фосфат
6) рибозо-5-фосфат + ксилозо-5-фосфат = глицеральдегид-3-фосфат + седогептулозо-7-фосфат
7) седогептулозо-7-фосфат + глицеральдегид-3-фосфат = фруктозо-6-фосфат + эритрозо-4-фосфат
8) ксилозо-5-фосфат + эритрозо-4-фосфат = фруктозо-6-фосфат + глицеральдегид-3-фосфат
Многие промежуточные соединения гексозомонофосфатного шунта могут участвовать в других процессах в организме – гликолизе, фотосинтезе и др. Ферменты гексозомонофосфатного шунта найдены в тканях животных, растений и в микроорганизмах. Его главные метаболические функции заключаются в том, чтобы:
1) служить источником НАДФ-Н для анаболических (биосинтетических) путей и 2) обеспечить промежуточные продукты для других биосинтезов, особенно рибозо-5-фосфата для синтеза нуклеиновых кислот и эритрозо-4-фосфата для биосинтеза фенилаланина, тирозина и триптофана. Важная особенность гексозомонофосфатного шунта – его гибкость: в разных ситуациях он может функционировать частично, и суммарный результат будет различным. Доля шунта в количественном превращении глюкозы обычно невелика, варьируется у разных организмов и зависит от типа ткани и её функционального состояния. У млекопитающих активность гексозомонофосфатного шунта высока в печени, надпочечниках, в эмбриональной ткани и в молочной железе в период лактации. Особенно высока его активность в жировой ткани и эритроцитах (ок. 50%). Главным регуляторным звеном шунта является первый фермент глюкозо-6-фосфатдегидрогеназа. Центральные стадии шунта – перестройка углеродного скелета сахаров – катализируются трансальдолазой (КФ 2.2.1.2) и транскетолазой (КФ 2.2.1.1). Для активности последней необходимы ионы Мg 2+ и тиаминпирофосфат – метаболически активная форма витамина В1.
Читайте также: Производство ткани 6 букв ответ
Врожденная недостаточность некоторых ферментов гексозомонофосфатного шунта в организме человека приводит к гемолитическим анемиям, синдрому Вернике – Корсакова и т.п. Скорость его определяется, в первую очередь, концентрацией НАДФ-Н. Он регулируется инсулином и другими гормонами, влияющими на углеводный обмен, а также глутатионом.
Hollmann S. Non-glycolytic pathways of metabolism of glucose. N. Y., 1964
Страйер Л. Биохимия, пер. с англ., М., Мир, 1985
Бохински Р. Современные воззрения в биохимии, пер. с англ., М, Мир, 1987
Гексозомонофосфатный путь
ГЕКСОЗОМОНОФОСФАТНЫЙ ПУТЬ ПРЕВРАЩЕНИЯ ГЛЮКОЗЫ
Окисление глюкозы по этому пути протекает в цитоплазме клеток и представлено двумя последовательными ветвями: окислительной и неокислительной. Особенно активно этот путь протекает в тех органах и тканях, в которых активно синтезируются липиды (печень, почки, жировая и эмбриональная ткань, молочные железы).
Биологическая роль этого пути окисления глюкозы связывается прежде всего с производством двух веществ:
1.НАДФ*Н2, который в отличие от НАДН2 , не окисляется в дыхательной цепи митохондрий, а используется в клетках в реакциях синтеза и восстановления и гидроксилирования веществ.
2.РИБОЗО-5-ФОСФАТ и его производные, которые используются в клетке для синтеза важнейших биологических молекул: нуклеиновых кислот (ДНК, РНК), нуклеозидтрифосфатов(НТФ) коферментов (, НАД, ФАД, Н5КОА).
Биологическая роль:1 .АНАБОЛИЧЕСКАЯ.
2.ЭНЕРГЕТИЧЕСКАЯ. При низком энергетическом статусе клетки излишки пентоз путём обратных реакций неокислительного пути превращаются в глицеральдегид-3-фосфат и фруктозо-6-фосфат которая затем включаются в анаэробный гликолиз, поддерживая биоэнергетику клеток в кризисных ситуациях.
Гексозомонофосфатный путь катаболизма глюкозы ещё обозначают как пентозный путь.
Окислительная стадия гексозомонофосфатного пути катаболизма глюкозы отличается от классического — гексозодифосфатного пути с этапа превращения глюкозо-6-фосфата:
рис. Окислительная стадия гексозомонофосфатного пути катаболизма глюкозы
Неокислительная стадия гексозомонофосфатного пути катаболизма глюкозы представлена двумя ТРАНСКЕТОЛАЗНЫМИ реакциями и одной ТРАНСАЛЬДОЛАЗНОЙ.
В результате этих реакций образуются субстраты для ГЛИКОЛИЗА, а также вещества характерные для ПЕНТОЗНОГО пути.
1. ТРАНСКЕТОЛАЗНЫЕ реакции:
2. ТРАНСАЛЬДОЛАЗНАЯ реакция:
А) СЕДОГЕПТУЛОЗО-7-Ф + ГЛИЦЕРАЛЬД-3-Ф = ФРУКТОЗО-6-Ф + ЭРИТРОЗО-4-Ф.
Баланс окислительной и неокислительной стадий гексозомонофосфатного пути превращения глюкозы можно записать в виде суммарного уравнения реакции.
3.4.3. Гексозомонофосфатный путь
ГЕКСОЗОМОНОФОСФАТНЫЙ ПУТЬ ПРЕВРАЩЕНИЯ ГЛЮКОЗЫ
Окисление глюкозы по этому пути протекает в цитоплазме клеток и представлено двумя последовательными ветвями: окислительной и неокислительной. Особенно активно этот путь протекает в тех органах и тканях, в которых активно синтезируются липиды (печень, почки, жировая и эмбриональная ткань, молочные железы).
Читайте также: Трусливая ткань 4 буквы
Биологическая роль этого пути окисления глюкозы связывается прежде всего с производством двух веществ:
1.НАДФ*Н2, который в отличие от НАДН2 , не окисляется в дыхательной цепи митохондрий, а используется в клетках в реакциях синтеза и восстановления и гидроксилирования веществ.
2.РИБОЗО-5-ФОСФАТ и его производные, которые используются в клетке для синтеза важнейших биологических молекул: нуклеиновых кислот (ДНК, РНК), нуклеозидтрифосфатов(НТФ) коферментов (, НАД, ФАД, Н5КОА).
Биологическая роль:1 .АНАБОЛИЧЕСКАЯ.
2.ЭНЕРГЕТИЧЕСКАЯ. При низком энергетическом статусе клетки излишки пентоз путём обратных реакций неокислительного пути превращаются в глицеральдегид-3-фосфат и фруктозо-6-фосфат которая затем включаются в анаэробный гликолиз, поддерживая биоэнергетику клеток в кризисных ситуациях.
Гексозомонофосфатный путь катаболизма глюкозы ещё обозначают как пентозный путь.
Окислительная стадия гексозомонофосфатного пути катаболизма глюкозы отличается от классического — гексозодифосфатного пути с этапа превращения глюкозо-6-фосфата:


рис. Окислительная стадия гексозомонофосфатного пути катаболизма глюкозы
Неокислительная стадия гексозомонофосфатного пути катаболизма глюкозы представлена двумя ТРАНСКЕТОЛАЗНЫМИ реакциями и одной ТРАНСАЛЬДОЛАЗНОЙ.
В результате этих реакций образуются субстраты для ГЛИКОЛИЗА, а также вещества характерные для ПЕНТОЗНОГО пути.
1. ТРАНСКЕТОЛАЗНЫЕ реакции:


2. ТРАНСАЛЬДОЛАЗНАЯ реакция:
А) СЕДОГЕПТУЛОЗО-7-Ф + ГЛИЦЕРАЛЬД-3-Ф = ФРУКТОЗО-6-Ф + ЭРИТРОЗО-4-Ф.
Баланс окислительной и неокислительной стадий гексозомонофосфатного пути превращения глюкозы можно записать в виде суммарного уравнения реакции.



3.4.4. Глюконеогенез
Основными источниками глюкозы для организма человека являются:
ГЛЮКОНЕОГЕНЕЗ — это биосинтез глюкозы из неуглеводных предшественников, главными из которых являются ПИРУВАТ, ЛАКТАТ, ГЛИЦЕРИН, МЕТАБОЛИТЫ ЦТК КРЕБСА, АМИНОКИСЛОТЫ.
ГЛЮКОНЕОГЕНЕЗ возможен не во всех тканях. Главным местом синтеза глюкозы является печень, в меньшей степени процесс идёт в почках и слизистой кишечника. Биологическая роль глюконеогенеза заключается не только в синтезе глюкозы, но и в возвращении лактата, образованного в реакциях анаэробного ГЛИКОЛИЗА, в клеточный фонд углеводов. За счет этого процесса поддерживается уровень глюкозы в тканях в кризисных ситуациях (при углеводном голодании, сахарном диабете, тканевой гипоксии).
Большинство реакций ГЛЮКОНЕОГЕНЕЗА представляют собой обратные реакции ГЛИКОЛИЗА, за исключением трёх термодинамически необратимых: ПИРУВАТКИНАЗНОЙ, ФОСФОФРУКТОКИНАЗНОЙ, ГЕКСОКИНАЗНОЙ. Эти реакции при ГЛЮКОНЕОГЕНЕЗЕ имеют обходные пути и связаны с образованием 2-фосфоенолпирувата, фруктозо-6-фосфата и глюкозы.


Образовавшаяся в реакциях глюконеогенеза, глюкоза может вновь участвовать в клеточном метаболизме как пластический, энергетический материал, откладываться про запас в виде гликогена.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом