
Общая гистология — введение, понятие ткани, классификация.
В результате эволюционного развития у высших многоклеточных организмов возникли ткани.
Ткани — это исторически (филогенетически) сложившиеся системы клеток и неклеточных структур, обладающих общностью строения, в ряде случаев — общностью происхождения, и специализированные на выполнении определенных функций.
В любой системе все ее элементы упорядочены в пространстве и функционируют согласованно друг с другом; система в целом обладает при этом свойствами, не присущими ни одному из ее элементов, взятому в отдельности. Соответственно и в каждой ткани ее строение и функции несводимы к простой сумме свойств отдельных входящих в нее клеток.
Ведущими элементами тканевой системы являются клетки. Кроме клеток, различают клеточные производные и межклеточное вещество.
К производным клеток относят симпласты (например, мышечные волокна, наружная часть трофобласта), синцитий (развивающиеся мужские половые клетки, пульпа эмалевого органа), а также постклеточные структуры (эритроциты, тромбоциты, роговые чешуйки эпидермиса и т. д.).
Межклеточное вещество подразделяют на основное вещество и на волокна. Оно может быть представлено золем, гелем или быть минерализованным.
Среди волокон различают обычно три вида: коллагеновые, ретикулярные, эластические.
Гистогенез
Ткани развиваются путем гистогенеза.
Гистогенез — единый комплекс координированных во времени и пространстве процессов пролиферации, дифференцировки, детерминации, интеграции и функциональной адаптации клеточных систем.
Развитие организма начинается с одноклеточной стадии — зиготы. В ходе дробления возникают бластомеры, но совокупность бластомеров – это еще не ткань. Бластомеры на начальных этапах дробления еще не детерминированы (они тотипотентны). Если отделить их один от другого, — каждый может дать начало полноценному самостоятельному организму – механизм возникновения монозиготных близнецов. Постепенно на следующих стадиях происходит ограничение потенций. В основе его лежат процессы, связанные с блокированием отдельных компонентов генома клеток и детерминацией.
Детерминация – это процесс определения дальнейшего пути развития материала эмбриональных зачатков с образование специфических тканей (на основе блокирования отдельных генов).
Понятие «коммитирование» тесно связано с клеточным делением (т.н. коммитирующий митоз).
Коммитирование – это ограничение возможных путей развития вследствие детерминации. Коммитирование совершается ступенчато. Сначала соответствующие преобразования генома касаются крупных его участков. Затем все более детализируются, поэтому вначале детерминируются наиболее общие свойства клеток, а затем и более частные.
Как известно, на этапе гаструляции возникают эмбриональные зачатки. Клетки, которые входят в их состав, еще не окончательно детерминированы, так что из одного зачатка возникают клеточные совокупности, обладающие разными свойствами. Следовательно, один эмбриональный зачаток может служить источником развития нескольких тканей.
ГИСТОГЕНЕЗ
ГИСТОГЕНЕЗ (histogenesis; греч, histos столб, ткань + genesis происхождение) — развитие тканей животных и человека в онтогенезе.
Изучение Г. привело к обоснованию клеточной теории (Т. Шванн, 1839; Р. Peмак, 1852) и представлению о взаимозависимости эмбрионального Г. и филогистогенеза (А. И. Бабухин, 1869; И. И. Мечников, 1886). Изучение Г. разных тканей и его закономерностей — одна из важнейших задач гистологии (см.).
Эволюционное направление в изучении Г. получило глубокое освещение в трудах А. А. Заварзина (1945, 1947), Н. Г. Хлопина (1946), 3. С. Кацнельсона (1930—1970), С. И. Щелкунова (1958, 1971) и продолжает развиваться.
Источником развития тканей в организме зародыша являются мало-дифференцированные клетки зачатков. Эти клетки отличаются друг от друга лишь неспецифическими признаками (величина, форма и взаиморасположение клеток, их ядер и др.), а также самыми общими функциональными особенностями (особенности и степень интенсивности обмена веществ, темпы митотического размножения и т. д.). Их цитоплазма всегда базофильна (богата рибонуклеиновой к-той), ядерно-цитоплазматическое отношение высокое (ядра сравнительно больших размеров, цитоплазмы мало). Г. включает: увеличение числа клеток путем их митотического размножения, клеточный рост, перемещения отдельных клеток и клеточных комплексов, детерминацию клеток (закрепление путей их дальнейшего развития), дифференциацию клеток и их неклеточных производных, межклеточные взаимодействия, приводящие к интеграции (объединению в единое целое — зачаток, тканевую систему, орган, организм), отмирание клеток.
Ведущими компонентами Г., определяющими качественную сторону развития, являются дифференциация и интеграция. Каждая ткань у высших животных всегда развивается из одного определенного зачатка, а каждый зачаток дает начало лишь строго ограниченному кругу тканевых производных. С наступлением периода специфической, или тканевой, дифференциации клетки приобретают характерные для той или иной ткани структуры — специальные органеллы (миофибриллы, реснички и т. п.), специфические включения (напр., гранулы зимогена в железистых клетках, пигмент в хроматофорах и т. п.), иногда образуют неклеточные производные — межклеточные вещества в соединительных, хрящевых и костных тканях, симпласты при образовании скелетно-мышечных волокон и т. п. Во многих тканях при этом происходит пространственное разграничение камбиальных (малодифференцированных, резервных) и высокодифференцированных активно функционирующих клеток. В последних, как правило, базофилия цитоплазмы снижается, может сменяться оксифилией, ядерно-цитоплазматическое отношение чаще всего резко уменьшается в силу преимущественного роста цитоплазмы. У нервных клеток наряду с образованием длинных, проводящих возбуждение отростков подавляется способность к митозу в симпластических структурах (напр., в волокнах скелетной мускулатуры, плазмодиотрофобласте), а их ядра могут делиться амитотическим путем. Некоторые направления дифференцировки связаны с регрессивными изменениями клеток. Напр., эритроциты млекопитающих и человека теряют ядро и все органеллы, становясь «контейнерами» гемоглобина; клетки поверхностных слоев эпидермиса омертвевают, превращаясь в роговые чешуйки, и т. п. Увеличение многообразия тканей и специализированных клеточных форм в ходе онтогенеза обеспечивается дивергенцией, т. е. все большим расхождением их морфол., функциональных и биохим, свойств и признаков.
Читайте также: Пальто прямое расход ткани
Г. протекает асинхронно. Ткани, функционирующие в постнатальном периоде развития и у взрослого организма (так наз. дефинитивные), дифференцируются у зародыша сравнительно поздно, тогда как ткани, обеспечивающие жизнь самого зародыша и существующие только в течение зародышевого развития, а затем погибающие (так наз. провизорные), дифференцируются значительно раньше. При этом их развитие протекает сокращенно и ускоренно по сравнению с дефинитивными тканями (С. И. Щелкунов, 1958). В частности, эпителий желточного мешка дифференцируется значительно раньше и быстрее, чем кишечный эпителий. Наиболее раннюю и ускоренную дифференцировку претерпевает у млекопитающих и человека трофобласт (см.), обеспечивающий питание зародыша с первых же дней прикрепления яйца к слизистой оболочке матки.
Асинхронность дифференцировки присуща и дефинитивным тканям. Так, кожный эпителий дифференцируется намного раньше кишечного, начинающего функционировать позднее; нейроны вегетативной нервной системы — позже нейронов соматической. Структуры, совместно обеспечивающие выполнение одной и той же сложной функции к определенному моменту развития, напр, к моменту рождения, дифференцируются одновременно (принцип системогенеза — П. К. Анохин). В одной и той же ткани активно функционирующие элементы дифференцируются раньше, камбиальные же клетки вступают в завершающий этап дифференцировки позднее, притом не все сразу, а по мере «снашивания» и гибели высокоспециализированных, ранее дифференцировавшихся клеток. Т. о., камбий расходуется и самовоспроизводится на протяжении всей жизни организма до глубокой старости, соответственно непрерывно происходят и процессы Г., именуемые в этом случае физиол, регенерацией тканей. При травмах как размножение клеток, так и тканевая дифференцировка резко интенсифицируются, что способствует заживлению дефектов (репаративная регенерация). Перечисленные процессы объединяются под названием постнатального Г. Возрастные изменения организма приводят в старости к значительному ослаблению гистогенетических процессов.
Стойкая специфичность свойств тканей у высокоорганизованных животных и человека, в частности неспособность тканей, развивающихся из разных зачатков, превращаться друг в друга, называется гистологической тканевой детерминацией (см.). В свете данных молекулярной биологии она связана с репрессией или дерепрессией генов, ответственных за синтез тканеспецифических белков (см. Оперон).
Причинные факторы Г. могут быть разделены на следующие группы: внутренние, или наследственные (контролируемая цитоплазмой деятельность ядра по синтезу тканеспецифических форм РНК); взаимодействие между клетками или их комплексами (эмбриональными зачатками, тканями); влияние окружающей среды (механические, физ., хим. и другие воздействия). Все эти факторы сложно взаимодействуют, причем внешние факторы опосредуются внутренними (тканевой детерминацией). Нарушение нормального их соотношения ведет к разнообразным отклонениям Г. (дизэмбриогистогенезу), вплоть до полного его извращения при опухолевом росте (см. Опухоли).
Процессы детерминации и дифференцировки тканей в онтогенезе обусловлены ходом эволюционных изменений тканевых структур в филогенезе животного мира. Поэтому в ходе эмбрионального, а также репаративной Г. находят отражение этапы филогистогенеза, пройденного каждой данной тканью, нередки и повторения признаков тканевых и клеточных структур предков (гистологические рекапитуляции — А. H. Северцов, 1935). Так, не только эмбриональный Г. костей при их развитии на месте хрящевых закладок, но и процесс сращивания костей после переломов идет в той же последовательности, что и в филогенезе позвоночных (хрящевая закладка — ее замещение грубоволокнистой костной тканью — рассасывание последней и замещение пластинчатой костной тканью). Изучение Г. способствует более глубокому пониманию многих патол, процессов, происходящих в человеческом организме.
Читайте также: Раппорт переплетения ткани это
Г. отдельных тканей — см. соответствующие статьи по названиям тканей (напр., Жировая ткань, Мышечная ткань и др.).
Библиография: Гистогенез и реактивность тканей, Труды Ленингр, сан.-гиг. мед. ин-та, т. 42, 1958; Кнорре А. Г. Эмбриональный гистогенез (Морфологические очерки), Л., 1971, библиогр.; Хлопин Н. Г. Общебиологические и экспериментальные основы гистологии, Л., 1946, библиогр.; X р у щ о в Н. Г. Гистогенез соединительной ткани, М., 1976, библиогр.; Щелкунов С. И. Цитологический и гистологический анализ развития нормальных и малигнизированных структур, Л., 1971, библиогр.
Гистогенез всех видов тканей
В развитии низших и высших позвоночных отчетливо прослеживается единая общебиологическая закономерность, выражающаяся в появлении зародышевых листков и обособлении основных зачатков органов и тканей. Процесс образования тканей из материала эмбриональных зачатков составляет суть учения о гистогенезах.
Эмбриональный гистогенез, по определению А.А. Клишова (1984), — это комплекс координированных во времени и пространстве процессов пролиферации, клеточного роста, миграции, межклеточных взаимодействий, дифференциации, детерминации, программированной гибели клеток и некоторых других. Все названные процессы в той или иной мере протекают в зародыше, начиная с самых ранних стадий его развития.
Пролиферация. Основной способ деления тканевых клеток — это митоз. По мере увеличения числа клеток возникают клеточные группы, или популяции, объединенные общностью локализации в составе зародышевых листков (эмбриональных зачатков) и обладающие сходными гистогенетическими потенциями. Клеточный цикл регулируется многочисленными вне- и внутриклеточными механизмами. К внеклеточным относятся влияния на клетку цитокинов, факторов роста, гормональных и нейрогенных стимулов. Роль внутриклеточных регуляторов играют специфические белки цитоплазмы. В течение каждого клеточного цикла существуют несколько критических точек, соответствующих переходу клетки из одного периода цикла в другой. При нарушении внутренней системы контроля клетка под влиянием собственных факторов регуляции элиминируется апоптозом, либо на некоторое время задерживается в одном из периодов цикла.
Метод радиографического анализа клеточных циклов в различных тканях выявил особенности соотношения клеточной репродукции и дифференцировки. Например, если в тканях (кроветворные ткани, эпидермис) имеется постоянный фонд пролиферирующих клеток, за счет которых обеспечивается непрерывное возникновение новых клеток взамен погибающих, то эти ткани относятся к обновляющимся. Другие ткани, например, некоторые соединительные, характеризуются тем, что в них увеличение количества клеток происходит параллельно с их дифференцировкой, клетки в этих тканях характеризуются низкой митотической активностью. Это растущие ткани. Наконец, нервная ткань характеризуется тем, что все основные процессы репродукции заканчиваются в период эмбрионального гистогенеза (когда формируется основной запас стволовых клеток, достаточный для последующего развития ткани). Поэтому она отнесена к стабильным (стационарным) тканям. Продолжительность жизни клеток в обновляющихся, растущих и стабильных тканях разная.
Наряду с обновлением клеточной популяции, в самих клетках постоянно наблюдается обновление внутриклеточных структур (внутриклеточная физиологическая регенерация).
Клеточный рост, миграция и межклеточные взаимодействия. Рост клеток проявляется в изменении их размеров и формы. При усилении функциональной активности и внутриклеточных биосинтезов наблюдается увеличение объема клетки. Если объем клетки превышает некую норму, то говорят о ее гипертрофии, и наоборот, при снижении функциональной активности происходит уменьшение объема клетки, а при переходе некоторых нормативных параметров возникает атрофия клетки. Рост клетки не беспределен и определяется оптимальным ядерно-цитоплазменным отношением.
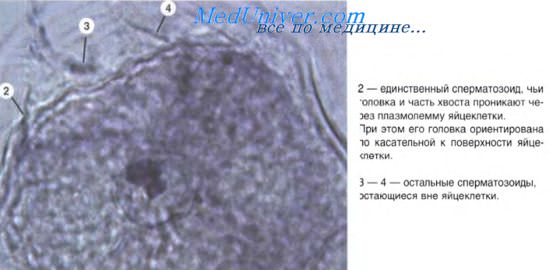
Важное значение для гистогенеза имеют процессы перемещения клеток. Миграция клеток наиболее характерна для периода гаструляции. Однако и в период гисто- и органогенеза происходят перемещения клеточных масс (например, смещения миобластов из миотомов в места закладки скелетных мышц; движение клеток из нервного гребня с образованием спинномозговых ганглиев и нервных сплетений, миграция гоноцитов и т. д.). Миграция осуществляется с помощью нескольких механизмов. Так, различают хемотаксис — движение клеток в направлении градиента концентрации какого-либо химического агента (перемещения спермиев к яйцеклетке, предшественников Т-лимфоцитов из костного мозга в закладку тимуса).
Читайте также: Какая ткань для штор лучше защищает от солнца
Гаптотаксис — механизм перемещения клеток по градиенту концентрации адгезионной молекулы (движение клеток протока пронефроса у амфибий по градиенту щелочной фосфатазы на поверхности мезодермы). Контактное ориентирование — когда в какой-либо преграде остается один канал для перемещения (описан у рыб при образовании плавников).
Контактное ингибирование — этот способ перемещения наблюдается у клеток нервного гребня. Суть способа заключается в том, что при образовании ламеллоподии одной клеткой и контакта ее с другой клеткой, ламеллоподия прекращает рост и постепенно исчезает, но в другой части мигрирующей клетки при этом формируется новая ламеллоподия.
В процессе миграции клеток важную роль играют межклеточные взаимодействия. Существует несколько механизмов такого взаимодействия (контактного и дистантного). Выделяется большая группа молекул клеточной адгезии (МКА). Так, кадгерины — это Са2+-зависимые МКА, отвечают за межклеточные контакты при образовании тканей, за формообразование и др. В молекуле кадгерина различают внеклеточный, трансмембранный и внутриклеточный домены. Например, внеклеточный домен ответственен за адгезию клеток с одинаковыми кадгеринами, а внутриклеточный — за форму клетки. Другой класс МКА — это иммуноглобулиновое суперсемейство Са2+-независимых МКА, обеспечивающих, например, адгезию аксонов к сарколемме мышечных волокон, или миграцию нейробластов вдоль радиальных глиоцитов в закладке коры большого мозга и др. Следующий класс МКА — это мембранные ферменты — гликозилтранферазы. Последние по типу «ключ-замок» соединяются с углеводными субстратами — гликозаминогликанами надмембранного комплекса клетки, осуществляя таким образом прочное сцепление клеток.
Кроме механизмов межклеточного взаимодействия, существуют механизмы взаимодействия клеток с субстратом. Они включают формирование рецепторов клетки к молекулам внеклеточного матрикса. К последним относят производные клеток, среди которых наиболее изученными адгезионными молекулами являются коллаген, фибронектин, ламинин, тенасцин и некоторые другие. Коллагены, среди которых различают несколько десятков типов, входят в состав межклеточного вещества рыхлой волокнистой соединительной ткани, базальной мембраны и пр. Фибронектин, секретируемый клетками, является связывающей молекулой между мигрирующей клеткой и межклеточным матриксом. Ламинин — компонент базальной мембраны, также связывает мигрирующие клетки с межклеточным матриксом (справедливо по отношению к эпителиоцитам и нейробластам).
Для осуществления связи мигрирующих клеток с межклеточным матриксом клетки формируют специфические рецепторы. К ним относятся, например, синдекан, который обеспечивает контакт эпителиоцита с базальной мембраной за счет сцепления с молекулами фибронектина и коллагена. Интегрины клеточных поверхностей связывают с внеклеточной стороны молекулы внеклеточного матрикса, а внутри клетки — белки цитоскелета (например, актиновые микрофиламенты). Так возникает связь внутри- и внеклеточных структур, что позволяет клетке использовать для перемещения собственный сократительный аппарат. Наконец, существует большая группа молекул, формирующих клеточные контакты, осуществляющие коммуникацию между клетками (щелевые контакты), механическую связь (десмосомы, плотные контакты).
Дистантные межклеточные взаимодействия осуществляются путем секреции гормонов и факторов роста (ФР). Последние — это вещества, оказывающие стимулирующее влияние на пролиферацию и дифференцировку клеток и тканей. К ним относятся, например, ФР, полученный из тромбоцитов и влияющий на переход клеток в фазу размножения (гладких миоцитов, фибробластов, глиоцитов); эпидермальный ФР — стимулирует пролиферацию эпителиоцитов, производных эктодермы; ФР фибробластов — стимулирует пролиферацию фибробластов. Особо выделяется большая группа пептидов (соматотропины, соматомедины, инсулин, лактоген), влияющих на развитие клеток плода.
Факторы, тормозящие пролиферацию и дифференцировку клеток, также принимают кооперативное участие в процессах развития клеток и тканей. К ним относятся, например, бета-интерферон и трансформирующий ФР. Последний, однако, в отношении разных клеточных типов действует по-разному: блокирует размножение многих типов эпителиоцитов, но стимулирует размножение соединительнотканных клеток.
— Вернуться в оглавление раздела «гистология»
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом