
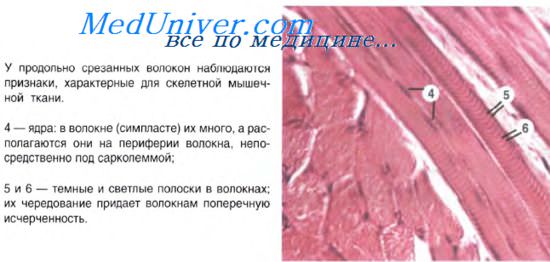
Структурно-функциональной единицей поперечнополосатой скелетной мышечной ткани является мышечное волокно. Волокно может достигать 12 см в длину, содержит большой объем саркоплазмы и сотни ядер. Каждое волокно покрыто сарколеммой, состоящей из двух слоев: внутреннего — плазмолеммы толщиной 8-10 нм и внешнего — базальной мембраны толщиной 30-40 нм. Между плазмолеммой и базальной мембраной имеется пространство шириной 15-25 нм. Кроме того, в базальную мембрану вплетаются ретикулярные волокна.
Значительный объем саркоплазмы занимают сократительные органеллы — миофибриллы. Каждая миофибрилла состоит из большого числа правильно чередующихся темных и светлых полос (дисков). В поляризованном свете темные диски обнаруживают двойное лучепреломление, поэтому называются анизотропными (А-дисками). Светлые диски таким свойством не обладают и называются изотропными (I-дисками). Каждая миофибрилла образована пучком параллельно идущих миофиламентов. А-диски состоят из толстых и тонких миофиламентов, а I-диски — только из тонких. Тонкие филаменты (5-8 нм) образованы белками актином, тропомиозином, тропонином, а толстые (10-12 нм) — миозином, белками М- и Н-полос и другими. Тонкие филаменты располагаются между толстыми, образуя гексагональное расположение.
Структурно-функциональной единицей миофибриллы является саркомер. Условная формула саркомера — 1/2 1-диска + А-диск + 1/2 I-диска. Линия сшивки соседних саркомеров соответствует Z-линии (телофрагме), которая состоит из белков альфа-актинина, десмина, вимен-тина. У позвоночных длина саркомера равна 2-3 мкм. Средняя часть миозинового диска, куда не доходят актиновые миофиламен-ты, более светлая и называется Н-полоской. Ее пересекает М-линия (мезофрагма), скрепляющая миозиновые нити посередине саркомера. В подмембранном слое сим-пласта обнаружены белки винкулин и спектрин, входящие в состав скелета симпласта.
Компоненты метаболической среды симпласта хорошо выражены. В гистогенезе с возрастанием степени зрелости симпластов наблюдается увеличение числа митохондрии, которые ориентируются по бокам Z-линии между миофибриллами и под сарколеммой. Гранулы гликогена, липидные капли формируют скопления между миофибриллами и под сарколеммой. Содержание миоглобина (связывающий кислород пигмент) варьирует в зависимости от образа жизни животного. Рибосомы представлены в виде полисом. Небольшое число лизосом принимают участие в процессах внутрисимпластической регенерации. Клеточный центр в симпласте отсутствует.
Саркоплазматическая сеть и Т-трубочки развиваются параллельно. Последние — это инвагинации плазмолеммы, которые опоясывают каждый саркомер. В продольном направлении вокруг каждой миофибриллы идут канальцы саркоплаз-матической сети. Так формируются продольная и поперечная системы, которые на срезах видны как триады. Триада — это комплекс, состоящий из поперечной трубочки и профилей двух цистерн саркоплазматической сети, расположенных симметрично по обе стороны от Т-трубочки. В цистернах саркоплазматической сети накапливаются ионы кальция, необходимые для сокращения миофибрилл.
В позднем онтогенезе происходит ряд ультраструктурных изменений в клетках и симпластах. Наиболее значимы — утолщение базальной мембраны, дезорганизация миофибрилл и Z-линии, возникновение скоплений митохондрий под сарколеммой, отделение миосателлитоцитов от симпласта и переход их в интерстициальное пространство. Иннервация мышечных волокон осуществляется двигательными нейронами передних рогов спинного мозга, которые формируют нервно-мышечные синапсы примерно в центральной части волокна.
Регенерация. Для успешной регенерации мышечной ткани необходимо сохранение напряжения мышцы, восстановление кровоснабжения и нервной связи. Основным источником регенерации являются миосателлитоциты. После активации последних происходит их митотическое деление, возникают миобласты, которые претерпевают дифференцировку, сливаются друг с другом и формируют симпласты. Развитие симпластов продолжается с участием размножающихся миосателлитоциов, часть которых сливается с растущими симпластами. Так формируются новые клеточно-симпластические системы — мышечные волокна.
Читайте также: Какие есть тянущиеся ткани
Гистология электронограммы мышечная ткань
Это ткань энтомезенхимного происхождения, которая делится на два вида: висцеральную и сосудистую. В эмбриональном гистогенезе даже электронно-микроскопически трудно отличить мезенхимные предшественники фибробластов от гладких миоцитов. В малодифференцированных гладких миоцитах развиты гранулярная эндоплазматическая сеть, комплекс Гольджи. Тонкие филаменты ориентированы вдоль длинной оси клетки. По мере развития размеры клетки и число филаментов в цитоплазме возрастают. Постепенно объем цитоплазмы, занятый сократительными филаментами, увеличивается, расположение их становится все более упорядоченным. Пролиферативная активность гладких миоцитов в миогенезе постепенно снижается. Это происходит в результате увеличения продолжительности клеточного цикла, выхода клеток из цикла репродукции и перехода в дифференцированное состояние.
Однако и в дефинитивном состоянии в гладкой мышечной ткани клеточная регенерация в виде размножения миоцитов полностью не прекращается. Существуют данные о том, что пролиферация и дифференцировка в большей степени свойственна субпопуляции малых (по размерам) гладких миоцитов.
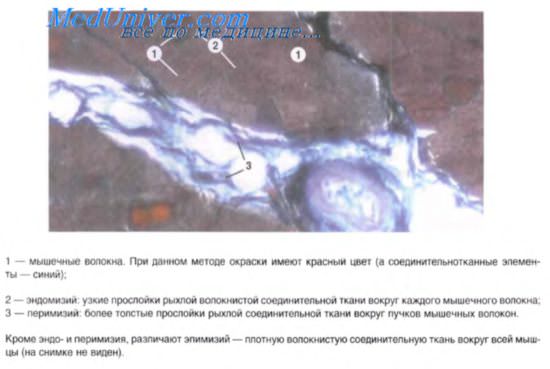
Строение гладкой мышечной ткани. Структура дефинитивных гладких миоцитов (лейомиоцитов), входящих в состав внутренних органов и стенки сосудов, имеет много общего, но в то же время характеризуется гетероморфией. Так, в стенках вен и артерий обнаруживаются овоидные, веретеновидные, отростчатые миоциты длиной 10-40 мкм, доходящие иногда до 140 мкм.
Гладкая мышечная ткань
Наибольшей длины гладкие миоциты достигают в стенке матки — до 500 мкм. Диаметр миоцитов колеблется от 2 до 20 мкм. В зависимости от характера внутриклеточных биосинтетических процессов различают контрактилъные и секреторные миоциты. Первые специализированы на функции сокращения, но вместе с тем сохраняют секреторную активность. Плазмолемма расслабленной клетки имеет ровную поверхность, а при сокращении становится складчатой. В центре клетки имеется палочковидное ядро, которое при сокращении клетки спиралевидно изгибается. Практически все ядра миоцитов содержат диплоидное количество ДНК. Гладкая эндоплазматическая сеть занимает примерно 2-7% объема цитоплазмы, а гранулярная сеть в контрактильных миоцитах выражена плохо. Митохондрии мелкие, сферические или овоидные, расположены у полюсов ядра. Характерной чертой гладких миоцитов является наличие множества впячиваний (кавеол) плазмолеммы, содержащих ионы кальция.
Секреторные миоциты (синтетические) по своей ультраструктуре напоминают фибробласты, однако содержат в цитоплазме пучки тонких миофиламентов, расположенные на периферии клетки. В цитоплазме хорошо развиты комплекс Гольджи, гранулярная эндоплазматическая сеть, много митохондрий, гранул гликогена, свободных рибосом и полисом. По степени зрелости такие клетки относят к малодифференцированным.
Сократительный аппарат миоцитов представлен тонкими актиновыми филамен-тами (гладкомышечным альфа-актином), связанными с тропомиозином. Толстые нити состоят из миозина, мономеры которого располагаются вблизи филаментов актина. Соотношение актиновых и миозиновых филаментов в гладком миоците составляет 12 к 1. Важным компонентом контрактильного аппарата миоцитов являются электронно-плотные структуры — тельца прикрепления, расположенные свободно в цитоплазме (плотные тельца) или тесно связанные с плазмолеммой. Основными белковыми компонентами плотных телец являются альфа-актинин, актин (немышечный) и кальпонин, что позволяет расссматривать их как функциональный эквивалент Z-линий миофибрилл скелетной мышцы. Актиновые филаменты фиксируются на плотных тельцах. Промежуточные филаменты, включающие десмин и виментин, обеспечивают связи между плотными тельцами и плазмолеммой, образуя прикрепительные пластины.
Читайте также: Ткани эконом в пскове
Сократительные белки формируют решетчатую структуру, закрепленную по окружности плазмолеммы, поэтому сокращение выражается в укорочении клетки, которая приобретает складчатую форму, тогда как в состоянии покоя клетка вытянута. При возникновении нервного импульса, распространяющегося по плазмолемме миоцита, происходит повышение уровня внутриклеточного Са2+, который поступает в цитоплазму из кавеол, отшнуровывающихся в цитоплазму в виде пузырьков. Высвобождение ионов кальция приводит к каскаду реакций, в результате которого происходит полимеризация миозина и образование перекрестных связей миозина вдоль актиновых филаментов по мере развития мышечного сокращения. Расслабление мышцы возникает при восстановлении концентрации исходного уровня Са2+ внутри клетки путем его перемещения внутрь саркоплазматической сети. При этом образовавшиеся в присутствии ионов кальция связи между актином и миозином нарушаются, акто-миозиновый комплекс распадается, гладкий миоцит расслабляется.
Гладкие миоциты синтезируют протеогликаны, гликопротеиды, проколлаген, проэластин, из которых формируются коллагеновые и эластические волокна и основное вещество межклеточного матрикса.
Взаимодействие миоцитов осуществляется с помощью цитоплазматических мостиков, взаимных впячиваний, нексусов, десмосом или простых участков мембранных контактов клеточных поверхностей.
Регенерация гладкой мышечной ткани
Гладкая мышечная ткань висцерального и сосудистого видов обладает значительной чувствительностью к воздействию экстремальных факторов.
В активированных миоцитах возрастает уровень биосинтетических процессов, морфологическим выражением которых являются синтез сократительных белков, укрупнение и гиперхроматоз ядра, гипертрофия ядрышка, возрастание показателей ядерно-цитоплазменного отношения, увеличение количества свободных рибосом и полисом, активация ферментов, аэробного и анаэробного фосфорилирования, мембранного транспорта. Клеточная регенерация осуществляется как за счет дифференцированных клеток, обладающих способностью вступать в митотический цикл, так и за счет активизации камбиальных элементов (миоцитов малого объема).
При действии ряда повреждающих факторов отмечается фенотипическая трансформация контрактильных миоцитов в секреторные. Данная трансформация часто наблюдается при повреждении интимы сосудов, формировании ее гиперплазии при развитии атеросклероза.
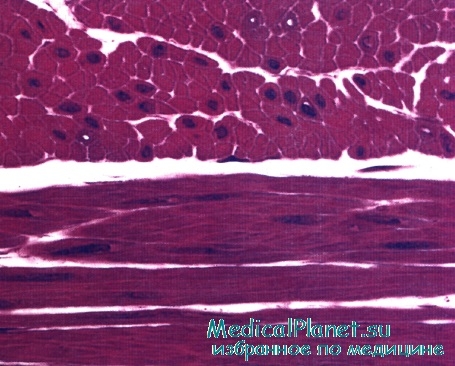
Гладкая мышечная ткань в поперечном (наверху) и продольном (внизу) разрезах. Обратите внимание на центрально расположенные ядра. Во многих клетках ядра не попали в срез.
Окраска: парарозанилин—толуидиновый синий. Среднее увеличение.
Гистология электронограммы мышечная ткань
Гистогенез сердечной мышечной ткани. Источники развития сердечной мышечной ткани находятся в прекардиальной мезодерме. В гистогенезе возникают парные складчатые утолщения висцерального листка спланхнотома — миоэпикардиальные пластинки, содержащие стволовые клетки сердечной мышечной ткани. Последние путем дивергентной дифференцировки дают начало следующим клеточным дифферонам: рабочим, ритмзадающим (пейсмекерным), проводящим и секреторным кардиомиоцитам.
Исходные клетки сердечной мышечной ткани — кардиомиобласты характеризуются рядом признаков: клетки уплощены, содержат крупное ядро, светлую цитоплазму, бедную рибосомами и митохондриями. В дальнейшем происходит развитие комплекса Гольджи, гранулярной эндоплазматической сети. В кардиомиобластах обнаруживаются фибриллярные структуры, но миофибрилл нет. Клетки обладают высоким пролиферативным потенциалом. После ряда митотических циклов кардиомиобласты дифференцируются в кардиомиоциты, в которых начинается саркомерогенез. В цитоплазме кардиомиоцитов увеличивается число полисом, канальцев гранулярной эндоплазматической сети, накапливаются гранулы гликогена, возрастает объем актомиозинового комплекса. Кардиомиоциты сокращаются, но не теряют способность к дальнейшей пролиферации и дифференцировке. Развитие сократительного аппарата в позднем эмбриональном и постнатальном периодах происходит путем надставки новых саркомеров и наслоения вновь синтезированных миофиламентов. Дифференцировка кардиомиоцитов сопровождается увеличением числа митохондрий, распределением их у полюсов ядер и между миофибриллами и протекает параллельно со специализацией контактирующих поверхностей клеток. Кардиомиоциты путем контактов «конец в конец», «конец в бок» формируют клеточные комплексы — сердечные мышечные волокна, и в целом ткань представляет собой сетевидную структуру.
Читайте также: Как из обрезков ткани сделать коврик из
Строение сердечной мышечной ткани.
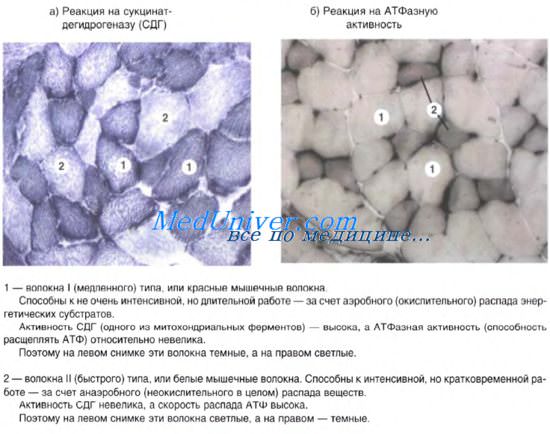
Структурно-функциональные единицы волокон — кардиомиоциты — это клетки, имеющие вытянутую прямоугольную форму. Длина рабочих кардиомиоцитов составляет 50-120 мкм, а ширина — 15-20 мкм. Одно-два ядра располагаются в центре клетки. Периферическую часть цитоплазмы кардиомиоцитов занимают поперечноисчерченные миофибриллы, аналогичные таковым в симпластах скелетномышечного волокна. Однако каналы саркоплазматической сети и Т-системы менее отчетливо выражены. Кардиомиоциты отличаются большим количеством митохондрий, расположенных тесными рядами между миофибриллами. Снаружи миоциты покрыты сарколеммой, в составе которой выделяются плазмолемма и базальная мембрана. Характерной особенностью ткани является наличие вставочных дисков на границе между контактирующими кардиомиоцитами. Вставочные диски пересекают волокно в виде волнистой или ступенчатой линии и включают межклеточные контакты от простых, по типу десмо-сом и до щелевых (нексусов).
Часть кардиомиоцитов на ранних этапах кардиомиогенеза являются сократительно-секреторными. В дальнейшем в результате дивергентной дифференцировки возникают «темные» (сократительные) и «светлые» (проводящие) миоциты, в которых исчезают секреторные гранулы, тогда как в предсердных миоцитах они сохраняются. Так формируется дифферон эндокринных кардиомиоцитов. Эти клетки содержат центрально расположенное ядро с диспергированным хроматином,
1-2 ядрышками. В цитоплазме хорошо развиты гранулярная эндоплазматическая сеть, диктиосомы комплекса Гольджи, в тесной связи с элементами которого находятся многочисленные секреторные гранулы диаметром около 2 мкм, содержащие электронноплотный материал. В дальнейшем секреторные гранулы обнаруживаются под сарколеммой и выделяются в межклеточное пространство путем экзоцитоза. Выделенный пептидный гормон кардиодилатин циркулирует в крови в виде кардионатрина, который вызывает сокращение гладких миоцитов артериол, увеличение почечного кровотока, ускоряет клубочковую фильтрацию и выделение натрия из организма.
Кардиомиоциты проводящей системы гетероморфны. В них слабо развит мио-фибриллярный аппарат, расположение миофиламентов в составе миофибрилл рыхлое, Z-линии имеют неправильную конфигурацию, эндоплазматическая сеть слабо развита, находится на периферии миоцитов, число митохондрий незначительное. По мере расположения этих кардиомиоцитов в проксимо-дистальном направлении соответственно движению импульсов от синусно-предсердного узла, через предсердно-желудочковый узел, пучок Гиса, его ножки и клетки Пуркиня к рабочим миоцитам проводящие кардиомиоциты по своей ультраструктуре приближаются к рабочим кардиомиоцитам.
Регенерация сердечной мышечной ткани.
В гистогенезе сердечной мышечной ткани специализированный камбий не возникает. Поэтому регенерация ткани протекает на основе внутриклеточных гиперпластических процессов. Вместе с тем для кардиомиоцитов млекопитающих, приматов и человека характерен процесс полиплоидизации. Например, у обезьян ядра до 50% терминально дифференцированных кардиомиоцитов становятся тетра- и октоплоидными. Полиплоидные кардиомиоциты возникают за счет ацитокинетического митоза, что приводит к многоядерности.
В условиях патологии сердечно-сосудистой системы человека (ревматизм, врожденные пороки сердца, инфаркт миокарда и другие) важная роль в компенсации повреждений кардиомиоцитов принадлежит внутриклеточной регенерации, полиплоидизации как ядер, так и кардиомиоцитов.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом