
Это ткань энтомезенхимного происхождения, которая делится на два вида: висцеральную и сосудистую. В эмбриональном гистогенезе даже электронно-микроскопически трудно отличить мезенхимные предшественники фибробластов от гладких миоцитов. В малодифференцированных гладких миоцитах развиты гранулярная эндоплазматическая сеть, комплекс Гольджи. Тонкие филаменты ориентированы вдоль длинной оси клетки. По мере развития размеры клетки и число филаментов в цитоплазме возрастают. Постепенно объем цитоплазмы, занятый сократительными филаментами, увеличивается, расположение их становится все более упорядоченным. Пролиферативная активность гладких миоцитов в миогенезе постепенно снижается. Это происходит в результате увеличения продолжительности клеточного цикла, выхода клеток из цикла репродукции и перехода в дифференцированное состояние.
Однако и в дефинитивном состоянии в гладкой мышечной ткани клеточная регенерация в виде размножения миоцитов полностью не прекращается. Существуют данные о том, что пролиферация и дифференцировка в большей степени свойственна субпопуляции малых (по размерам) гладких миоцитов.
Строение гладкой мышечной ткани. Структура дефинитивных гладких миоцитов (лейомиоцитов), входящих в состав внутренних органов и стенки сосудов, имеет много общего, но в то же время характеризуется гетероморфией. Так, в стенках вен и артерий обнаруживаются овоидные, веретеновидные, отростчатые миоциты длиной 10-40 мкм, доходящие иногда до 140 мкм.
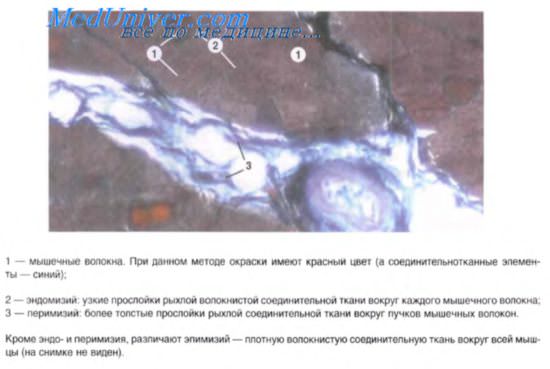
Гладкая мышечная ткань
Наибольшей длины гладкие миоциты достигают в стенке матки — до 500 мкм. Диаметр миоцитов колеблется от 2 до 20 мкм. В зависимости от характера внутриклеточных биосинтетических процессов различают контрактилъные и секреторные миоциты. Первые специализированы на функции сокращения, но вместе с тем сохраняют секреторную активность. Плазмолемма расслабленной клетки имеет ровную поверхность, а при сокращении становится складчатой. В центре клетки имеется палочковидное ядро, которое при сокращении клетки спиралевидно изгибается. Практически все ядра миоцитов содержат диплоидное количество ДНК. Гладкая эндоплазматическая сеть занимает примерно 2-7% объема цитоплазмы, а гранулярная сеть в контрактильных миоцитах выражена плохо. Митохондрии мелкие, сферические или овоидные, расположены у полюсов ядра. Характерной чертой гладких миоцитов является наличие множества впячиваний (кавеол) плазмолеммы, содержащих ионы кальция.
Секреторные миоциты (синтетические) по своей ультраструктуре напоминают фибробласты, однако содержат в цитоплазме пучки тонких миофиламентов, расположенные на периферии клетки. В цитоплазме хорошо развиты комплекс Гольджи, гранулярная эндоплазматическая сеть, много митохондрий, гранул гликогена, свободных рибосом и полисом. По степени зрелости такие клетки относят к малодифференцированным.
Сократительный аппарат миоцитов представлен тонкими актиновыми филамен-тами (гладкомышечным альфа-актином), связанными с тропомиозином. Толстые нити состоят из миозина, мономеры которого располагаются вблизи филаментов актина. Соотношение актиновых и миозиновых филаментов в гладком миоците составляет 12 к 1. Важным компонентом контрактильного аппарата миоцитов являются электронно-плотные структуры — тельца прикрепления, расположенные свободно в цитоплазме (плотные тельца) или тесно связанные с плазмолеммой. Основными белковыми компонентами плотных телец являются альфа-актинин, актин (немышечный) и кальпонин, что позволяет расссматривать их как функциональный эквивалент Z-линий миофибрилл скелетной мышцы. Актиновые филаменты фиксируются на плотных тельцах. Промежуточные филаменты, включающие десмин и виментин, обеспечивают связи между плотными тельцами и плазмолеммой, образуя прикрепительные пластины.
Сократительные белки формируют решетчатую структуру, закрепленную по окружности плазмолеммы, поэтому сокращение выражается в укорочении клетки, которая приобретает складчатую форму, тогда как в состоянии покоя клетка вытянута. При возникновении нервного импульса, распространяющегося по плазмолемме миоцита, происходит повышение уровня внутриклеточного Са2+, который поступает в цитоплазму из кавеол, отшнуровывающихся в цитоплазму в виде пузырьков. Высвобождение ионов кальция приводит к каскаду реакций, в результате которого происходит полимеризация миозина и образование перекрестных связей миозина вдоль актиновых филаментов по мере развития мышечного сокращения. Расслабление мышцы возникает при восстановлении концентрации исходного уровня Са2+ внутри клетки путем его перемещения внутрь саркоплазматической сети. При этом образовавшиеся в присутствии ионов кальция связи между актином и миозином нарушаются, акто-миозиновый комплекс распадается, гладкий миоцит расслабляется.
Гладкие миоциты синтезируют протеогликаны, гликопротеиды, проколлаген, проэластин, из которых формируются коллагеновые и эластические волокна и основное вещество межклеточного матрикса.
Взаимодействие миоцитов осуществляется с помощью цитоплазматических мостиков, взаимных впячиваний, нексусов, десмосом или простых участков мембранных контактов клеточных поверхностей.
Регенерация гладкой мышечной ткани
Гладкая мышечная ткань висцерального и сосудистого видов обладает значительной чувствительностью к воздействию экстремальных факторов.
В активированных миоцитах возрастает уровень биосинтетических процессов, морфологическим выражением которых являются синтез сократительных белков, укрупнение и гиперхроматоз ядра, гипертрофия ядрышка, возрастание показателей ядерно-цитоплазменного отношения, увеличение количества свободных рибосом и полисом, активация ферментов, аэробного и анаэробного фосфорилирования, мембранного транспорта. Клеточная регенерация осуществляется как за счет дифференцированных клеток, обладающих способностью вступать в митотический цикл, так и за счет активизации камбиальных элементов (миоцитов малого объема).
При действии ряда повреждающих факторов отмечается фенотипическая трансформация контрактильных миоцитов в секреторные. Данная трансформация часто наблюдается при повреждении интимы сосудов, формировании ее гиперплазии при развитии атеросклероза.
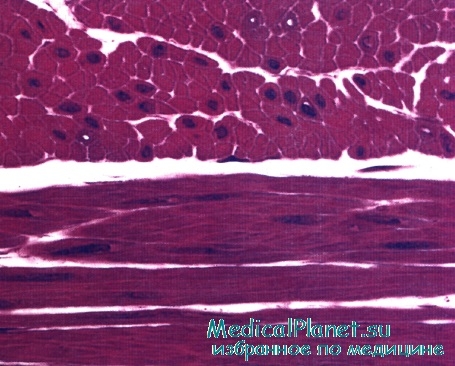
Гладкая мышечная ткань в поперечном (наверху) и продольном (внизу) разрезах. Обратите внимание на центрально расположенные ядра. Во многих клетках ядра не попали в срез.
Окраска: парарозанилин—толуидиновый синий. Среднее увеличение.
Гладкая мышечная ткань это эндотелий

Эндотелий — активный эндокринный орган, самый большой в организме, диффузно рассеянный вместе с сосудами по всем тканям. Эндотелий, по классическому определению гистологов, — однослойный пласт специализированных клеток, выстилающих изнутри все сердечно-сосудистое дерево, весом около 1,8 кг. Один триллион клеток со сложнейшими биохимическими функциями, включающий системы синтеза белков и низкомолекулярных веществ, рецепторы, ионные каналы [1].
Эндотелиоциты синтезируют субстанции, важные для контроля свертывания крови, регуляции сосудистого тонуса, артериального давления, фильтрационной функции почек, сократительной активности сердца, метаболического обеспечения мозга. Эндотелий способен реагировать на механическое воздействие протекающей крови, величину давления крови в просвете сосуда и степень напряжения мышечного слоя сосуда. Клетки эндотелия чувствительны к химическим воздействиям, которые могут приводить к повышенной агрегации и адгезии циркулирующих клеток крови, развитию тромбоза, оседанию липидных конгломератов (табл. 1).
Все эндотелиальные факторы делятся на вызывающие сокращение и расслабление мышечного слоя сосудистой стенки (констрикторы и дилятаторы). Основные констрикторы представлены ниже.
Большой эндотелин — неактивный предшественник эндотелина, содержащий 38 аминокислотных остатков, обладает менее выраженной вазоконстрикторной (по сравнению с эндотелином) активностью in vitro. Конечный процессинг большого эндотелина осуществляется при участии эндотелинпревращающего фермента.
Эндотелин (ЭТ). Японский исследователь М. Янагасава и соавт. (1988) описали новый эндотелиальный пептид, активно сокращающий гладкомышечные клетки сосудов. Открытый пептид, названный ЭТ, сразу стал предметом интенсивного изучения. ЭТ- сегодня один из самых популярных в списке биоактивных регуляторов. Это — вещество с наиболее мощной сосудосуживающей активностью образуется в эндотелии. В организме присутствуют несколько форм пептида, различающихся небольшими нюансами химического строения, но весьма не схожих по локализации в организме и физиологической активности. Синтез ЭТ стимулируют тромбин, адреналин, ангиотензин (АТ), интерлейкины, клеточные ростовые факторы и др. В большинстве случаев ЭТ секретируется из эндотелия «внутрь», к мышечным клеткам, где расположены чувствительные к нему ЕТА-рецепторы. Меньшая часть синтезируемого пептида, взаимодействуя с рецепторами ЕТВ-типа, стимулирует синтез NO. Таким образом, один и тот же фактор регулирует две противоположные сосудистые реакции (констрикцию и дилятацию), реализуемые различными химическими механизмами.
Читайте также: Как покрасить грубую ткань
Факторы, синтезируемые в эндотелии и регулирующие его функцию
Факторы, вызывающие сокращение и расслабление мышечного слоя сосудистой стенки
Эндотелиновый фактор деполяризации (EDHF)
Факторы прогоагуляционные и антикоагуляционные
Тромбоцитарный фактор роста (ТФРβ)
Ингибитор тканевого активатора плазминогена (ИТАП)
Тканевой активатор плазминогена (ТАП)
Фактор Виллебранда (VIII фактор свертывания)
Фактор активации тромбоцитов (ФАТ)
Факторы, влияющие на рост сосудов и гладкомышечных клеток
Натриуретический пептид С
Эндотелиальный фактор роста (ECGF)
Гепариноподобные ингибиторы роста
Факторы провоспалительные и противовоспалительные
Фактор некроза опухоли α (ФНО-α)
Для ЭТ выявлены подтипы рецепторов, не схожие по клеточной локализации и запускающие «сигнальные» биохимические реакции. Четко прослеживается биологическая закономерность, когда одно и то же вещество, в частности, ЭТ регулирует различные физиологические процессы (табл. 2).
ЭТ — это группа полипептидов, состоящая из трех изомеров (ЭТ-1, ЭТ-2, ЭТ-3), отличающихся некоторыми вариациями и последовательностью расположения аминокислот. Имеется большое сходство между структурой ЭТ и некоторыми нейротоксическими пептидами (яды скорпиона, роющей змеи).
Основной механизм действия всех ЭТ заключается в увеличении содержания в цитоплазме гладкомышечных клеток сосудов ионов кальция, что вызывает:
- стимуляцию всех фаз гемостаза, начиная с агрегации тромбоцитов и заканчивая образованием красного тромба;
- сокращение и рост гладких мышц сосудов, приводящие к вазоконстрикции и утолщению стенки сосудов и уменьшению их диаметра.
Подтипы рецепторов ЭТ: локализация, физиологические эффекты
и участие вторичных посредников
Высвобождение NO, PGI2, EDGF
Высвобождение NO и др. факторы
Эффекты ЭТ неоднозначны и определяются рядом причин. Наиболее активен изомер — ЭТ-1. Он образуется не только в эндотелии, но и в гладких мышцах сосудов, нейронах, глие, мезенгиальных клетках почек, печени и других органах. Полупериод жизни — 10-20 мин, в плазме крови — 4-7 мин. ЭТ-1 причастен к ряду патологических процессов: инфаркту миокарда, нарушению ритма сердца, легочной и системной гипертезии, атеросклерозу и др. [6, 9].
Поврежденный эндотелий синтезирует большое количество ЭТ, вызывающего вазоконстрикцию [7]. Большие дозы ЭТ приводят к значительным изменениям системной гемодинамики: снижению частоты сердечных сокращений и ударного объема сердца, увеличению на 50 % сосудистого сопротивления в большом круге кровообращения и на 130 % в малом [7].
Ангиотензин II (AT II) — физиологически активный пептид прогипертензивного действия. Это гормон, образующийся в крови человека при активации ренин-ангиотензиновой системы, участвует в регуляции артериального давления и водно-солевого обмена. Этот гормон вызывает сужение выносящих артериол почечных клубочков [35]. Он увеличивает реабсорбцию в почечных канальцах натрия и воды. АТ II суживает артерии и вены, а также стимулирует выработку таких гормонов, как вазопрессин и альдостерон, что приводит к повышению давления. Сосудосуживающая активность АТ II определяется его взаимодействием с AT I рецептором [10].
Тромбоксан А2 (ТхА2) — способствует быстрой агрегации тромбоцитов, увеличивая доступность их рецепторов для фибриногена, чем активирует коагуляцию, вызывает вазоспазм и бронхоспазм. Кроме того, TхA2 является медиатором в опухолеобразовании, тромбозах и астме. ТхА2 вырабатывается также гладкими мышцами сосудов, тромбоцитами. Одним из факторов, стимулирующих выделение ТхА2, является кальций, который в большом количестве выделяется из тромбоцитов в начале их агрегации. ТхА2 сам увеличивает содержание кальция в цитоплазме тромбоцитов. Кроме того, кальций активирует сократительные белки тромбоцитов, что усиливает их агрегацию и дегрануляцию. Он активирует фосфолипазу А2, превращающую арахидоновую кислоту в простагландины G2, Н2 — вазоконстрикторы [7].
Простагландин H2 (PGH2) — обладает ярко выраженной биологической активностью. Он стимулирует агрегацию тромбоцитов и вызывает сокращение гладких мышц с формированием вазоспазма.
Группа веществ под названием дилятаторы, представлена следующими биологически активными веществами.
Оксид азота (NO) — это низкомолекулярная и не несущая заряда молекула, способная быстро диффундировать и свободно проникать через плотные клеточные слои и межклеточное пространство. По строению NO содержит неспаренный электрон, имеет высокую химическую активность и легко реагирует со многими клеточными структурами и химическими компонентами, что обусловливает исключительное многообразие ее биологических эффектов. NO способен вызывать различные и даже противоположные эффекты в клетках-мишенях, что зависит от наличия дополнительных факторов: окислительно-восстановительного и пролиферативного статуса и ряда прочих условий. NO влияет на эффекторные системы, контролирующие пролиферацию, апоптоз и дифференцировку клеток, а также на их устойчивость к стрессовым воздействиям. NO выполняет функции посредника в передаче паракринного сигнала. Действие NO вызывает быстрый и относительно кратковременный ответ в клетках-мишенях, обусловленный снижением уровня кальция, а также долговременные эффекты, обусловленные индукцией определенных генов. В клетках-мишенях NO и ее активные производные, такие как пероксинитрит, действуют на белки, содержащие гем, железосерные центры и активные тиолы, также ингибируют железосерные ферменты. Кроме того, NO рассматривают как один из мессенджеров внутри и межклеточной сигнализации в центральной и периферической нервной системе [3] и рассматривают как регулятор пролиферации лимфоцитов. Эндогенный NO — важный компонент системы регуляции кальциевого гомеостаза в клетках [4] и соответственно активности Са 2+ -зависимых протеинкиназ. Образование NO в организме происходит при ферментативном окислении L-аргинина. Синтез NO осуществляется семейством цитохром ‒ P-450-подобных гемопротеинов — NO-синтаз.
По определению ряда исследователей — NO — «двуликий Янус»:
- NO как усиливает процессы перекисного окисления липидов (ПОЛ) в мембранах клеток и липопротеинах сыворотки, так и ингибирует их;
- NO вызывает вазодилятацию, но может вызывать и вазоконстрикцию [2];
- NO индуцирует апоптоз, но оказывает защитный эффект в отношении апоптоза, индуцированного другими агентами;
- NO способен модулировать развитие воспалительной реакции [2] и ингибировать окислительное фосфорилирование в митохондриях и синтез АТФ [5].
Читайте также: Боль в тканях при герпесе
Простациклин (PGI2) — образуется преимущественно в эндотелии. Синтез простациклина происходит постоянно. Он подавляет агрегацию тромбоцитов, кроме того, оказывает вазодилятирующее действие за счет стимуляции специфических рецепторов гладкомышечных клеток сосудов, что приводит к повышению активности в них аденилатциклазы и к увеличению образования в них цАМФ.
Эндотелий зависимый гиперполяризующий фактор (EDHF) — по своей структуре он не идентифицирован, как NO или простациклин. EDHF вызывает гиперполяризацию гладкомышечного слоя артериальной стенки и соответственно его релаксацию. G. Edwards и соавт. (1998) было установлено, что EDHF не что иное как К+, который выделяется эндотелиоцитами в миоэндотелиальное пространство стенки артерии при действии на последнюю адекватного раздражителя. EDHF способен играть важную роль в регуляции артериального давления.
Адреномедулин содержится в сосудистой стенке, обоих предсердиях и желудочках сердца, спинномозговой жидкости. Имеются указания на то, что адреномедулин может синтезироваться легкими и почками. Адреномедулин стимулирует продукцию эндотелием NO, что способствует вазодилятации, расширяет сосуды почек и увеличивает скорость клубочковой фильтрации и диурез, повышает натрийурез, снижает пролиферацию гладкомышечных клеток, препятствует развитию гипертрофии и ремоделирования миокарда и сосудов, ингибирует синтез альдостерона и ЭТ.
Следующая функция сосудистого эндотелия — участие в реакциях гемостаза за счет выделения протромбогенных и антитромбогенных факторов.
Группа протромбогенных факторов представлена следующими агентами.
Тромбоцитарный фактор роста (PDGF) является наиболее хорошо изученным представителем группы белковых факторов роста. PDGF может изменять пролиферативный статус клетки, влияя на интенсивность белкового синтеза, но, не затрагивая при этом усиления транскрипции генов раннего ответа, как c-myc и c-fos. Сами тромбоциты не синтезируют белок. Синтез и процессинг PDGF осуществляется в мегакариоцитах — клетках костного мозга, предшественниках тромбоцитов — и запасается в α-гранулах тромбоцитов. Пока PDGF находится внутри тромбоцитов, он недоступен для других клеток, однако при взаимодействии с тромбином происходит активация тромбоцитов с последующим высвобождением содержимого в сыворотку. Тромбоциты являются главным источником PDGF в организме, но вместе с тем показано, что некоторые другие клетки также могут синтезировать и секретировать этот фактор: это в основном клетки мезенхимального происхождения.
Ингибитор тканевого активатора плазминогена-1 (ИТАП-1) — продуцируется эндотелиоцитами, клетками гладких мышц, мегакариоцитами и мезотелиальными клетками; депонируется в тромбоцитах в неактивной форме и является серпином. Уровень ИТАП-1 в крови регулируется очень точно и возрастает при многих патологических состояниях. Его продукция стимулируется тромбином, трансформирующим фактором роста β, тромбоцитарным фактором роста, ИЛ-1, ФНО-α, инсулиноподобным фактором роста, глюкокортикоидами. Основная функция ИТАП-1 — ограничить фибринолитическую активность местом расположения гемостатической пробки за счет ингибирования ТАП. Это выполняется легко за счет большего содержания его в сосудистой стенке по сравнению с тканевым активатором плазминогена. Таким образом, на месте повреждения активированные тромбоциты выделяют избыточное количество ИТАП-1, предотвращая преждевременный лизис фибрина.
Ингибитор тканевого активатора плазминогена 2 (ИТАП-2) — основной ингибитор урокиназы.
Фактор фон Виллебранда (VIII — vWF) — синтезируется в эндотелии и мегакариоцитах; стимулирует начало тромбообразования: способствует прикреплению рецепторов тромбоцитов к коллагену и фибронектину сосудов, усиливает адгезию и агрегацию тромбоцитов. Синтез и выделение этого фактора возрастает под влиянием вазопрессина, при повреждении эндотелия. Поскольку все стрессорные состояния увеличивают выделение вазопрессина, то при стрессах, экстремальных состояниях тромбогенность сосудов возрастает.
АТ II быстро метаболизируется (период полураспада — 12 мин) при участии аминопептидазы А с образованием АТ III и далее под влиянием аминопептидазы N — ангиотензина IV, обладающих биологической активностью. АТ IV, предположительно, участвует в регуляции гемостаза, опосредует угнетение клубочковой фильтрации.
Важная роль отводится фибронектину — гликопротеиду, состоящему из двух цепей, соединенных дисульфидными связями. Вырабатывается он всеми клетками сосудистой стенки, тромбоцитами. Фибронектин является рецептором для фибринстабилизирующего фактора. Способствует адгезии тромбоцитов, участвуя в образовании белого тромба; связывает гепарин. Присоединяясь к фибрину, фибронектин уплотняет тромб. Под действием фибронектина клетки гладких мышц, эпителиоцитов, фибробластов повышают свою чувствительность к факторам роста, что может вызвать утолщение мышечной стенки сосудов и повышение общего периферического сопротивления сосудов.
Тромбоспондин — гликопротеид, который не только вырабатывается эндотелием сосудов, но находится и в тромбоцитах. Он образует комплексы с коллагеном, гепарином, являясь сильным агрегирующим фактором, опосредующим адгезию тромбоцитов к субэндотелию.
Фактор активации тромбоцитов (ФАТ) — образуется в различных клетках (лейкоциты, эндотелиальные клетки, тучные клетки, нейтрофилы, моноциты, макрофаги, эозинофилы и тромбоциты), относится к веществам с сильным биологическим действием.
ФАТ задействован в патогенезе аллергических реакций немедленного типа. Он стимулирует агрегацию тромбоцитов с последующей активацией фактора XII (фактора Хагемана). Активированный фактор XII, в свою очередь, активирует образование кининов, наибольшее значение из которых имеет брадикинин.
Группа антитромбогенных факторов представлена нижеперечисленными биологически активными веществами.
Тканевой активатор плазминогена (tPA, фактор III, тромбопластин, ТАП) — сериновая протеаза катализирует превращение неактивного профермента плазминогена в активный фермент плазмин и является важным компонентом системы фибринолиза. ТАП является одним из ферментов, наиболее часто вовлекаемых в процессы деструкции базальной мембраны, внеклеточного матрикса и инвазии клеток. Он продуцируется эндотелием и локализован в стенке сосудов. ТАП представляет собой фосфолипопротеин, эндотелиальный активатор, высвобождаемый в кровоток под действием разных стимулов.
Основные функции сводятся к инициации активации внешнего механизма свертывания крови. Он обладает высоким сродством к циркулирующему в крови ф.VII. В присутствии ионов Са2+ ТАП образует комплекс с ф.VII, вызывая его конформационные изменения и превращая последний в сериновую протеиназу ф.VIIа. Возникающий комплекс (ф.VIIа-Т.ф.) превращает ф.Х в сериновую протеиназу ф.Ха. Комплекс ТАП-фактор VII способен активировать как фактор X, так и фактор IX, что, в конечном итоге, способствует образованию тромбина.
Тромбомодулин — протеогликан, содержащийся в сосудах и являющийся рецептором для тромбина. Эквимолярный комплекс тромбин-тромбомодулин не вызывает превращения фибриногена в фибрин, ускоряет инактивацию тромбина антитромбином III и активирует протеин C, один из физиологических антикоагулянтов крови (ингибиторов свертывания крови). В комплексе с тромбином тромбомодулин функционирует в качестве кофактора. Связанный с тромбомодулином тромбин в результате изменения конформации активного центра приобретает повышенную чувствительность в отношении инактивации его антитромбином III и полностью теряет способность взаимодействовать с фибриногеном и активировать тромбоциты.
Жидкое состояние крови поддерживается благодаря ее движению, адсорбции факторов свертывания эндотелием и, наконец, благодаря естественным антикоагулянтам. Важнейшие из них — это антитромбин III, протеин С, протеин S и ингибитор внешнего механизма свертывания.
Читайте также: Сколько в рулоне вафельной ткани
Антитромбин III (АТ III) — нейтрализует активность тромбина и других активированных факторов свертывания крови (фактора XIIa, фактора XIa, фактора Xa и фактора IXa). В отсутствие гепарина комплексирование АТ III с тромбином протекает медленно. При связывании остатков лизина АТ III с гепарином в ее молекуле происходят конформационные сдвиги, способствующие быстрому взаимодействию реактивного места АТ III с активным центром тромбина. Это свойство гепарина лежит в основе его антикоагулянтного действия. АТ III образует комплексы с активированными факторами свертывания крови, блокируя их действие. Эта реакция в сосудистой стенке и на эндотелиальных клетках ускоряется гепариноподобными молекулами.
Протеин С — синтезируемый в печени витамин-К-зависимый белок, который связывается с тромбомодулином и превращается тромбином в активную протеазу. Взаимодействуя с протеином S, активированный протеин С разрушает фактор Va и фактор VIIIa, прекращая образование фибрина. Активированный протеин С может также стимулировать фибринолиз. Уровень протеина С не столь жестко связан с наклонностью к тромбозам, как уровень АТ III. Кроме того, протеин С стимулирует выделение тканевого активатора плазминогена эндотелиальными клетками. Кофактором протеина С служит протеин S.
Протеин S — фактор протромбинового комплекса, кофактор протеина С. Снижение уровня АТ III, протеина С и протеина S или их структурные аномалии ведут к повышению свертываемости крови. Протеин S — витамин К — зависимый одноцепочечный плазменный протеин, является кофактором активированного протеина С, вместе с которым регулирует скорость свертывания крови. Протеин S синтезируется в гепатоцитах, эндотелиальных клетках мегакариоцитах, клетках Лейдинга, а также в клетках мозга. Протеин S функционирует как неэнзиматический кофактор активированного белка C, сериновая протеаза, участвующая в протеолитической деградации факторов Va и VIIIa.
Все факторы, влияющие на рост сосудов и гладкомышечных клеток, делятся на стимуляторы и ингибиторы. Основные стимуляторы представлены ниже.
Ключевой активной формой кислорода является супероксид анион-радикал (Ō2), образующийся при присоединении одного электрона к молекуле кислорода в основном состоянии. Ō2 представляет опасность тем, что способен повреждать белки, содержащие железо-серные кластеры, такие как аконитаза, сукцинатдегидрогеназа и НАДН-убихинон оксидоредуктаза. При кислых значениях рН Ō2 может протонироваться с образованием более реакционноспособного пероксидного радикала. Присоединение двух электронов к молекуле кислорода или одного электрона к Ō2 приводит к образованию Н2О2, которая является окислителем умеренной силы.
Опасность любых реакционно-активных соединений в значительной степени зависит от их стабильности. Экзогенно возникшие Ō2 могут проникать в клетку и (наряду с эндогенными) участвовать в реакциях, приводящих к различным повреждениям: перекисном окислении ненасыщенных жирных кислот, окислении SH-групп белков, повреждении ДНК и др.
Фактор роста эндотелиальных клеток (beta-Endothelial Cell Growth Factor) — обладает свойствами ростового фактора эндотелиальных клеток. 50 % аминокислотной последовательности молекулы ECGF соответствует структуре фактора роста фибробластов (FGF). Оба эти пептида также обнаруживают сходную аффинность к гепарину и ангиогенную активность in vivo. Основной фактор роста фибробластов (bFGF) считается одним из важных индукторов опухолевого ангиогенеза.
Главные ингибиторы роста сосудов и гладкомышечных клеток представлены следующими веществами.
Эндотелиальный натрийуретический пептид С — вырабатывается, главным образом, в эндотелии, но обнаруживается также в миокарде предсердий, желудочков и в почках. Вазоактивным действием обладает CNP, выделяющийся из эндотелиальных клеток и паракринно воздействующий на рецепторы гладкомышечных клеток, вызывая и вазодилятацию. Синтез CNP усиливается в условиях дефицита NO, что имеет компенсаторное значение при развитии артериальной гипертензии и атеросклерозе.
Макроглобулин α2 — это гликопротеин, который относится к α2-глобулинам и представляет собой одну полипептидную цепь с молекулярной массой 725000 кДа. Нейтрализует плазмин, оставшийся неинактивированным после взаимодействия с α2-антиплазмином. Угнетает активность тромбина.
Кофактор II гепарина — гликопротеин, одноцепочечный полипептид с молекулярной массой 65000 кДа. Его концентрация в крови равна 90 мкг/мл. Инактивирует тромбин, образуя с ним комплекс. Реакция значительно ускоряется в присутствии дерматансульфата.
Сосудистый эндотелий также вырабатывает факторы, влияющие на развитие и течение воспаления.
Они делятся на провоспалительные и противовоспалительные. Ниже представлены провоспалительные факторы.
Фактор некроза опухоли-α (ФНО-α, кахектин) — это пироген, во многом дублирует действие ИЛ-1, но кроме того, играет важную роль в патогенезе септического шока, вызванного грамотрицательными бактериями. Под влиянием ФНО-α резко увеличивается образование макрофагами и нейтрофилами Н2О2 и других свободных радикалов. При хроническом воспалении ФНО-α активирует катаболические процессы и тем самым способствует развитию кахексии.
Цитотоксическое действие ФНО-α на опухолевую клетку связано с деградацией ДНК и нарушением функционирования митохондрий.
Индикатором эндотелиальной дисфункции может служить С-реактивный белок (С-РБ). Накоплено достаточно сведений о взаимосвязи С-РБ с развитием поражений сосудистой стенки и его непосредственном участии в этом процессе. Ввиду этого уровень С-РБ рассматривается сегодня в качестве надежного предиктора осложнений сосудистых заболеваний мозга (инсульт), сердца (инфаркт), периферических сосудистых нарушений. С-РБ опосредует инициальные стадии повреждения сосудистой стенки: активацию эндотелиальных молекул адгезии (ICAM-l, VCAM-l), секрецию хемотаксических и провоспалительных факторов (МСР-1 — хемотаксический для макрофагов белок, ИЛ-6), способствуя привлечению и адгезии иммунных клеток к эндотелию. Об участии С-РБ в повреждении сосудистой стенки свидетельствуют, кроме того, и данные о депозитах С-РБ, обнаруженных в стенках пораженных сосудов при инфаркте миокарда, атеросклерозе, васкулитах.
Основной противовоспалительный фактор — оксид азота (его функции представлены выше).
Таким образом, сосудистый эндотелий, находясь на границе между кровью и другими тканями организма, полностью выполняет свои основные функции за счет биологически активных веществ: регуляция параметров гемодинамики, тромборезистентность и участие в процессах гемостаза, участие в воспалении и ангиогенезе.
При нарушении функции или структуры эндотелия резко меняется спектр выделяемых им биологически активных веществ. Эндотелий начинает секретировать агреганты, коагулянты, вазоконстрикторы, причем часть из них (ренин-ангиотензиновая система) [10] оказывает влияние на всю сердечно-сосудистую систему. При неблагоприятных условиях (гипоксия, нарушения обмена веществ, атеросклероз и т. п.) эндотелий становится инициатором (или модулятором) многих патологических процессов в организме [6, 7, 8, 9].
Бердичевская Е.М., д.м.н., профессор, зав. кафедрой физиологии ФГОУ ВПО «Кубанский государственный университет физической культуры, спорта и туризма» г. Краснодар;
Быков И.М., д.м.н., профессор, зав. кафедрой фундаментальной и клинической биохимии ГБОУ ВПО КубГМУ Минздравсоцразвития России, г. Краснодар.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом