
Гладкие мышцы содержат актиновые и миозиновые нити, имеющие химические характеристики, подобные актиновым и миозиновым нитям скелетных мышц. Но в гладких мышцах нет тропонинового комплекса, необходимого для запуска сокращения скелетной мышцы, следовательно, механизм инициации сокращения в них другой. Этот механизм подробно обсуждается далее в нашей статье.
Химические исследования показали, что актиновые и миозиновые нити, извлеченные из гладких мышц, взаимодействуют друг с другом во многом так же, как и в скелетной мышце. Более того, процесс сокращения активируется ионами кальция, а энергия для сокращения обеспечивается разрушением АТФ до АДФ.
Существуют, однако, значительные различия в морфологической организации гладких и скелетных мышц, а также в сопряжении возбуждения и сокращения, механизме запуска ионами кальция сократительного процесса, длительности сокращения и количестве энергии, необходимой для сокращения.
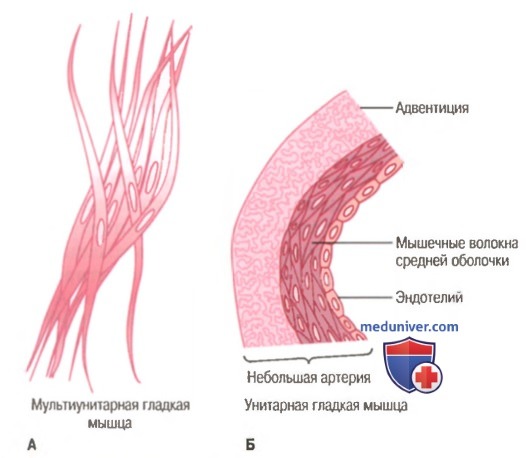
Мультиунитарная (А) и унитарная (Б) гладкие мышцы.
Морфологическая основа сокращения гладких мышц
Гладкие мышцы не имеют такой упорядоченной организации актиновых и миозиновых нитей, которая обнаруживается в скелетных мышцах, придавая им «полосатость». С помощью техники электронной микрофотографии выявляется гистологическая организация. Видно большое число актиновых нитей, прикрепленных к так называемым плотным тельцам. Некоторые из этих телец прикрепляются к клеточной мембране, другие распределяются внутри клетки. Некоторые из мембранных плотных телец соседних клеток связываются вместе мостиками из внутриклеточных белков. Через эти мостики в основном передается сила сокращения от одной клетки к другой.
В мышечном волокне среди актиновых нитей разбросаны миозиновые нити. Их диаметр более чем в 2 раза превышает диаметр актиновых нитей. На электронных микрофотографиях актиновых нитей обычно обнаруживают в 5-10 раз больше, чем миозиновых.
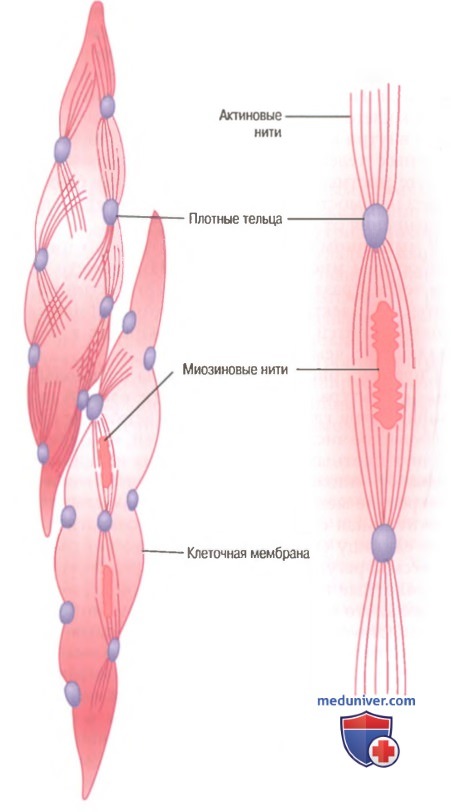
Морфологическая структура гладкой мышцы. В волокне слева вверху видны актиновые нити, исходящие из плотных телец. В волокнах слева внизу и справа на рисунке показано взаимоотношение между миозиновыми и актиновыми нитями.
На рисунке представлена предполагаемая структура отдельной сократительной единицы внутри гладкомышечной клетки, где видно большое число актиновых нитей, исходящих от двух плотных телец; концы этих нитей перекрывают миозиновую нить, расположенную посередине между плотными тельцами. Эта сократительная единица похожа на сократительную единицу скелетной мышцы, но без специфической регулярности ее структуры. В сущности, плотные тельца гладкой мышцы играют ту же роль, что и Z-диски в скелетной мышце.
Существует и другое различие. Большинство миозиновых нитей имеют поперечные мостики с так называемой боковой полярностью. Мостики организованы следующим образом: на одной стороне они шарнирно фиксируются в одном направлении, а на другой — в противоположном направлении. Это позволяет миозину тянуть актиновую нить с одной стороны в одном направлении, одновременно продвигая с другой стороны другую актиновую нить в противоположном направлении. Такая организация позволяет гладкомышечным клеткам сокращаться с укорочением до 80% их длины вместо укорочения менее чем на 30%, характерного для скелетной мышцы.
Большинство скелетных мышц сокращаются и расслабляются быстро, но сокращения гладких мышц в основном являются длительными тоническими сокращениями, которые иногда продолжаются в течение нескольких часов или даже дней. Следовательно, можно ожидать, что морфологические и химические особенности гладких мышц должны отличаться от соответствующих характеристик скелетных мышц. Далее обсуждаются некоторые из этих отличий.
Медленная циклическая активность миозиновых поперечных мостиков. В гладкой мышце по сравнению соскелетной гораздо меньше скорость циклической активности миозиновых поперечных мостиков, т.е. скорость их прикрепления к актину, отсоединение от актина и повторное прикрепление для осуществления следующего цикла. Фактически частота циклов составляет лишь от 1/10 до 1/300 этого показателя в скелетной мышце. Однако, как считают, в гладкой мышце значительно больше относительное количество времени, в течение которого поперечные мостики остаются прикрепленными к актиновым нитям, что является главным фактором, определяющим силу сокращения. Возможной причиной медленного циклирования является гораздо меньшая по сравнению со скелетной мышцей АТФ-азная активность головок поперечных мостиков, в связи с чем скорость разрушения АТФ — источника энергии для движения головок поперечных мостиков — значительно снижена с соответствующим замедлением скорости их циклов.
Видео физиология мышц и мышечного сокращения — профессор, д.м.н. П.Е. Умрюхин
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Гладкая мышечная ткань физиология
Это ткань энтомезенхимного происхождения, которая делится на два вида: висцеральную и сосудистую. В эмбриональном гистогенезе даже электронно-микроскопически трудно отличить мезенхимные предшественники фибробластов от гладких миоцитов. В малодифференцированных гладких миоцитах развиты гранулярная эндоплазматическая сеть, комплекс Гольджи. Тонкие филаменты ориентированы вдоль длинной оси клетки. По мере развития размеры клетки и число филаментов в цитоплазме возрастают. Постепенно объем цитоплазмы, занятый сократительными филаментами, увеличивается, расположение их становится все более упорядоченным. Пролиферативная активность гладких миоцитов в миогенезе постепенно снижается. Это происходит в результате увеличения продолжительности клеточного цикла, выхода клеток из цикла репродукции и перехода в дифференцированное состояние.
Однако и в дефинитивном состоянии в гладкой мышечной ткани клеточная регенерация в виде размножения миоцитов полностью не прекращается. Существуют данные о том, что пролиферация и дифференцировка в большей степени свойственна субпопуляции малых (по размерам) гладких миоцитов.
Строение гладкой мышечной ткани. Структура дефинитивных гладких миоцитов (лейомиоцитов), входящих в состав внутренних органов и стенки сосудов, имеет много общего, но в то же время характеризуется гетероморфией. Так, в стенках вен и артерий обнаруживаются овоидные, веретеновидные, отростчатые миоциты длиной 10-40 мкм, доходящие иногда до 140 мкм.
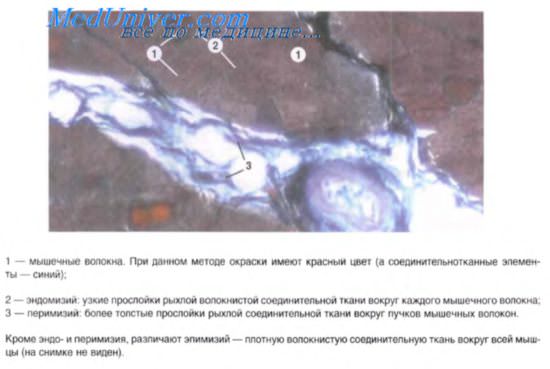
Гладкая мышечная ткань
Наибольшей длины гладкие миоциты достигают в стенке матки — до 500 мкм. Диаметр миоцитов колеблется от 2 до 20 мкм. В зависимости от характера внутриклеточных биосинтетических процессов различают контрактилъные и секреторные миоциты. Первые специализированы на функции сокращения, но вместе с тем сохраняют секреторную активность. Плазмолемма расслабленной клетки имеет ровную поверхность, а при сокращении становится складчатой. В центре клетки имеется палочковидное ядро, которое при сокращении клетки спиралевидно изгибается. Практически все ядра миоцитов содержат диплоидное количество ДНК. Гладкая эндоплазматическая сеть занимает примерно 2-7% объема цитоплазмы, а гранулярная сеть в контрактильных миоцитах выражена плохо. Митохондрии мелкие, сферические или овоидные, расположены у полюсов ядра. Характерной чертой гладких миоцитов является наличие множества впячиваний (кавеол) плазмолеммы, содержащих ионы кальция.
Секреторные миоциты (синтетические) по своей ультраструктуре напоминают фибробласты, однако содержат в цитоплазме пучки тонких миофиламентов, расположенные на периферии клетки. В цитоплазме хорошо развиты комплекс Гольджи, гранулярная эндоплазматическая сеть, много митохондрий, гранул гликогена, свободных рибосом и полисом. По степени зрелости такие клетки относят к малодифференцированным.
Читайте также: Что пьют спортсмены для восстановления хрящевой ткани суставов препараты
Сократительный аппарат миоцитов представлен тонкими актиновыми филамен-тами (гладкомышечным альфа-актином), связанными с тропомиозином. Толстые нити состоят из миозина, мономеры которого располагаются вблизи филаментов актина. Соотношение актиновых и миозиновых филаментов в гладком миоците составляет 12 к 1. Важным компонентом контрактильного аппарата миоцитов являются электронно-плотные структуры — тельца прикрепления, расположенные свободно в цитоплазме (плотные тельца) или тесно связанные с плазмолеммой. Основными белковыми компонентами плотных телец являются альфа-актинин, актин (немышечный) и кальпонин, что позволяет расссматривать их как функциональный эквивалент Z-линий миофибрилл скелетной мышцы. Актиновые филаменты фиксируются на плотных тельцах. Промежуточные филаменты, включающие десмин и виментин, обеспечивают связи между плотными тельцами и плазмолеммой, образуя прикрепительные пластины.
Сократительные белки формируют решетчатую структуру, закрепленную по окружности плазмолеммы, поэтому сокращение выражается в укорочении клетки, которая приобретает складчатую форму, тогда как в состоянии покоя клетка вытянута. При возникновении нервного импульса, распространяющегося по плазмолемме миоцита, происходит повышение уровня внутриклеточного Са2+, который поступает в цитоплазму из кавеол, отшнуровывающихся в цитоплазму в виде пузырьков. Высвобождение ионов кальция приводит к каскаду реакций, в результате которого происходит полимеризация миозина и образование перекрестных связей миозина вдоль актиновых филаментов по мере развития мышечного сокращения. Расслабление мышцы возникает при восстановлении концентрации исходного уровня Са2+ внутри клетки путем его перемещения внутрь саркоплазматической сети. При этом образовавшиеся в присутствии ионов кальция связи между актином и миозином нарушаются, акто-миозиновый комплекс распадается, гладкий миоцит расслабляется.
Гладкие миоциты синтезируют протеогликаны, гликопротеиды, проколлаген, проэластин, из которых формируются коллагеновые и эластические волокна и основное вещество межклеточного матрикса.
Взаимодействие миоцитов осуществляется с помощью цитоплазматических мостиков, взаимных впячиваний, нексусов, десмосом или простых участков мембранных контактов клеточных поверхностей.
Регенерация гладкой мышечной ткани
Гладкая мышечная ткань висцерального и сосудистого видов обладает значительной чувствительностью к воздействию экстремальных факторов.
В активированных миоцитах возрастает уровень биосинтетических процессов, морфологическим выражением которых являются синтез сократительных белков, укрупнение и гиперхроматоз ядра, гипертрофия ядрышка, возрастание показателей ядерно-цитоплазменного отношения, увеличение количества свободных рибосом и полисом, активация ферментов, аэробного и анаэробного фосфорилирования, мембранного транспорта. Клеточная регенерация осуществляется как за счет дифференцированных клеток, обладающих способностью вступать в митотический цикл, так и за счет активизации камбиальных элементов (миоцитов малого объема).
При действии ряда повреждающих факторов отмечается фенотипическая трансформация контрактильных миоцитов в секреторные. Данная трансформация часто наблюдается при повреждении интимы сосудов, формировании ее гиперплазии при развитии атеросклероза.
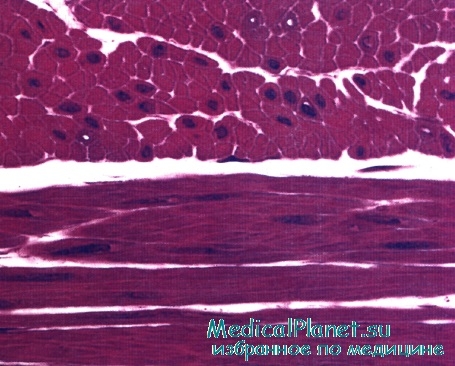
Гладкая мышечная ткань в поперечном (наверху) и продольном (внизу) разрезах. Обратите внимание на центрально расположенные ядра. Во многих клетках ядра не попали в срез.
Окраска: парарозанилин—толуидиновый синий. Среднее увеличение.
Нормальная физиология
По структуре гладкая мышца отличается от поперечнополосатой скелетной мышцы и мышцы сердца. Она состоит из клеток длиной от 10 до 500 мкм, шириной 5-10 мкм, содержащих одно ядро.
Гладкая мышца играет важную роль в регуляции просвета воздухоносных путей, кровеносных сосудов, двигательной активности желудочно-кишечного тракта, матки и др.
Гладкие мышцы органов существенно отличаются друг от друга. Основные отличия заключаются в том, что они имеют различные размеры, организованы в пучки или пласты, по разному отвечают на раздражители, имеют разную иннервацию и функцию. Все же для простоты гладкие мышцы подразделяют, главным образом, на два типа: мультиунитарные и унитарные.
Мультиунитарная гладкая мышца. Этот тип гладкой мышцы состоит из отдельных гладкомышечных клеток, каждая из которых, находится независимо друг от друга. Мультиунитарная гладкая мышца имеет большую плотность иннервации. Как и поперечно-полосатые мышечные волокна, они снаружи покрыты веществом, напоминающим базальную мембрану, в состав которого входят, изолирующие клетки друг от друга, коллагеновые и гликопротеиновые волокна.
Существенной особенностью мультиунитарной гладкой мышцы является то, что каждая мышечная клетка может сокращаться отдельно и ее активность регулируется нервными импульсами. Мультиунитарные мышцы входят в состав цилиарной мышцы, мышц радужки глаза, мышцы поднимающей волос.
Унитарная гладкая мышца (висцеральная). Данный термин является не совсем правильным, так как обозначает не одиночные мышечные волокна. В действительности это сотни миллионов гладкомышечных клеток, сокращающихся как единое целое. Обычно висцеральная мышца представляет собой пласт или пучок, а сарколеммы отдельных миоцитов имеют множественные точки соприкосновения. Это позволяет возбуждению распространяться от одной клетки к другой. Более того, мембраны рядом расположенных клеток образуют множественные плотные контакты ( gap junctions ), через которые ионы имеют возможность свободно передвигаться из одной клетки в другую. Таким образом, потенциал действия, возникающий на мембране гладкомышечной клетки, и ионные потоки могут распространяться по мышечному волокну, обеспечивая возможность одновременного сокращения большого количества отдельных клеток. Данный тип взаимодействия известен как функциональный синцитий. Подобный тип гладкой мышцы представлен в стенках большинства внутренних органов, включая кишечник, желчевыводящие протоки, мочеточник и большинство кровеносных сосудов.
В гладкой мышцы отсутствуют саркомеры. Толстые и тонкие миофиламеты распределены по всей саркоплазме гладкого миоцита и не имеют такой стройной организации, как в поперечно-полосатой скелетной мышце. При этом тонкие филаменты прикрепляются к плотным тельцам. Некоторые из этих телец расположены на внутренней поверхноти сарколеммы, но большинство из них находятся в саркоплазмме. Плотные тельца состоят из альфа-актинина – белка обнаруженного в структуре Z-мембраны поперечнополосатых мышечных волокон. Некоторые из плотных телец расположенных на внутренней поверхности мембраны соприкасаются с плотными тельцами прилегающей клетки. Тем самым сила, создаваемая одной клеткой может передаваться следующей.
Толстые миофиламенты гладкой мышцы содержат миозин, а тонкие– актин, тропомиозин, кальдесмон, кальпонин, лейкотонин А и С. Однако в составе тонких миофиламентов не обнаружен тропонин.
В гладкомышечных клетках практически отсутствуют Т-трубочки. Кроме того, гладкомышечные клетки значительно меньше поперечнополосатых мышечных волокон и поэтому не имеют развитой системы Т-трубочек, предназначенных для проведения возбуждения к расположенному в глубине сократительному аппарату. Вместо них наблюдаются небольшие углубления в сарколемме, которые получили название кавеолы. Благодаря им увеличивается площадь поверхности миоцита, а также может обеспечиваться взаимосвязь потенциалов, возникающих на мембране и саркоплазматическим ретикулумом.
Потенциал покоя. Величина разности потенциалов на мембране гладкомышечных клеток существенно различается в зависимоти как от типа гладкой мышцы так и от тех условий, в которых она находится. Обычно в состоянии покоя мембранный потенциал гладкомышечной клетки нестабилен и находится в пределах от –30 до –50 мВ, что на 30 мВ меньше, чем в скелетной мышце.
Потенциал действия унитарной мышцы. Потенциал действия в унитарной (висцеральной) гладкой мышце возникает так же как и в скелетной мышце. В висцеральных гладких мышцах потенциал действия различается по форме, амплитуде и продолжительности. Он бывает (1) в виде спайка или (2) потенциал действия, имеющий плато. Типичный spike-потенциал, характерен для гладкой и скелетной мышцы. Его продолжительность от 10 до 50 мсек. Данный потенциал возникает при нанесении на гладкую мышцу электрического, химического раздражения, а также растяжения. Кроме того, потенциала действия подобного типа может возникать спонтанно. Потенциал действия, имеющий плато, своим началом напоминает spike-потенциал. Однако сразу после быстрой деполяризации начинается быстрая реполяризации. Однако она задерживается вплоть до 1000 мсек. Так формируется плато потенциала действия. Во время плато гладкая мышца длительное время остается укороченной. Подобный тип возбуждения имеет место в гладкой мышце мочевого пузыря, матки и др.
Необходимо отметить, что в мембране гладкомышечной клетки обнаружено гораздо большее количество потенциалзависимых кальциевых каналов, чем в мембране поперечнополосатых мышечных волокон. Более того, ионы натрия играют малую роль в гененерации потенциала действия. Вместо них большое значение в генерации потенциала действия принадлежит потоку ионов кальция внутрь гладкомышечной клетки. Однако кальциевые каналы открываются значительно медленее, чем натриевые каналы, но остаются открытыми значительно дольше. На основании этого можно понять почему потенциал действия гладкой мышцы развивается в течение столь длительного времени. Другой важной задачей входящего во время потенциала действия кальция является их прямое влияние на сократительный аппарат клетки.
Некоторые гладкомышечные клетки обладают способностью к самовозбуждению, то есть способны генерировать потенциал действия без воздействия внешнего раздражителя. Это часто связано с периодическими колебаниями мембранного потенциала. Очень часто подобная активность наблюдается в гладкой мышце кишечника. Медленные волновые колебания мембранного потенциала не являются потенциалом действия. Одним из возможных механизмов, объясняющих появление этих волновых колебаний мембранного потенциала, является периодическая активация и затухание активности натрий-калиевого насоса. Разность потенциалов на мембране гладкомышечной клетки увеличивается во время активации Na/K насоса и уменьшается при ее снижении. Другой возможной причиной данного явления является ритмическое увеличение или снижение проводимости ионных каналов.
Физиологическое значение медленных колебаний мембранного потенциала состоит в том, что они могут инициировать появление потенциала действия. Это возникает в том момент когда во время медленной волны разность потенциалов на мембране клетки снижается до –35 мВ. При этом, как правило, успевает возникнуть несколько потенциалов действия. Следовательно, медленные волны можно назвать пейсмекерными волнами и, таким образом, становится понятным каким образом они обуславливают ритмические сокращения кишки.
Одним из важных раздражителей, инициирующих сокращение гладких мышц, является их растяжение. Достаточное растяжение гладкой мышцы обычно сопровождается появлением потенциалов действия. Таким образом, появлению потенциалов действия при растяжении гладкой мышцы способствует два фактора: (1) медленные волновые колебания мембранного потенциала, на которые наслаивается (2) деполяризация, вызываемая растяжением гладкой мышцы. Данное свойство гладкой мышцы позволяет ей автоматически сокращаться при растяжении. Например, во время переполнения тонкого кишечника возникает перистальттическая волна, которая и продвигает содержимое.
Деполяризация мультиунитарной гладкой мышцы. В обычных условиях мультиунитарная гладкая мышца сокращается в ответ на поступление нервного импульса. Чаще всего из нервного окончания высвобождается ацетилхолин, в некоторых мультиунитарных мышцах, норадреналин или другой медиатор. В любом случае медиатор приводит к деполяризации мембраны гладкой мышцы и к последующему ее сокращению. Потенциал действия при этом не возникает. Причина данного явления в том, что мультиунитарные гладкомышечные клетки слишком малы для того чтобы генерировать потенциал действия. (Когда потенциал действия возникает на мембране висцеральной (унитарной) гладкой мышцы, то от 30 до 40 гладкомышечных клеток должны деполяризоваться одновременно до того момента, когда потенциал действия будет способен самостоятельно растпространяться вдоль мембраны гладкой мышцы. В мультиунитарной гладкой мышце не возникает потенциал действия, а локальная деполяризация, вызываемая высвобождением медиатора, способна к электроническому распространению.
Особенности актомиозинованого взаимодействия. В гладкой мышце движение актомиозиновых мостиков является более медленным процессом, по сравнению с поперечнополосатой мышцей. Однако время, в течение которого головки миозиновых молекул остаются прикрепленными к актину оказывается более длительным. Причиной столь медленного движения актомиозиновых мостиков гладкомышечных клеток является более низкая АТФ-азная активность головок их миозиновых молекул. Поэтому распад молекул АТФ и высвобождение энергии, необходимой для обеспечения движения актомиозиновых мостиков происходит не так быстро как в поперечнополосатой мышечной ткани. Это можно понять, если представить, что одна молекула АТФ необходима для одного движения актомиозинового мостика независимо от продолжительности данного движения. Экономичность энерготрат в гладкой мышце является чрезвычайно важным в общем потреблении организмом энергии, так как, кровеносные сосуды, тонкий кишечник, мочевой пузырь, желчный пузырь и другие внутренние органы постоянно находятся в тонусе.
Особенность электромеханического сопряжения. Продолжительность сокращения гладких мышц может варьировать от 0,2 до 30 сек. Сокращение типичной гладкой мышцы начинается спустя от 50 до 100 мсек после начала ее возбуждения, достигая своего максимума через 0,5 сек, а затем угасает в течение последующих 1-2 сек. Таким образом, продолжительность сокращения составляет 1-3 сек, что является в 30 раз более длительным, чем в поперечнополосатой мышце.
Возникновение сокращения в гладкомышечных клетках в ответ на увеличение внутриклеточной концентрации ионов кальция – электромеханическое сопряжение осуществляется гораздо медленнее, чем в поперечнополосатой мышце.
Механизм электромеханического сопряжения в гладкой мышце отличается от поперечнополосатой или сердечной мышцы. В гладкой мышце появление на сарколемме потенциала действия активирует фосфолипазу С и появление инозитол-3-фосфата, который связывается со специфическим для него рецептором, расположенным на кальциевом канале терминальной цистерны СПР. Это приводит к открытию этих каналов и выходу кальция из цистерны СПР.
Особенность силы сокращения и укорочения гладкой мышцы . Сила сокращения гладкой мышцы равна от 4 до 6 кг/см2 поперечного сечения гладкой мышцы. В тоже время поперечнополосатая мышца развивает силу от 3 до 4 кг/см2. Данный факт является следствием значительного времени взаимодействия актиновых и миозиновых филаментов.
Ещё одной особенностью гладкой мышцы является то, что она во время сокращения способна укорачиваться вплоть до 2/3 ее первоначальной длины (скелетная мышца от 1/4 до 1/3 длины). Это позволяет полым органам выполнять свою функцию — изменять свой просвет от в значительных пределах. Точный механизм данного явления не известен. Но подобное возможно благодаря двум причинам:
• в гладкой мышце существует оптимальная площадь соприкосновения актиновых и миозиновых нитей;
• длина актиновых нитей в гладкой мышце гораздо больше, чем в поперечнополосатой. Поэтому взаимодействие актиновых и миозиновых нитей может происходить в них на гораздо более продолжительном расстоянии, чем это имеет место при сокращении поперечнополосатой мышцы.
Стресс-релаксация гладкой мышцы. Еще одной важной особенностью висцеральной гладкой мышцы многих полых органов является ее способность возвращаться к первоначальной силе сокращения спустя секунды или минуты после того, как она была растянута или сокращена. Например, внезапное увеличение объёма жидкости в полости мочевого пузыря сопровождается растяжением гладкой мышцы его стенки, что обязательно приводит к увеличению внутрипузырного давления. Однако в последующие от 15 сек до нескольких минут, несмотря на постоянно действующую растягивающую силу внутрипузырное давление возвращается к почти исходному значению.
Читайте также: Белизна средство для отбеливания тканей
Несмотря на то, что скелетные мышечные волокна сокращаются быстро, для большинства гладких мышц характерно длительное тоническое сокращение, иногда достигающее часов или даже дней. Можно предположить, что механизм сокращения гладкой мышцы отличается от поперечнополосатой мышцы. Подобно скелетной мышце для начала мышечного сокращения гладкомышечных волокон необходимы ионы кальция. Увеличение внутриклеточной концентрации ионов кальция может происходить при раздражении нервных волокон, под влиянием гормонов, растяжения гладкой мышцы или даже в результате изменения химического окружения мышечного волокна.
В гладкой мышце движение поперечных актомиозиновых мостиков, лежащее в основе сокращения, начинается благодаря кальций-зависимому процессу фосфорилирования головок миозиновых молекул.
Миозиновые молекулы содержат 4 легкие цепи, две из которых связаны с головкой молекулы миозина. Головка миозиновой молекулы присоединяется к актину только после того, как на ней фосфорилируется одна из легких цепей, получившая название регуляторной. Фосфорилирование легкой цепи миозина катализируется киназой легких цепей миозина (КЛЦМ), которая активируется кальмодулином после его взаимодействия с ионами кальция.
Дефосфорилирование легких цепей миозина осуществляется фосфатазой легких цепей миозина (ФЛЦМ). Скорость укорочения гладкого миоцита (то есть скорость циклинга акто-миозиновых мостиков) зависит от интенсивности фосфорилирования легких цепей миозина. При преобладании процесса дефосфорилирования над процессом фосфорилирования гладкая мышца расслабляется.
Ионы кальция могут поступать в клетку несколькими путями.
• Под влиянием медиаторов. При взаимодействии медиатора с расположенным на поверхности гладкомышечной клетки соответствующим рецептором, происходит открытие рецептор-активируемого Са ++ канала и вход ионов кальция внутрь клетки.
• Через потенциал-зависимые каналы, открывающиеся при изменении разности потенциалов на мембране гладкомышечной клетки. Ионы кальция могут поступать в клетку через потенциал-зависимые кальциевые каналы, которые открывается в мембране гладкомышечных клеток при появлении на ней потенциала действия.
• Источником ионов кальция может быть саркоплазматический ретикулум. В мембране саркоплазматического ретикулума есть каналы, которые активируются (открываются) инозитолтрифосфатом (IP3) и поэтому получили название IP3-рецепторов. Это название позволяет отличать их от рианодиновых рецепторов, обнаруженных в саркоплазматическом ретикулуме поперечнополосатых мышц.
Механизм длительно удерживаемого укорочения ( «latch» -механизм). «Мостики на замке» . Дефосфорилированые поперечные мостики, но оставшиеся прикрепленными к актину получили название мостики на замке. Это позволяет гладкой мышце поддерживать тонус при минимальных энергетических затратах и связано с тем, что данные мостики не циклируют и поэтому не требуют большого количества энергии АТФ. Подобное явление в значительно меньшей степени имеет место и в поперечнополосатой скелетной мышце, и также не требует большого числа нервных импульсов и концентрации гормонов.
Читайте также: Ткань оксфорд 240 pu 3000
Влияние тканевых метаболитов на сократительную активность гладкой мышцы. Сократительная активность гладкомышечной ткани, например, сосудистой, в значительной степени зависит от химического состава окружающей межклеточной жидкости. Таким образом, осуществляется регуляция сосудистого тонуса по принципу отрицательной обратной связи. Уменьшение напряжения кислорода, увеличение напряжения углекислого газа или ионов водорода в тканевой жидкости сопровождается вазодилатацией. Кроме того, увеличение концентрации аденозина, молочной кислоты, ионов калия или уменьшения ионов кальция, а также уменьшение температуры тела приводит к локальной вазодилатации.
Влияние гормонов на сократительную активность гладкой мышцы. Среди гормонов, циркулирующих в крови, обладающих выраженным эффектом на активность гладкой мышцы можно выделить следующие: адреналин, норадреналин, вазопресин, ангиотензин, окситоцин, а также такие биоактивные вещества как ацетилхолин, серотонин и гистамин. В гладкой мышце под влиянием гормона приходит активация сокращения только в том случае если на поверхности ее мембраны находится соответствующий рецептор, связанный с каналом, имеющим лиганд-активируемое воротное устройство. Напротив, гормон вызывает торможение активности гладких миоцитов если взаимодействует с ингибиторным рецептором.
Механизм сокращения и расслабления гладкой мышцы, вызванного гормонами и тканевыми метаболитами. Если гормоно-рецепторное взаимодействие приводит к открытию натриевых или кальциевых каналов, то развивается деполяризация их мембраны таким же образом как это происходит при воздействии нервного импульса. В некоторых случаях развивается потенциал действия. Однако очень часто деполяризация наблюдается без потенциала действия. Как правило, эта деполяризация обусловлена входом внутрь клетки ионов кальция, которые инициирует сокращение гладкой мышцы.
В том случае, если гормон-рецепторное взаимодействие ингибирует сокращение, то, как правило, это связано с закрытием натриевых или кальциевых каналов, что не позволяет положительным ионам входить в клетку или приводит к открытию калиевых каналов, через которые положительно заряженные ионы калия, выходят из клетки. В любом случае увеличивается элетроотрицательность внутренней поверхности мембраны и развивается ее гиперполяризация. Кроме того, существует возможность активации сократительной активности гладкой мышцы без изменения мембранного потенциала. В этом случае под влиянием гормон-рецепторного взаимодействия не происходит открытие каких-либо каналов, расположенных в сарколемме, но вместо этого кальций высвобождается из саркоплазматического ретикулума и инициирует сокращение мышцы. В другом случае гормоно-рецепторное взаимодействие приводит к активации аденилат или гуанилатциклазы, расположенной на внутренней поверхности сарколеммы. При этом происходит увеличение внутриклеточной концентрации вторичных посредников, таких как ц-АМФ или ц-ГМФ. В свою очередь, ц-АМФ и ц-ГМФ обладают множеством разнообразных эффектов, один из которых заключается в том, что под их влиянием происходит фосфорилирование протеинкиназ, а затем и ферментов, участвующих в угнетении сократительной активности гладкой мышцы. Данному эффекту способствует и то, что данные вещества активируют кальциевый насос, откачивающий ионы кальция из саркоплазмы в саркоплазматический ретикулум.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом
- Правообладателям
- Политика конфиденциальности