
Гладкие мышцы содержат актиновые и миозиновые нити, имеющие химические характеристики, подобные актиновым и миозиновым нитям скелетных мышц. Но в гладких мышцах нет тропонинового комплекса, необходимого для запуска сокращения скелетной мышцы, следовательно, механизм инициации сокращения в них другой. Этот механизм подробно обсуждается далее в нашей статье.
Химические исследования показали, что актиновые и миозиновые нити, извлеченные из гладких мышц, взаимодействуют друг с другом во многом так же, как и в скелетной мышце. Более того, процесс сокращения активируется ионами кальция, а энергия для сокращения обеспечивается разрушением АТФ до АДФ.
Существуют, однако, значительные различия в морфологической организации гладких и скелетных мышц, а также в сопряжении возбуждения и сокращения, механизме запуска ионами кальция сократительного процесса, длительности сокращения и количестве энергии, необходимой для сокращения.
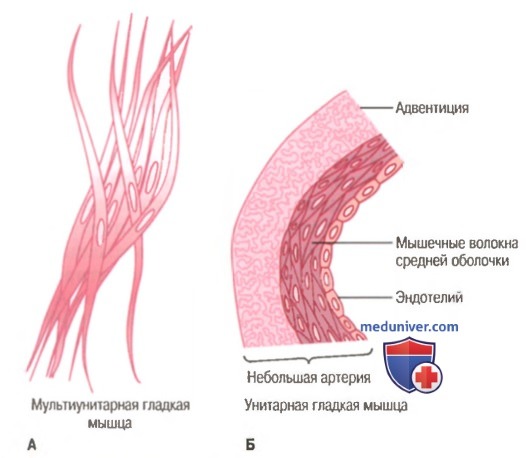
Мультиунитарная (А) и унитарная (Б) гладкие мышцы.
Морфологическая основа сокращения гладких мышц
Гладкие мышцы не имеют такой упорядоченной организации актиновых и миозиновых нитей, которая обнаруживается в скелетных мышцах, придавая им «полосатость». С помощью техники электронной микрофотографии выявляется гистологическая организация. Видно большое число актиновых нитей, прикрепленных к так называемым плотным тельцам. Некоторые из этих телец прикрепляются к клеточной мембране, другие распределяются внутри клетки. Некоторые из мембранных плотных телец соседних клеток связываются вместе мостиками из внутриклеточных белков. Через эти мостики в основном передается сила сокращения от одной клетки к другой.
В мышечном волокне среди актиновых нитей разбросаны миозиновые нити. Их диаметр более чем в 2 раза превышает диаметр актиновых нитей. На электронных микрофотографиях актиновых нитей обычно обнаруживают в 5-10 раз больше, чем миозиновых.
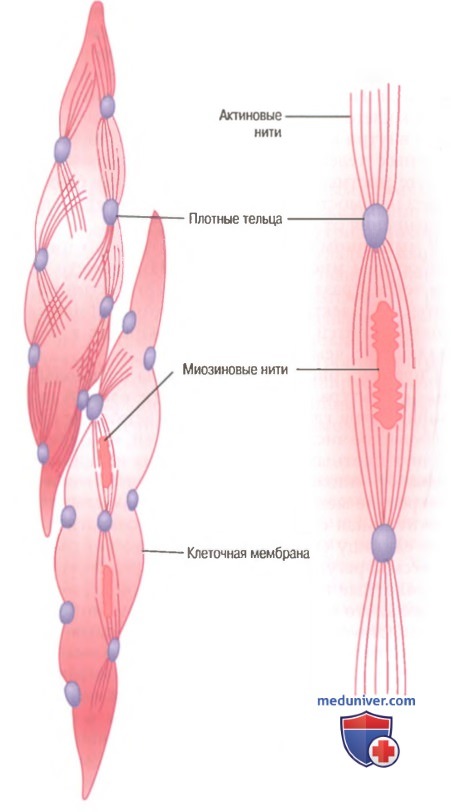
Морфологическая структура гладкой мышцы. В волокне слева вверху видны актиновые нити, исходящие из плотных телец. В волокнах слева внизу и справа на рисунке показано взаимоотношение между миозиновыми и актиновыми нитями.
На рисунке представлена предполагаемая структура отдельной сократительной единицы внутри гладкомышечной клетки, где видно большое число актиновых нитей, исходящих от двух плотных телец; концы этих нитей перекрывают миозиновую нить, расположенную посередине между плотными тельцами. Эта сократительная единица похожа на сократительную единицу скелетной мышцы, но без специфической регулярности ее структуры. В сущности, плотные тельца гладкой мышцы играют ту же роль, что и Z-диски в скелетной мышце.
Существует и другое различие. Большинство миозиновых нитей имеют поперечные мостики с так называемой боковой полярностью. Мостики организованы следующим образом: на одной стороне они шарнирно фиксируются в одном направлении, а на другой — в противоположном направлении. Это позволяет миозину тянуть актиновую нить с одной стороны в одном направлении, одновременно продвигая с другой стороны другую актиновую нить в противоположном направлении. Такая организация позволяет гладкомышечным клеткам сокращаться с укорочением до 80% их длины вместо укорочения менее чем на 30%, характерного для скелетной мышцы.
Большинство скелетных мышц сокращаются и расслабляются быстро, но сокращения гладких мышц в основном являются длительными тоническими сокращениями, которые иногда продолжаются в течение нескольких часов или даже дней. Следовательно, можно ожидать, что морфологические и химические особенности гладких мышц должны отличаться от соответствующих характеристик скелетных мышц. Далее обсуждаются некоторые из этих отличий.
Медленная циклическая активность миозиновых поперечных мостиков. В гладкой мышце по сравнению соскелетной гораздо меньше скорость циклической активности миозиновых поперечных мостиков, т.е. скорость их прикрепления к актину, отсоединение от актина и повторное прикрепление для осуществления следующего цикла. Фактически частота циклов составляет лишь от 1/10 до 1/300 этого показателя в скелетной мышце. Однако, как считают, в гладкой мышце значительно больше относительное количество времени, в течение которого поперечные мостики остаются прикрепленными к актиновым нитям, что является главным фактором, определяющим силу сокращения. Возможной причиной медленного циклирования является гораздо меньшая по сравнению со скелетной мышцей АТФ-азная активность головок поперечных мостиков, в связи с чем скорость разрушения АТФ — источника энергии для движения головок поперечных мостиков — значительно снижена с соответствующим замедлением скорости их циклов.
Читайте также: Как сделать чтобы колпак из ткани стоял
Видео физиология мышц и мышечного сокращения — профессор, д.м.н. П.Е. Умрюхин
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Гладкая мышечная ткань гормоны
Это ткань энтомезенхимного происхождения, которая делится на два вида: висцеральную и сосудистую. В эмбриональном гистогенезе даже электронно-микроскопически трудно отличить мезенхимные предшественники фибробластов от гладких миоцитов. В малодифференцированных гладких миоцитах развиты гранулярная эндоплазматическая сеть, комплекс Гольджи. Тонкие филаменты ориентированы вдоль длинной оси клетки. По мере развития размеры клетки и число филаментов в цитоплазме возрастают. Постепенно объем цитоплазмы, занятый сократительными филаментами, увеличивается, расположение их становится все более упорядоченным. Пролиферативная активность гладких миоцитов в миогенезе постепенно снижается. Это происходит в результате увеличения продолжительности клеточного цикла, выхода клеток из цикла репродукции и перехода в дифференцированное состояние.
Однако и в дефинитивном состоянии в гладкой мышечной ткани клеточная регенерация в виде размножения миоцитов полностью не прекращается. Существуют данные о том, что пролиферация и дифференцировка в большей степени свойственна субпопуляции малых (по размерам) гладких миоцитов.
Строение гладкой мышечной ткани. Структура дефинитивных гладких миоцитов (лейомиоцитов), входящих в состав внутренних органов и стенки сосудов, имеет много общего, но в то же время характеризуется гетероморфией. Так, в стенках вен и артерий обнаруживаются овоидные, веретеновидные, отростчатые миоциты длиной 10-40 мкм, доходящие иногда до 140 мкм.
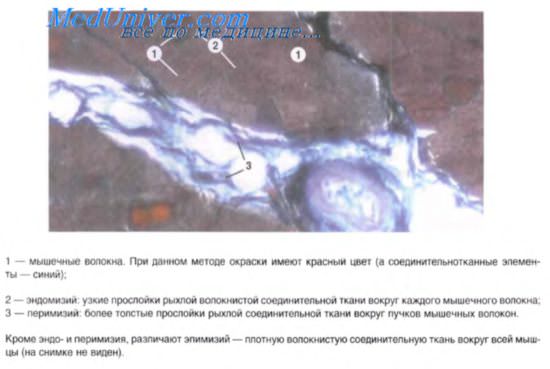
Гладкая мышечная ткань
Наибольшей длины гладкие миоциты достигают в стенке матки — до 500 мкм. Диаметр миоцитов колеблется от 2 до 20 мкм. В зависимости от характера внутриклеточных биосинтетических процессов различают контрактилъные и секреторные миоциты. Первые специализированы на функции сокращения, но вместе с тем сохраняют секреторную активность. Плазмолемма расслабленной клетки имеет ровную поверхность, а при сокращении становится складчатой. В центре клетки имеется палочковидное ядро, которое при сокращении клетки спиралевидно изгибается. Практически все ядра миоцитов содержат диплоидное количество ДНК. Гладкая эндоплазматическая сеть занимает примерно 2-7% объема цитоплазмы, а гранулярная сеть в контрактильных миоцитах выражена плохо. Митохондрии мелкие, сферические или овоидные, расположены у полюсов ядра. Характерной чертой гладких миоцитов является наличие множества впячиваний (кавеол) плазмолеммы, содержащих ионы кальция.
Секреторные миоциты (синтетические) по своей ультраструктуре напоминают фибробласты, однако содержат в цитоплазме пучки тонких миофиламентов, расположенные на периферии клетки. В цитоплазме хорошо развиты комплекс Гольджи, гранулярная эндоплазматическая сеть, много митохондрий, гранул гликогена, свободных рибосом и полисом. По степени зрелости такие клетки относят к малодифференцированным.
Сократительный аппарат миоцитов представлен тонкими актиновыми филамен-тами (гладкомышечным альфа-актином), связанными с тропомиозином. Толстые нити состоят из миозина, мономеры которого располагаются вблизи филаментов актина. Соотношение актиновых и миозиновых филаментов в гладком миоците составляет 12 к 1. Важным компонентом контрактильного аппарата миоцитов являются электронно-плотные структуры — тельца прикрепления, расположенные свободно в цитоплазме (плотные тельца) или тесно связанные с плазмолеммой. Основными белковыми компонентами плотных телец являются альфа-актинин, актин (немышечный) и кальпонин, что позволяет расссматривать их как функциональный эквивалент Z-линий миофибрилл скелетной мышцы. Актиновые филаменты фиксируются на плотных тельцах. Промежуточные филаменты, включающие десмин и виментин, обеспечивают связи между плотными тельцами и плазмолеммой, образуя прикрепительные пластины.
Сократительные белки формируют решетчатую структуру, закрепленную по окружности плазмолеммы, поэтому сокращение выражается в укорочении клетки, которая приобретает складчатую форму, тогда как в состоянии покоя клетка вытянута. При возникновении нервного импульса, распространяющегося по плазмолемме миоцита, происходит повышение уровня внутриклеточного Са2+, который поступает в цитоплазму из кавеол, отшнуровывающихся в цитоплазму в виде пузырьков. Высвобождение ионов кальция приводит к каскаду реакций, в результате которого происходит полимеризация миозина и образование перекрестных связей миозина вдоль актиновых филаментов по мере развития мышечного сокращения. Расслабление мышцы возникает при восстановлении концентрации исходного уровня Са2+ внутри клетки путем его перемещения внутрь саркоплазматической сети. При этом образовавшиеся в присутствии ионов кальция связи между актином и миозином нарушаются, акто-миозиновый комплекс распадается, гладкий миоцит расслабляется.
Читайте также: Эталон ткани для спецодежды
Гладкие миоциты синтезируют протеогликаны, гликопротеиды, проколлаген, проэластин, из которых формируются коллагеновые и эластические волокна и основное вещество межклеточного матрикса.
Взаимодействие миоцитов осуществляется с помощью цитоплазматических мостиков, взаимных впячиваний, нексусов, десмосом или простых участков мембранных контактов клеточных поверхностей.
Регенерация гладкой мышечной ткани
Гладкая мышечная ткань висцерального и сосудистого видов обладает значительной чувствительностью к воздействию экстремальных факторов.
В активированных миоцитах возрастает уровень биосинтетических процессов, морфологическим выражением которых являются синтез сократительных белков, укрупнение и гиперхроматоз ядра, гипертрофия ядрышка, возрастание показателей ядерно-цитоплазменного отношения, увеличение количества свободных рибосом и полисом, активация ферментов, аэробного и анаэробного фосфорилирования, мембранного транспорта. Клеточная регенерация осуществляется как за счет дифференцированных клеток, обладающих способностью вступать в митотический цикл, так и за счет активизации камбиальных элементов (миоцитов малого объема).
При действии ряда повреждающих факторов отмечается фенотипическая трансформация контрактильных миоцитов в секреторные. Данная трансформация часто наблюдается при повреждении интимы сосудов, формировании ее гиперплазии при развитии атеросклероза.
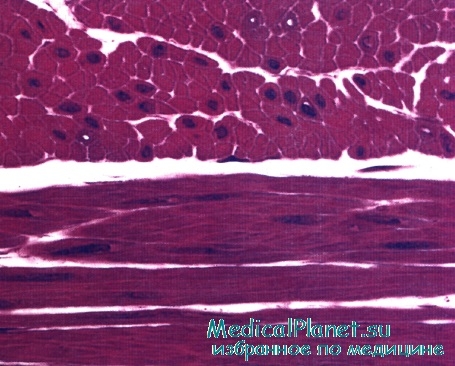
Гладкая мышечная ткань в поперечном (наверху) и продольном (внизу) разрезах. Обратите внимание на центрально расположенные ядра. Во многих клетках ядра не попали в срез.
Окраска: парарозанилин—толуидиновый синий. Среднее увеличение.
Гладкая мышечная ткань гормоны
Возбуждение висцеральных гладких мышц при их растяжении. При достаточной степени растяжения висцеральной (унитарной) гладкой мышцы обычно генерируются спонтанные потенциалы действия. Они возникают в результате комбинации: (1) нормальных медленноволновых потенциалов; (2) снижения общей отрицательности мембранного потенциала, связанного с самим растяжением. Это позволяет кишечной стенке при чрезмерном растяжении автоматически и ритмически сокращаться. Например, когда кишечник переполняется, местные автоматические сокращения часто вызывают перистальтические волны, которые перемещают содержимое из переполненной кишки обычно в направлении ануса.
Гладкомышечные волокна мультиунитарных гладких мышц (например, мышца радужной оболочки глаза или мышца, поднимающая волос) в норме сокращаются в основном в ответ на нервные стимулы. В одних мультиунитарных гладких мышцах нервные окончания секретируют ацетилхолин, в других — норадреналин. В обоих случаях медиатор вызывает деполяризацию мембраны гладкомышечных волокон, а это, в свою очередь, ведет к их сокращению. Потенциалы действия обычно не развиваются в связи с тем, что эти волокна слишком малы для генерации потенциала действия. (Для возникновения самораспространяющегося потенциала действия в висцеральной (унитарной) гладкой мышце должны деполяризоваться одновременно 30-40 гладкомышечных волокон.) Однако в маленьких гладкомышечных клетках даже без потенциала действия локальная деполяризация (называемая также синаптическим потенциалом), возникающая под действием нейромедиатора, сама распространяется электротонически по всему волокну, что вполне достаточно для мышечного сокращения.
Вероятно, половина всех гладкомышечных сокращений инициируется стимулирующими факторами, действующими непосредственно на сократительный аппарат гладкой мышцы без потенциалов действия. Часто вовлекаются два типа стимулирующих факторов, действие которых не связано с нервными влияниями или развитием потенциалов действия, а именно:
(1) местные тканевые химические факторы;
(2) различные гормоны.
Реакция гладких мышц на действие местных тканевых химических факторов. В нашей статье обсуждается сокращение артериол, метартериол и прекапиллярных сфинктеров. Мельчайшие из этих сосудов не имеют или почти не имеют нервного снабжения. Однако гладкая мышца обладает высокой сократимостью, быстро реагируя на изменения местных химических условий в окружающей интерстициальной жидкости.
Читайте также: Что такое деградация костной ткани
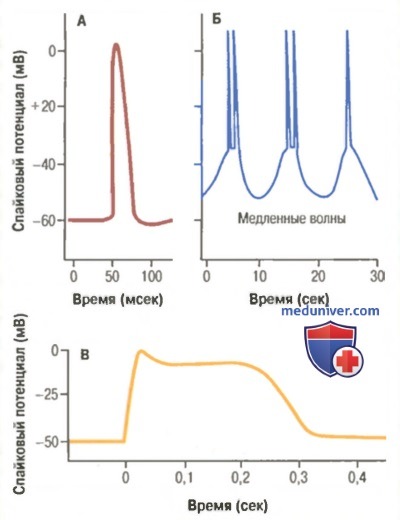
A. Типичный потенциал действия гладкой мышцы (спайковый потенциал), вызванный внешним стимулом.
Б. Повторные спайковые потенциалы, возбуждаемые медленными ритмичными волновыми изменениями мембранного потенциала, которые возникают спонтанно в гладкой мышце кишечной стенки.
B. Потенциал действия с плато, зарегистрированный от гладкомышечного волокна матки.
В норме в состоянии покоя многие из этих мелких кровеносных сосудов остаются суженными. Но когда необходим дополнительный приток крови к ткани, множество факторов могут расслабить сосудистую стенку, способствуя увеличению кровотока. Так мощная местная контролирующая система обратной связи регулирует кровоток через локальную тканевую область. Далее перечислен ряд специфических регулирующих факторов, вызывающих расширение сосудов (вазодилатацию).
1. Недостаток кислорода в тканях, вызывающий расслабление гладкой мышцы.
2. Избыток углекислого газа.
3. Увеличение концентрации ионов водорода.
Аденозин, молочная кислота, увеличение количества ионов калия, снижение концентрации ионов кальция и увеличение температуры тела могут вызвать местное расширение сосудов.
Влияние гормонов на сокращение гладких мышц. Большинство циркулирующих в крови гормонов влияют на сократительную активность гладких мышц в определенной степени, а некоторые из них оказывают мощное влияние. К наиболее важным относят норадреналин, адреналин, ацетилхолин, ангиотензин, эндотелии, вазопрессин, окситоцин, серотонин и гистамин.
Гормон вызывает сокращение гладкой мышцы, если мембрана мышечной клетки содержит возбуждающие рецепторы, управляемые соответствующим гормоном. И наоборот, гормон вызывает торможение, если мембрана содержит тормозные рецепторы для данного гормона.
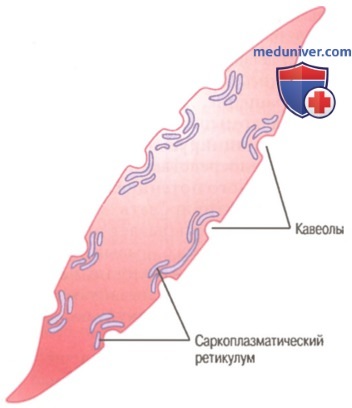
Трубочки саркоплазматического ретикулума в крупном гладкомышечном волокне. Показана связь трубочек с инвагинациями клеточной мембраны, которые называют кавеолами.
Механизмы возбуждения или торможения гладких мышц под влиянием гормонов или местных тканевых факторов. Некоторые гормональные рецепторы в мембране гладкомышечных волокон открывают натриевые или кальциевые ионные каналы и деполяризуют мембрану так же, как после нервной стимуляции. Иногда в результате развивается потенциал действия или, если потенциалы действия уже возникли, может усилиться импульсная активность. В других случаях возникающая деполяризация без развития потенциалов действия позволяет ионам кальция войти в клетку, способствуя ее сокращению.
Напротив, торможение возникает, когда гормон (или другой тканевой фактор) закрывает натриевые или кальциевые канальи предупреждая вход в клетку ионов Na+ и Са . Торможение происходит также, если открываются в норме закрытые калиевые каналы, что позволяет положительным калиевым ионам диффундировать из клетки. Оба эти действия увеличивают степень отрицательности внутри мышечной клетки, вызывая состояние, называемое гиперполяризацией, которое мощно тормозит мышечное сокращение.
Иногда гладкомышечное сокращение или его торможение инициируются гормонами, не вызывающими непосредственно каких-либо изменений мембранного потенциала. В этих случаях гормон может активировать мембранный рецептор, который не открывает никаких ионных каналов, а вместо этого вызывает внутреннее изменение в мышечном волокне, например выделение ионов кальция из внутриклеточного саркоплазматического ретикулума; затем кальций вызывает сокращение.
Для торможения сокращения известны другие рецепторные механизмы, которые активируют ферменты аденилатциклазу или гуанилатциклазу в клеточной мембране. Погруженные внутрь клетки части рецепторов сопряжены с этими ферментами, и их активация вызывает формирование циклического аденозинмонофосфата (цАМФ) или циклического гуанозинмонофосфата (цГМФ) — так называемых вторичных посредников. цАМФ и цГМФ имеют много эффектов, одним из которых является изменение степени фосфорилирования некоторых ферментов, опосредованно тормозящих сокращение. Активируется насос, удаляющий ионы кальция из саркоплазмы в саркоплазматический ретикулум, а также насос клеточной мембраны, удаляющий кальций из самой клетки; это снижает концентрацию ионов кальция в саркоплазме, тормозя сокращение.
Ответная реакция гладких мышц на действие различных гормонов, нейромедиаторов или других веществ отличается большим разнообразием. В некоторых случаях одно и то же вещество может вызывать либо сокращение, либо расслабление в гладких мышцах разной локализации. Например, норадреналин тормозит сокращение гладкой мышцы кишечника, но стимулирует сокращение гладких мышц кровеносных сосудов.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом