
Это ткань энтомезенхимного происхождения, которая делится на два вида: висцеральную и сосудистую. В эмбриональном гистогенезе даже электронно-микроскопически трудно отличить мезенхимные предшественники фибробластов от гладких миоцитов. В малодифференцированных гладких миоцитах развиты гранулярная эндоплазматическая сеть, комплекс Гольджи. Тонкие филаменты ориентированы вдоль длинной оси клетки. По мере развития размеры клетки и число филаментов в цитоплазме возрастают. Постепенно объем цитоплазмы, занятый сократительными филаментами, увеличивается, расположение их становится все более упорядоченным. Пролиферативная активность гладких миоцитов в миогенезе постепенно снижается. Это происходит в результате увеличения продолжительности клеточного цикла, выхода клеток из цикла репродукции и перехода в дифференцированное состояние.
Однако и в дефинитивном состоянии в гладкой мышечной ткани клеточная регенерация в виде размножения миоцитов полностью не прекращается. Существуют данные о том, что пролиферация и дифференцировка в большей степени свойственна субпопуляции малых (по размерам) гладких миоцитов.
Строение гладкой мышечной ткани. Структура дефинитивных гладких миоцитов (лейомиоцитов), входящих в состав внутренних органов и стенки сосудов, имеет много общего, но в то же время характеризуется гетероморфией. Так, в стенках вен и артерий обнаруживаются овоидные, веретеновидные, отростчатые миоциты длиной 10-40 мкм, доходящие иногда до 140 мкм.
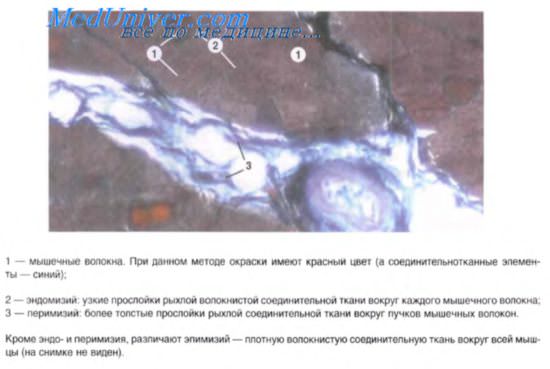
Гладкая мышечная ткань
Наибольшей длины гладкие миоциты достигают в стенке матки — до 500 мкм. Диаметр миоцитов колеблется от 2 до 20 мкм. В зависимости от характера внутриклеточных биосинтетических процессов различают контрактилъные и секреторные миоциты. Первые специализированы на функции сокращения, но вместе с тем сохраняют секреторную активность. Плазмолемма расслабленной клетки имеет ровную поверхность, а при сокращении становится складчатой. В центре клетки имеется палочковидное ядро, которое при сокращении клетки спиралевидно изгибается. Практически все ядра миоцитов содержат диплоидное количество ДНК. Гладкая эндоплазматическая сеть занимает примерно 2-7% объема цитоплазмы, а гранулярная сеть в контрактильных миоцитах выражена плохо. Митохондрии мелкие, сферические или овоидные, расположены у полюсов ядра. Характерной чертой гладких миоцитов является наличие множества впячиваний (кавеол) плазмолеммы, содержащих ионы кальция.
Секреторные миоциты (синтетические) по своей ультраструктуре напоминают фибробласты, однако содержат в цитоплазме пучки тонких миофиламентов, расположенные на периферии клетки. В цитоплазме хорошо развиты комплекс Гольджи, гранулярная эндоплазматическая сеть, много митохондрий, гранул гликогена, свободных рибосом и полисом. По степени зрелости такие клетки относят к малодифференцированным.
Сократительный аппарат миоцитов представлен тонкими актиновыми филамен-тами (гладкомышечным альфа-актином), связанными с тропомиозином. Толстые нити состоят из миозина, мономеры которого располагаются вблизи филаментов актина. Соотношение актиновых и миозиновых филаментов в гладком миоците составляет 12 к 1. Важным компонентом контрактильного аппарата миоцитов являются электронно-плотные структуры — тельца прикрепления, расположенные свободно в цитоплазме (плотные тельца) или тесно связанные с плазмолеммой. Основными белковыми компонентами плотных телец являются альфа-актинин, актин (немышечный) и кальпонин, что позволяет расссматривать их как функциональный эквивалент Z-линий миофибрилл скелетной мышцы. Актиновые филаменты фиксируются на плотных тельцах. Промежуточные филаменты, включающие десмин и виментин, обеспечивают связи между плотными тельцами и плазмолеммой, образуя прикрепительные пластины.
Сократительные белки формируют решетчатую структуру, закрепленную по окружности плазмолеммы, поэтому сокращение выражается в укорочении клетки, которая приобретает складчатую форму, тогда как в состоянии покоя клетка вытянута. При возникновении нервного импульса, распространяющегося по плазмолемме миоцита, происходит повышение уровня внутриклеточного Са2+, который поступает в цитоплазму из кавеол, отшнуровывающихся в цитоплазму в виде пузырьков. Высвобождение ионов кальция приводит к каскаду реакций, в результате которого происходит полимеризация миозина и образование перекрестных связей миозина вдоль актиновых филаментов по мере развития мышечного сокращения. Расслабление мышцы возникает при восстановлении концентрации исходного уровня Са2+ внутри клетки путем его перемещения внутрь саркоплазматической сети. При этом образовавшиеся в присутствии ионов кальция связи между актином и миозином нарушаются, акто-миозиновый комплекс распадается, гладкий миоцит расслабляется.
Гладкие миоциты синтезируют протеогликаны, гликопротеиды, проколлаген, проэластин, из которых формируются коллагеновые и эластические волокна и основное вещество межклеточного матрикса.
Взаимодействие миоцитов осуществляется с помощью цитоплазматических мостиков, взаимных впячиваний, нексусов, десмосом или простых участков мембранных контактов клеточных поверхностей.
Регенерация гладкой мышечной ткани
Гладкая мышечная ткань висцерального и сосудистого видов обладает значительной чувствительностью к воздействию экстремальных факторов.
В активированных миоцитах возрастает уровень биосинтетических процессов, морфологическим выражением которых являются синтез сократительных белков, укрупнение и гиперхроматоз ядра, гипертрофия ядрышка, возрастание показателей ядерно-цитоплазменного отношения, увеличение количества свободных рибосом и полисом, активация ферментов, аэробного и анаэробного фосфорилирования, мембранного транспорта. Клеточная регенерация осуществляется как за счет дифференцированных клеток, обладающих способностью вступать в митотический цикл, так и за счет активизации камбиальных элементов (миоцитов малого объема).
Читайте также: Балкон в многоквартирном доме: является ли он общедомовым имуществом?
При действии ряда повреждающих факторов отмечается фенотипическая трансформация контрактильных миоцитов в секреторные. Данная трансформация часто наблюдается при повреждении интимы сосудов, формировании ее гиперплазии при развитии атеросклероза.
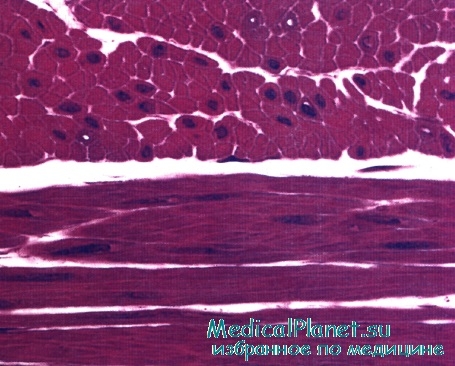
Гладкая мышечная ткань в поперечном (наверху) и продольном (внизу) разрезах. Обратите внимание на центрально расположенные ядра. Во многих клетках ядра не попали в срез.
Окраска: парарозанилин—толуидиновый синий. Среднее увеличение.
Гладкая мышечная ткань нервный аппарат
Волокна скелетной мышцы стимулируются исключительно нервными сигналами, но сокращения гладкой мышцы могут вызываться множеством типов сигналов: нервными, гормональными, растяжением мышцы и некоторыми другими. Основная причина различий состоит в том, что гладкомышечная мембрана содержит много типов рецепторных белков, способных запустить сократительный процесс. Кроме того, есть и другие рецепторные белки, тормозящие сокращение гладкой мышцы, что также отличает ее от скелетной мышцы. Обсудим сначала нервную регуляцию гладкомышечного сокращения, а затем гормональный контроль и другие способы управления.
Физиологическая анатомия нервно-мышечных соединений гладких мышц. Сложно организованные нервно-мышечные соединения, обнаруживаемые на волокнах скелетных мышц, в гладких мышцах не встречаются. Вместо этого вегетативные нервные волокна, иннервирующие гладкие мышцы, обычно диффузно разветвляются на наружной поверхности пласта мышечных волокон. В большинстве случаев эти волокна не входят в непосредственный контакт с клеточными мембранами гладкомышечных волокон, а формируют так называемые диффузные соединения, которые секретируют медиаторы в матрикс, покрывающий гладкую мышцу, часто на расстоянии от нескольких нанометров до нескольких микрометров от мышечных клеток; затем медиатор диффундирует к клеткам.
Более того, при наличии многих слоев мышечных клеток нервные волокна часто иннервируют только наружный слой, и возбуждение распространяется от этого наружного слоя к внутренним слоям путем проведения потенциалов действия по мышечной массе или путем дополнительной диффузии медиатора.
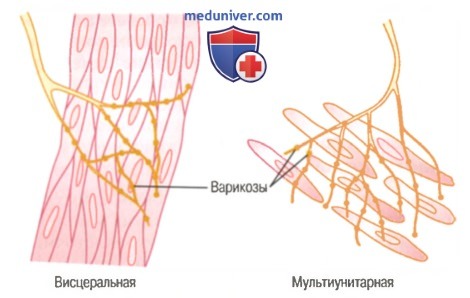
Иннервация гладких мышц.
Аксоны, иннервирующие гладкомышечные волокна, не имеют типичных ветвящихся окончаний, характерных для двигательной концевой пластинки волокон скелетной мышцы. Вместо этого большинство тонких концевых частей аксонов имеют множество варикозных расширений (варикозов), распределенных вдоль их осей. В этих местах связь между шванновскими клетками, окутывающими аксоны, прерывается, и медиатор может секретироваться через стенки варикозов. В варикозах находятся везикулы, которые подобно везикулам в концевой пластинке скелетных мышц содержат медиатор. Но в противоположность везикулам в соединениях скелетных мышц, которые всегда содержат ацетилхолин, везикулы окончаний вегетативных нервных волокон содержат ацетилхолин в одних волокнах и норадреналин — в других, а иногда и другие вещества.
В некоторых случаях, особенно в гладких мышцах мультиунитарного типа, варикозы отдалены от мембраны мышечных клеток на 20-30 нм, что равно ширине синаптической щели в соединении скелетной мышцы. Такие соединения называют контактными, и они функционируют во многом так же, как нервно-мышечные соединения скелетных мышц. Скорость сокращения этих гладкомышечных волокон значительно больше, чем у волокон, стимулируемых диффузными соединениями.
Возбуждающие и тормозящие медиаторы, секретируемые в нервно-мышечных соединениях гладких мышц. Самыми важными медиаторами, которые секретируются вегетативными нервами, иннервирующими гладкие мышцы, являются ацетилхолин и норадреналин, однако они никогда не выделяются одними и теми же нервными волокнами. Ацетилхолин для гладких мышц одних органов является возбуждающим медиатором, а на гладкие мышцы других органов действует как тормозящий агент. Если ацетилхолин возбуждает мышечное волокно, норадреналин обычно тормозит его. И наоборот, если ацетилхолин тормозит волокно, норадреналин, как правило, его возбуждает.
Но почему возникают такие разные реакции? Ответ заключается в том, что ацетилхолин и норадреналин возбуждают или тормозят гладкую мышцу, связываясь сначала с рецепторным белком на поверхности мембраны мышечной клетки. Некоторые из этих рецепторных белков являются возбуждающими рецепторами, тогда как другие — тормозящими рецепторами. Следовательно, тип рецептора определяет, как будет реагировать гладкая мышца — торможением или возбуждением, а также какой из двух медиаторов (ацетилхолин или норадреналин) будет проявлять возбуждающее или тормозящее действие.
Видео физиология гуморальной регуляции и ее отличие от гормональной — профессор, д.м.н. П.Е. Умрюхин
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021