
Гормональные регуляторы минерального гомеостаза костной ткани
Болезни костно-мышечной системы и соединительной ткани представляют важную медико-социальную проблему в связи с широкой распространенностью среди лиц трудоспособного возраста, занимая одно из ведущих мест в структуре общей и первичной заболеваемости населения Российской Федерации и других стран мира [10].
Одним из наиболее часто встречающихся в общетерапевтической и ревматологической практике вариантов наследственных нарушений соединительной ткани является синдром гипермобильности суставов (СГМС). По данным различных авторов, генерализованная суставная гипермобильность и СГМС отмечаются в популяции у 6–31,6% с вариабельностью в зависимости от возрастных, гендерных и этнических факторов [1, 5, 7].
Сочетание СГМС с костно-мышечными синдромами обусловлено облигатным вовлечением костной ткани в процесс дисплазии с разнообразием и полиморфизмом клинических проявлений. Более того, идентифицированные локомоторные симптомы относятся к согласованным критериям диагностики СГМС [5, 9].
При изучении костно-метаболических характеристик нарушения структуры и функции соединительной ткани у пациентов с СГМС особое значение приобретает интегральная оценка системных гормональных влияний, обеспечивающих регуляцию процессов костного и хрящевого ремоделирования.
Динамика противоположных, но взаимосвязанных и взаимообусловленных процессов костной резорбции и формирования, протекающих на протяжении всей жизни человека, составляет сущность цикла костного ремоделирования. Последовательно сменяющие друг друга фазы резорбции, реверсии и формирования кости генетически детерминированы и находятся под сложным многоуровневым нейрогуморальным контролем. На клеточные системы кости оказывают влияние гормоны, локальные тканевые факторы, сигналы межклеточных взаимодействий, центральные нейрогенные импульсы, обеспечивая постоянную репарацию и адаптацию костной ткани и ее собственную гормонально-метаболическую активность. Нарушение костного обмена и разобщение процессов ремоделирования способны вызвать остеопению и остеопороз [8].
Системные гормональные воздействия направлены в первую очередь на поддержание гомеостаза остеотропных минералов и их резервирование, что является важной функцией костной ткани. Удержание константы кальция в системной циркуляции, сопряженность его метаболизма в костной ткани, а также баланс внутри- и внеклеточных форм элемента регулируются системой «паратгормон (ПТГ) – кальцитонин – активная форма витамина D (кальцитриол)». При этом биологические эффекты ПТГ направлены на повышение уровня кальциемии за счет стимуляции почечной реабсорбции с одновременным снижением экскреции кальция с мочой. Синергизм ПТГ и кальцитриола по отношению к кальциемии реализуется через стимуляцию последним кишечной абсорбции кальция. Кроме того, кальцитриол стимулирует всасывание фосфора в тонком кишечнике и его канальцевую реабсорбцию в нефронах, тогда как ПТГ оказывает выраженный фосфатурический эффект. Фосфор стимулирует функциональную активность остеобластов, образующих белковые компоненты матрикса костной ткани; без него невозможна минерализация скелета. Модуляция синтеза ключевых гормонов-регуляторов фосфорно-кальциевого обмена – ПТГ и кальцитриола — происходит реципрокно за счет транскрипции соответствующих генов [2].
На клеточные системы, обеспечивающие костное ремоделирование, помимо кальций-регулирующих, оказывают влияние и другие гормоны эндокринной системы. Гуморальный сигналинг процессов костно-хрящевого роста, репарации и минерализации реализуется через основные – гормон роста, половые и так называемые пермиссивные гормоны – тиреоидные, кортикостероиды, инсулин.
Целью нашего исследования явилась оценка состояния некоторых облигатных и пермиссивных системных гормональных регуляторов костно-хрящевого ремоделирования у пациентов с синдромом гипермобильности суставов.
Материал и методы
Показатели минерального обмена и уровни гормонов, оказывающих влияние на костное и хрящевое ремоделирование, изучены у 80 пациентов (34 мужчин и 46 женщин) с СГМС в возрасте от 18 до 30 лет (средний возраст 22,9±3,8 лет). Гипермобильный синдром диагностировали по шкале Р. Beighton согласно действующим критериям [5, 9]. В исследование не включались больные с классифицируемыми наследственными заболеваниями. Изучали уровни кальция и фосфора в сыворотке крови, показатели суточной экскреции кальция и фосфора с мочой. С помощью иммунохемилюминисцентного анализа определяли уровни паратиреоидного гормона (ПТГ), 25(ОН) витамина D (25(ОН)D3), соматотропного гормона (СТГ), гормонов, характеризующих функцию щитовидной железы: тиреотропного (ТТГ), свободной фракции тироксина (сТ4), трийодтиронина (Т3), а также антитела к тиреоидным антигенам (тиреопероксидазе) – АТ к ТПО.
В контрольную группу были включены 25 человек, сопоставимых по полу и возрасту (средний возраст 22,3±3,6 лет), без внешних признаков дисплазии соединительной ткани, в том числе гипермобильного синдрома.
Полученные результаты статистически обработаны. Количественные данные представлены в виде средней и стандартной ошибки средней (M±m). Достоверность межгрупповых различий устанавливали с помощью критерия Ньюмена—Кейлса. Достоверность различий качественных признаков оценивали с помощью критерия χ2. Достоверными считали различия при p≤0,05.
Результаты и обсуждение
Выраженность суставной гипермобильности по шкале Beighton в 43,8% случаев соответствовала 5 баллам, у 20% обследуемых данный показатель составил 3 балла, у 26,2% – 4 балла. Значительные степени гипермобильности – 7 и 9 баллов — выявлены соответственно у 2,5% и 7,5% пациентов.
Оценка функции системы основных кальцийрегулирующих гормонов установила в группе пациентов с СГМС достоверное снижение среднего уровня ПТГ, однако значений, выходящих за пределы референсных, в обеих группах обследованных не отмечено (рис. 1).
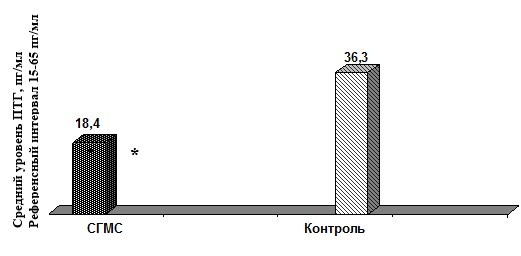
*р 0,05) в среднем результаты исследуемой группы соответствовали уровню «недостаточности» витамина D при референсном интервале 47,7–144,0 нмоль/л [2].
Выявленные в группе пациентов с СГМС особенности костного минерального обмена характеризовались незначительно сниженными или низконормальными показателями общего кальция крови на фоне сохраненного референсного значения фосфатемии. Гипокальциемия при этом отмечена у 23,8% обследуемых – достоверно (р 0,05).
Случаи изолированной гипотироксинемии в исследуемой группе, вероятно, могут быть расценены как сниженные адаптивно-приспособительные возможности у больных СГМС в условиях легкой йодной эндемии. Это указывает на необходимость применения более активных профилактических стратегий возмещения дефицита йода у молодых пациентов с СГМС.
Изолированное повышение титра антитиреоидных антител без нарушения функции железы и соответствующих ультразвуковых или гистологических феноменов, как известно, самостоятельного клинического значения не имеет и может определяться в 5–10% здоровой популяции, особенно среди женщин [6]. Вместе с тем носительство антител рассматривается как фактор риска аутоиммунных и деструктивных тиреопатий и развития стойкого гипотиреоза с целесообразностью динамического контроля функции щитовидной железы, особенно у женщин репродуктивного возраста.
Читайте также: Пути связывания аммиака в тканях биохимия
Классическая развернутая клиническая картина тиреоидита Хашимото, включающая наличие манифестного или субклинического гипотиреоза у иммуннокомпетентных лиц, наблюдалась в 11,25% случаев СГМС (р>0,05). Учитывая молодой возраст обследуемых, во всех случаях, в том числе и при субклиническом гипотиреозе, пациентам назначена заместительная тиреоидная терапия препаратами Левотироксина.
У 4 (5%) больных СГМС наблюдалось сочетание повышенного титра АТ к ТПО и соответствующих ультразвуковых феноменов при сохраненном эутиреоидном статусе, что по существующим клиническим рекомендациям позволило установить диагноз аутоиммунного тиреоидита без нарушения функции щитовидной железы [6] с необходимостью последующего динамического контроля тиреоидной функции.
Всего же повышенный титр антитиреоидных антител установлен у 24 (30%) пациентов с СГМС, что, безусловно, превышает популяционные показатели (р≤0,05). Таким образом, лица с гипермобильным синдромом имеют повышенный риск развития аутоиммунных тиреопатий и могут рассматриваться в качестве целевой группы для скрининга нарушений функции щитовидной железы.
Болезнь Грейвса отмечена у 2 (2,5%) пациентов с СГМС: ее распространенность существенно не отличалась от общепопуляционной [6]. Это позволило исключить тиреотоксикоз из числа факторов, приводящих к нарушению метаболических процессов и костного ремоделирования при гипермобильном синдроме.
Установленные особенности в состоянии системных гормональных регуляторов костного метаболизма способствуют пониманию механизмов развития остеопенического синдрома у пациентов с СГМС, расширяя представления о нарушениях костно-хрящевого ремоделирования при гипермобильном синдроме. Для своевременного выявления недостаточности витамина D с адекватным его восполнением и коррекции дефицита кальция необходимо включение показателей костного метаболизма в алгоритм обследования пациентов с СГМС. Также в группе больных с гипермобильным синдромом представляется целесообразным проведение тиреоидного скрининга.
Ягода А.В., д.м.н., профессор, заслуженный деятель науки РФ, заведующий кафедрой госпитальной терапии Ставропольского государственного медицинского университета, г. Ставрополь;
Чагай Н.Б., д.м.н., профессор кафедры эндокринологии, детской эндокринологии и диабетологии Ставропольского государственного медицинского университета, г. Ставрополь.
Метаболизм костной ткани и остеопороз
Рассмотрены подходы к выбору средств для профилактики и лечения потерь костной ткани, восстановления ее структуры и качества. Применяемый препарат должен способствовать синтезу коллагена, формированию костного матрикса, его минерализации и, соответственно
Approaches to selecting the methods of prevention and treatment of bone tissue losses, restoration of its structure and quality, were considered. The used preparation must contribute to collagen synthesis, formation of bone matrix, its mineralization, and, respectively, increase of the bone density and strength.
Остеопороз (ОП) — прогрессирующее системное заболевание скелета, характеризующееся снижением костной массы и нарушением микроархитектоники (качества) костной ткани, что приводит к хрупкости костей и повышению риска переломов. ОП — самое распространенное заболевание костной ткани: остеопоротические переломы отмечается у половины всех женщин, находящихся в периоде постменопаузы, а также у мужчин старших возрастных групп [1]. Очевидно, что рано начатые активные профилактические мероприятия у значительной части населения могут существенно повлиять на распространенность, прогрессирование и исходы заболевания, а также снизить риск переломов. В связи с этим изучение различных лекарственных препаратов и методов, применяемых для профилактики ОП, приобретает особый смысл.
Кость — специализированная разновидность соединительной ткани, состоящая из клеток и межклеточного вещества. В течение всей жизни основные функции костной ткани, такие как жесткость и гибкость, снижаются, поскольку с возрастом наблюдаются повреждение матрикса и потеря минералов. В противовес указанным проявлениям, в кости осуществляется ремоделирование — процесс, направленный на самостоятельное обновление и сохранение скелета как структурного и функционального органа.
Основными клетками костной ткани, функциями которой регулируется гомеостаз кости, являются остеобласты, остеокласты и остеоциты. Основной функцией остеобластов является создание органического межклеточного матрикса кости, остеоида. Остеобласты синтезируют и выделяют в окружающую среду фибриллы коллагена, протеогликаны и гликозаминогликаны. Наряду с этим остеобласты активно синтезируют и выделяют во внеклеточное пространство значительное количество глицерофосфолипидов, способствующих связыванию Ca 2+ и участвующих в процессах минерализации. Клетки сообщаются между собой через десмосомы, которые позволяют проходить Ca 2+ и цАМФ. Они также обеспечивают непрерывный рост кристаллов гидроксиапатитов и выступают в качестве посредников при связывании минеральных кристаллов с белковой матрицей.
В ходе формирования кости некоторые остеобласты оказываются замурованными в толщу матрикса и становятся остеоцитами. Остеоциты контактируют друг с другом через отростки, являются основными компонентами в сформировавшейся костной ткани. Основная функция остеоцитов — поддержание нормального состояния костного матрикса и баланса кальция и фосфора в организме.
Остеокласты — клетки, выполняющие функцию разрушения кости; развиваются из стволовой кроветворной клетки и являются специализированными макрофагами. В процессе ремоделирования кости резорбтивный стимул запускает процесс привлечения остеокластов к участку кости. Прикрепившись к кости, остеокласты продуцируют множество протеолитических ферментов и формируют полость в кальцинированном матриксе. Таким образом, они осуществляют непрерывный процесс резорбции и обновления костной ткани, обеспечивая необходимый рост и развитие скелета, структуру, прочность и упругость.
Важнейшим компонентом костной ткани является межклеточное вещество — уникальный комплекс органических и неорганических компонентов, заполняющих пространство между клетками. Минерализованный матрикс костной ткани поддерживает структуру скелета и под координирующим влиянием остеобластов и остеокластов обеспечивает резервуар как ионов, так и факторов роста, которые высвобождаются в процессе метаболизма.
Читайте также: Как сделать цветы гвоздики из ткани
Органический межклеточный матрикс костной ткани представлен семейством коллагеновых белков. Состав кости необычен тем, что фактически в ней представлен только коллаген I типа (90%), хотя наряду с коллагеном I типа в кости все же присутствуют следы других типов коллагена, таких как V, XI, XII. Скорее всего, что эти типы коллагена принадлежат другим тканям, которые и находятся в костной ткани, но не входят в состав костного матрикса. Например, коллаген V типа обычно обнаруживается в сосудах, которые пронизывают кость. Коллаген XI типа находится в хрящевой ткани и может соответствовать остаткам кальцифицированного хряща. Коллагеновые фибриллы в кости строго ориентированы в соответствии с распределенной функциональной нагрузкой на кость, что обеспечивает упругость и эластичность кости. Веретенообразные и пластинчатые кристаллы гидроксиапатита находятся на коллагеновых волокнах, в их пределах и в окружающем пространстве. Как правило, они ориентированы в том же направлении, что и коллагеновые волокна.
Неколлагеновая часть матрикса (10%) представлена основным веществом (витамин К-зависимыми глютамилпротеинами (остеокальцином), матричными протеинами, остеопонтином, остеонектином, фибронектином, фосфопротеидами, сиалопротеидами, а также протеогликанами).
Минеральные вещества, которыми пропитан органический матрикс, представлены главным образом кристаллами гидроксиапатита Ca10(PO4)6(OH)2. Кроме того, в кости обнаружены ионы Mg 2+ , Na + , K + , SO4 2- , HCO 3- , гидроксильные и другие ионы, которые могут принимать участие в образовании кристаллов.
Важно подчеркнуть, что ОП является результатом уменьшения органического матрикса кости, а вовсе не плохой кальцификацией костной ткани. При ОП существенно снижается скорость образования остеоида, необходимого для формирования кости. Поэтому при планировании профилактических мероприятий чрезвычайно важно учитывать потенциальную возможность препаратов, наряду с адекватной минерализацией, оказывать влияние на синтез органического матрикса.
Разумеется, качественная структура и прочность кости, ее эффективное функционирование и своевременное самообновление возможны лишь при адекватной обеспеченности макро- и микроэлементами, которые, подобно кальцию и витамину D, принимают непосредственное участие в биохимических процессах костной ткани [2–5]. Магний, медь, цинк, марганец, бор, являясь кофакторами ферментов, регулируют синтез костного матрикса, его минерализацию, а также равномерный рост, гибкость и прочность костной ткани. Известно, что дефицит этих веществ замедляет формирование костной массы в детстве и подростковом возрасте, способствует ее ускоренной потере в пожилом возрасте. Соответственно, дефицит любого из известных минеральных веществ в организме препятствует успешной терапии и профилактике нарушений структуры кости [6, 7].
Одним из основных минералов, играющих важную роль в формировании и поддержании структуры костной ткани, является кальций. Поскольку кальций не производится в организме, то для поддержания оптимальной концентрации он должен регулярно поступать извне. Причем желательно, чтобы его поступление в организм обеспечивалось за счет натуральных молочных продуктов, молока и его производных (кефира, простокваши, ряженки, йогурта, творога, сыра). Вместе с тем биодоступность кальция из пищи составляет порядка 30%, причем с высокой индивидуальной вариабельностью. Более того, у лиц пожилого возраста нередко имеет место непереносимость молочных продуктов, связанная со снижением концентрации лактазы в желудочном соке, что приводит к низкому потреблению кальция.
Согласно эпидемиологическим исследованиям, среди женщин в возрасте старше 45 лет, проживающих в мегаполисах, непереносимость молока встречается с частотой 25,0–34,0%. При этом достаточное потребление кальция с продуктами питания имеет место менее чем у 5% женщин [8]. Фактически содержание кальция в пищевом рационе постменопаузальных женщин не соответствует рекомендованным нормам. Очевидно, что обеспечение должного уровня потребления кальция возможно лишь при условии дополнительного регулярного назначения медикаментозных препаратов.
Витамин D — основной регулятор активной абсорбции кальция в организме. Витамин D относят к группе жирорастворимых витаминов. Хотя в отличие от всех других витаминов он биологически не активен. В активную, гормональную, форму он превращается за счет двухступенчатой метаболизации в организме и оказывает многообразные биологические эффекты за счет взаимодействия со специфическими рецепторами, локализованными в ядрах клеток тканей и органов. Другое дело — активный метаболит витамина D. Он действует как истинный гормон, хотя в научной литературе его традиционно называют витамином D [9, 10].
Природная форма витамина D — витамин D2 (эргокальциферол) поступает в организм человека в относительно небольших количествах — не более 20–30% от потребности. В основном из злаковых растений, рыбьего жира, сливочного масла, маргарина, молока, яичного желтка и др. В организме витамин D2 метаболизируется с образованием производных, обладающих сходным с метаболитами витамина D3 действием.
Еще одна природная форма витамина D — витамин D3, или холекальциферол, является ближайшим аналогом витамина D2, но его синтез мало зависит от поступления извне. Холекальциферол образуется в организме позвоночных животных, в том числе амфибий, рептилий, птиц и млекопитающих, в связи с чем играет значительно бóльшую роль в процессах жизнедеятельности человека, чем поступающий в небольших количествах с пищей витамин D2. В организме витамин D3 образуется из находящегося в дермальном слое кожи предшественника (7-дегидрохолестерина) под влиянием коротковолнового ультрафиолетового облучения спектра В (УФ–В/солнечного света, длина волны 290–315 нм) при температуре тела в результате фотохимической реакции раскрытия В-кольца стероидного ядра и термоизомеризации, характерной для секостероидов [9, 10].
В последующем поступивший с пищей и/или образовавшийся в организме в процессе эндогенного синтеза витамин D подвергается реакции 25-гидроксилирования в печени. Важно, что гидроксилирование витамина D3 в печени представляет собой полностью субстратзависимый процесс, который протекает весьма быстро и ведет к повышению уровня 25(ОН)D в сыворотке крови. Уровень этого вещества отражает как образование витамина D в коже, так и его поступление с пищей, в связи с чем может использоваться как маркер статуса витамина D [9, 10].
Читайте также: Мебель арт ткани для мебели
Вторая реакция гидроксилирования 25(ОН)D, с образованием наиболее важной, качественно и количественно значимой активной гормональной формы — 1a,25-дигидроксивитамина D3 (1α,25(ОН)2D3), называемой также D-гормоном, кальцитриолом, протекает уже в основном в почках, в клетках проксимальных отделов канальцев коры почек при участии фермента 1α-гидроксилазы (CYP27В1). Этот процесс строго регулируется рядом эндогенных и экзогенных факторов. Во-первых, регуляция синтеза 1a,25(ОН)2D3 в почках является непосредственной функцией паратиреоидного гормона (ПТГ), на концентрацию которого в крови, в свою очередь, по механизму обратной связи оказывают влияние как уровень самого активного метаболита витамина D3, так и концентрация кальция и фосфора в плазме крови. Во-вторых, активация синтеза 1a-гидроксилазы и реакции 1a-гидроксилирования зависит от половых гормонов (эстрогенов и андрогенов), кальцитонина, пролактина, гормона роста (через ИПФР-1) и др. В-третьих, ингибирующее влияние на активность 1a-гидроксилазы оказывают глюкокортикостероидные гормоны, 1α,25(ОН)2D3 и ряд его синтетических аналогов. Фактор роста из фибробластов (FGF23), секретируемый в клетках кости, вызывает образование натрий-фосфат-котранспортера, который действует в клетках почек и тонкого кишечника, оказывает тормозящее влияние на синтез 1,25-дигидроксивитамина D3. На метаболизм витамина D оказывают влияние и некоторые лекарственные средства, например, противоэпилептические препараты.
Основными реакциями, в которых участвует D-гормон, являются абсорбция кальция в желудочно-кишечном тракте и его реабсорбция в почках. D-гормон усиливает кишечную абсорбцию кальция в тонком кишечнике за счет взаимодействия со специфическими РВD. Об эффективности данного механизма свидетельствует тот факт, что без участия витамина D лишь 10–15% пищевого кальция и 60% фосфора абсорбируются в кишечнике. Взаимодействие между 1a,25-дигидроксивитамином D3 и РВD повышает эффективность кишечной абсорбции Са 2+ до 30–40%, т. е. в 2–4 раза, а фосфора — до 80%. Сходные механизмы действия D-гормона лежат в основе осуществляемой под его влиянием реабсорбции Са 2+ в почках.
В костях 1α,25(ОН)2D3 связывается с рецепторами на кость-формирующих клетках — остеобластах, вызывая повышение экспрессии ими лиганда рецептора активатора ядерного фактора кВ (RANKL). Рецептор-активатор ядерного фактора кВ (RANK), являющийся рецептором для RANKL, локализованным на преостеокластах, связывает RANKL, что вызывает быстрое созревание преостеокластов и их превращение в зрелые остеокласты. В процессах костного ремоделирования зрелые остеокласты резорбируют кость, что сопровождается выделением кальция и фосфора из минерального компонента (гидроксиапатита) и обеспечивает поддержание уровня кальция и фосфора в крови. В свою очередь, адекватный уровень кальция (Са 2+ ) и фосфора необходим для нормальной минерализации скелета [11–13].
Многочисленные исследования показали, что назначение препаратов кальция и/или витамина D способствует уменьшению потери костной ткани [14–19]. У женщин в поздней постменопаузе с низким употреблением пищевого кальция прием кальция предотвращает потерю костной ткани в позвоночнике [20, 21]. В свою очередь, назначение добавок кальция лицам старше 60 лет приводит к снижению потери костной массы в области бедра среди белых мужчин и женщин в возрасте моложе 72 лет [22]. Эффект назначения цитрата кальция на минеральную плотность кости (МПК) у женщин в раннем (до 5 лет) и среднем (от 5 до 10 лет) постменопаузальном периоде в течение двух лет проявлялся в виде прироста МПК в поясничном отделе на 1%, наряду со значимым снижением МПК на 2,4% в группе, получавшей плацебо [23]. Метаанализ 9 рандомизированных клинических исследований с общей выборкой более 50 тыс. человек, в 6 из которых сравнивалось комбинированное лечение витамином D (400 или 700–800 МЕ/сут) и кальцием с группами плацебо или без лечения, продемонстрировал достоверное снижение риска перелома бедра на 18% (RR 0,82 [95% ДИ 0,71–0,94], р = 0,0005) и риска внепозвоночных переломов на 12% (RR 0,88 [95% ДИ 0,78–0,99], р = 0,036) в группах, получавших комбинированную терапию, по сравнению с группами без добавок [24]. В исследованиях, где применялся витамин D в дозе 700–800 МЕ/сут, эффект на риск перелома бедра был выше, чем при приеме 400 МЕ (21% и 18% соответственно). Соответственно, в исследованиях, в которых пациенты получали только витамин D или плацебо (4 РКИ с общей численностью 9083 пациента), не было получено снижения риска внепозвоночных переломов как при применении дозы 400 МЕ (RR 1,14 [95% ДИ 0,87–1,49]), так при использовании 700–800 МЕ (RR 1,04 [95% ДИ 0,75–1,46]), что подтверждает ранее представленные данные о том, что витамин D без добавления кальция не снижает риск переломов [24].
Магний
Известно, что 60–65% магния находится именно в скелете и от обеспеченности костей магнием зависит обмен кальция и витамина D. Являясь структурным компонентом значительного числа ферментов, магний образует кристаллы с фосфатами, принимает участие в росте и стабилизации кристалла гидроксиапатита — структурной единицы минерального компонента костной ткани [25, 26]. Магний регулирует секрецию паратгормона (ПГ), повышает чувствительность клеток-мишеней к ПГ и витамину D, стимулирует действие кальцитонина. Длительное во времени нарушение соотношения Mg/Ca в сторону дефицита магния сопровождается замедлением обменных процессов в кости. Специальные магний-дефицитные диеты, сопровождающиеся уменьшением сывороточной концентрации магния, способствуют системной потере костной массы, снижению толщины надкостницы, характерным изменениям провоспалительных маркеров и маркеров резорбции кости. Уже по истечении достаточно короткого срока (4 недели), магний-дефицитная диета приводит к значимому снижению содержания минеральных веществ кости (р
М. И. Шупина, кандидат медицинских наук
Г. И. Нечаева 1 , доктор медицинских наук, профессор
Д. В. Шупин
Е. В. Надей
А. А. Семенкин, доктор медицинских наук, профессор
ГБОУ ВПО ОмГМУ МЗ РФ, Омск
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом