
Действие инсулина на клетки-мишени начинается после его связывания со специфическими димерными мембранными рецепторами (рис. 6.22), при этом внутриклеточный домен рецептора обладает тирозинкиназной активностью. Инсулин-рецепторный комплекс не только передает сигнал внутрь клетки, но и частично путем эндоцитоза поступит внутрь клетки к лизо-сомам. Под влиянием лизосомальной протеазы инсулин отщепляется от рецептора, при этом последний либо разрушается, либо возвращается к мембране и вновь встраивается в нее. Многократное перемещение рецептора от мембраны к лизосомам и обратно к мембране носит название рециклизация рецептора. Процесс рециклизации важен для регуляции колич-ства инсулиновых рецепторов, в частности обеспечения обратной зависимости между концентрацией инсулина и количеством мембранных рецепторов к нему.
Образование инсулин-рецепторного комплекса активирует тирозинкиназу, запускающую процессы фосфорилирования внутриклеточных белков. Происходящее при этом аутофосфорилирование рецептора ведет к усилению первичного сигнала. Инсулин-рецепторный комплекс вызывает активирование фосфолипазы С, образование вторичных посредников инозитолтрифосфата и диацилглицерола, активацию протеинкиназы С, ингибирование цАМФ. Участие нескольких систем вторичных посредников объясняет многообразие и различия эффектов инсулина в разных тканях.
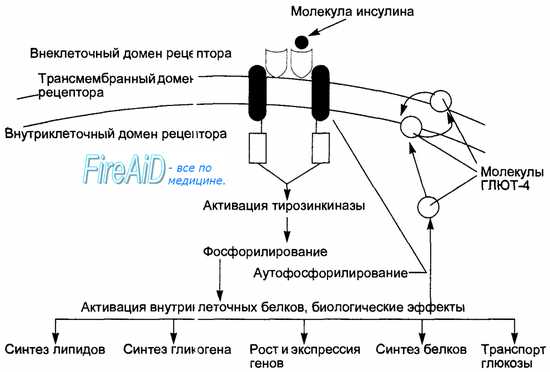
Рис. 6.22. Схема механизма действия инсулина на клетку-мишень.
Инсулин оказывает влияние на все виды обмена веществ, способствует анаболическим процессам, увеличивая синтез гликогена, жиров и белков, тормозя эффекты многочисленных контринсулярных гормонов (глюкагона, катехоламинов, глюкокортикоидов и соматотропина). Все эффекты инсулина по скорости их реализации подразделяют на 4 группы: очень быстрые (через несколько секунд) — гиперполяризация мембран клеток (за исключением гепатоцитов), повышение проницаемости для глюкозы, активация Na-K-АТФазы, входа К+ и откачивания Na , подавление Са-насоса и задержка Са2+; быстрые эффекты (в течение нес кольких минут) — активация и торможение различных ферментов, подавляющих катаболизм и усиливающих анаболические процессы; медленные процессы (в течение нескольких часов) — повышенное поглощение амиминокислот, изменение синтеза РНК и белков-ферментов; очень медленные эффекты (от часов до суток) — активация митогенеза и размножения клеток.
Важнейшим эффектом инсулина в организме является увеличение в 20—50 раз транспорта глюкозы через мембраны мышечных и жировых клеток путем облегченной диффузии по градиенту концентрации с помощью чувствительных к гормон) мембранных белковых переносчиков, называемых ГЛЮТ. В мембранах разных видов клеток выявлены 6 типов ГЛЮТ (рис. 6.23), но только один из них — ГЛЮТ-4 — является инсулинозависимым и находится в мембранах клеток скелетных мышц, миокарда, жировой ткани.
Инсулин влияет на угле водный обмен, что проявляется:
1) активацией утилизации глюкозы клетками,
2) усилением процессов фосфорилирования;
3) подавлением распад; и стимуляцией синтеза гликогена;
4) угнетением глюконеогенеза;
5) активацией процессов гликолиза;
6) гипогликемией.
Действие инсулина на белковый обмен состоит в: 1) повышении проницаемости мембран для аминоокислот; 2) усилении синтеза иРНК; 3) активации в печени синтеза aминокислот; 4) повышении синтеза и подавлении распада белка.
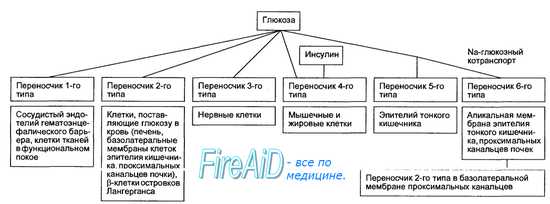
Рис. 6.23. Схема транспорта глюкозы через клеточные мембраны. Переносчики имеют общее название ГЛЮТ-1, 2, 3, 4, 5, 6. Только ГЛЮТ-4 является инсулинозависимым.
Основные эффекты инсулина на липидный обмен:
• стимуляция синтеза свободных жирных кислот из глюкозы;
• стимуляция синтеза липопротеиновой липазы в клетках эндотелия сосудов и благодаря этому активация гидролиза связанных с липо-протеинами крови триглицеридов и поступления жирных кислот в клетки жировой ткани;
• стимуляция синтеза триглицеридов;
• подавление распада жира;
• активация окисления кетоновых тел в печени.
Благодаря влиянию на клеточную мембрану инсулин поддерживает высокую внутриклеточную концентрацию ионов калия, что необходимо для обеспечения нормальной в возбудимости клеток.
Широкий спектр метаболических эффектов инсулина в организме свидетельствует о том, что гормон необходим для осуществления функционирования всех тканей, оргаганов и физиологических систем, реализации эмоциональных и поведенческих актов, поддержания гомеостазиса, осуществления механизмов приспособления и защиты организма от неблагоприятных факторов среды.
Недостаток инсулина (относительный дефицит по сравнению с уровнем контринсулярных гормонов, прежде всего глюкагона) приводит к сахарному диабету. Избыток инсулина в крови, например при передозировке, вызывает гипогликемию с резкими нарушениями функций центральной нервной системы, использующей глюкозу как основной источник энергии независимо от инсулина.
Механизмы восприятия инсулина тканями организма

Состояние клеточных мембран
Общими чертами биологических мембран, важными для их функционирования, являются текучесть, способность к конформационным перестройкам и фазовым переходам. Развитие сахарного диабета 1 и 2 типа сопровождается увеличением жесткости клеточных мембран. Причиной этого являются не только интенсификация процессов перекисного окисления липидов, но и погружение части поверхностных белков в липидный слой с изменением топографии мембранного гликокаликса При снижении дозы инсулина больным, имеющим низкий показатель экспонирования белков на фоне гиперлактацидемии, состояние мембран улучшалось: показатель экспонирования белков увеличивался и отмечалась тенденция к снижению повышенного параметра упорядоченности мембран [Вахрушева Л.Л. и др., 2004].
Эритроциты больных сахарным диабетом помимо изменения морфологии теряют способность к проникновению в поры фильтров, изменяется скорость и характер растекания плазмы и сыворотки в порах фильтров [Воинов Д.А. и др., 2003]. При оценке электронного парамагнитного резонанса с использованием в качестве зонда короткоцепочечного остатка жирной кислоты СдР9 с иминоксильным фрагментом обнаружилось, что образцы крови больных сахарным диабетом теряют способность взаимодействия с зондом (зонд не встраивается в липидный слой мембраны) [Воинов Д.А. и др., 2003].
Подобные изменения цитоплазматических мембран клеток гепатоцитов, почечной ткани, скелетных мышц, легочной ткани наблюдались при экспериментальном сахарном диабете [Микаэлян Н.П. и др., 1999].
Параметры инсулинорезистентности зависят от степени пероксидации фосфолипидов в липидном бислое мембран. Экспериментальный сахарный диабет характеризуется повышением гидролитической и трансферазной активность эндогенных липолитических ферментов. Активность фосфолипазы Д повышается при образовании холина и зависит от тяжести течения сахарного диабета [Расулова В.Б., Рахимова Н.М., 2004].
Строение и кинетика инсулинового рецептора
Своё действие на уровне клетки инсулин осуществляет через соответствующий рецептор. Анализу сведений о локализации, химической природе, обмене и функциональной активности рецепторов инсулина в норме и патологии посвящён ряд фундаментальных работ [Комиссаренко В.П. и соавт.,1984, Сергеев П.В., Шимановский Н.Л., 1987, Микаелян Н.П., 1988, 1990]. Инсулиновый рецептор подробно исследован с помощью биохимических методов и технологии рекомбинантных ДНК. Рецептор инсулина (РИ) представляет собой тирозиновую протеинкиназу, т.е. протеинкиназу, фосфорилирующую белки по ОН-группе остатков тирозина. Это гликопротеин, построенный из двух a — субъединиц (130 кДа) и двух b -субъединиц (95 кДа); первые расположены целиком вне клетки, на ее поверхности, вторые пронизывают плазматическую мембрану.
Рецептор инсулина постоянно синтезируется и распадается; его период полужизни составляет 7–12 ч. Рецептор синтезируется в виде одноцепочечного пептида в шероховатом эндоплазматическом ретикулуме и быстро гликозируется в аппарате Гольджи. Предшественник человеческого рецептора инсулина состоит из 1382 аминокислот, его мол.масса составляет 190000, при расщеплении он образует зрелые a — и -b субъединицы. У человека ген инсулинового рецептора локализован в хромосоме 19. Инсулиновый рецептор имеет высоко консервативную структуру, еще более консервативную, чем структура самого инсулина.
Строение инсулинового рецептора, способность различных инсулинов связываться с рецепторами и вызывать биологические реакции, практически идентичны в клетках всех типов и всех видов.
Рецептор к инсулину с высокой специфичностью распознает в молекуле места связывания инсулина и осуществляет комплексирование с ним; опосредует передачу соответствующего сигнала, направленного на активацию внутриклеточных обменных процессов; осуществляет эндоцитоз гормонального комплекса, что приводит к лизосомальному протеолизу инсулина с одновременным возвращением субъединицы к мембране клетки.
Комиссаренко В.П. и соавт., (1984) выделяет три основные функции рецептора инсулина:
- Питание клеток (увеличение потока питательных веществ внутрь клеток и их утилизацию во всех направлениях, приводящих к преобладанию анаболических процессов над катаболическими);
- Обеспечение транспорта инсулина с эритроцитами крови к тканям;
- Обеспечение перехода инсулина из крови через гистогематические барьеры в межклеточную жидкость.
Взаимодействие гормона с рецепторами характеризуется быстротой, обратимостью, зависимостью от температуры, величины рН, присутствия одновалентных и двухвалентных катионов и гуаниновых нуклеотидов, кооперативностью и наличием специфических и неспецифических участков связывания. При взаимодействии инсулина с рецепторами имеет место отрицательная и положительная кооперативность связывания. Отрицательная кооперативность, сопровождающаяся снижением сродства рецепторов к гормону в 10 раз, обусловлена увеличением скорости диссоциации комплекса гормон-рецептор, снижением размеров солюбилизированного рецептора.
Инсулиновые рецепторы расположены в каких тканях
а) Активация инсулином рецепторов клеток-мишеней и результирующие ответы клеток. Для воздействия на клетку-мишень инсулин прежде всего связывается и активирует рецептор, расположенный на мембране клеток, — белок с молекулярной массой около 300000 (для облегчения понимания просим вас изучить рисунок ниже).
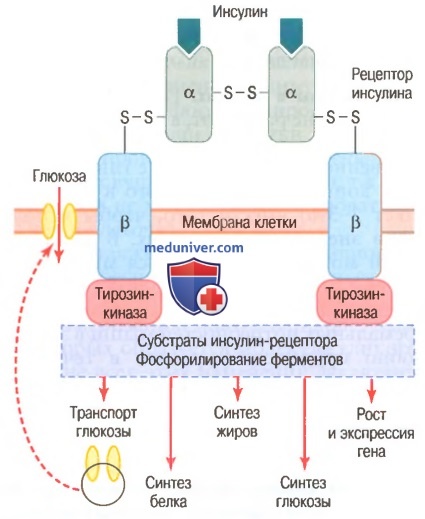
Схема инсулин-рецептора. Инсулин связывается с альфа-субъединицей рецептора, что вызывает фосфорилирование бета-субъединицы рецептора.
Это, в свою очередь, индуцирует активность тирозинкиназы. Рецептор тирозинкиназной активности запускает каскад реакций фосфорилирования в клетке, что увеличивает или снижает активность ферментов, включая субстраты инсулин-рецептора, которыми опосредованы влияния глюкозы на метаболизм глюкозы, жиров и белков.
Например, транспортеры глюкозы продвигаются к мембране клетки, облегчая поступление глюкозы в клетку
Активация рецептора сопровождается определенной последовательностью реакций.
Рецептор инсулина состоит из 4 субъединиц, связанных дисульфидными связями: 2 альфа-субъединицы располагаются практически снаружи мембраны и 2 бета-субъединицы прободают мембрану и продолжаются в цитоплазму. Инсулин взаимодействует с 2 наружными субъединицами, но благодаря существенным собственным связям субъединиц между собой внутренние бета-субъединицы при этом аутофосфорилируются, поэтому инсулиновый рецептор является примером энзим-связанного рецептора.
Аутофосфорилирование бета-субъединиц рецептора приводит к местной активации тирозинкиназы, которая в итоге вызывает фосфорилирование многих других внутриклеточных ферментов, включая группу, названную субстратами инсулин-рецептора. Различные типы СИР (например, СИР1, СИР2, СИРЗ) представлены в различных клетках. Система действует, активируя одни ферменты одновременно с инактивацией других. Таким образом, инсулин управляет внутриклеточными метаболическими механизмами, вызывая желательные реакции обмена белков, жиров и углеводов. Окончательные эффекты действия инсулина следующие.
1. Через несколько секунд после взаимодействия инсулина с рецептором проницаемость мембран для глюкозы у 80% клеток организма заметно повышается. Это особенно справедливо для мышечных клеток и адипоцитов, но совершенно не распространяется на нейроны мозга. Поступившая в клетки глюкоза фосфорилируется и становится субстратом для всех обычных этапов обмена углеводов.
Увеличение транспорта глюкозы в клетку предположительно является результатом перемещения множества внутриклеточных пузырьков к мембране клетки; эти пузырьки несут на своей мембране множество белков, обеспечивающих транспорт глюкозы; взаимодействуя с мембраной клетки, они облегчают поступление глюкозы в клетку. Если инсулина уже не оказывается в наличии, пузырьки отделяются от мембраны в течение 3-5 мин и возвращаются в середину клетки, сохраняя способность к использованию их по мере необходимости.
2. Мембрана становится более проницаемой для многих аминокислот, ионов калия, фосфатов, повышая транспорт этих веществ в клетку.
3. Медленные эффекты возникают через 10-15 мин и заключаются в изменении уровня активности большинства внутриклеточных метаболических ферментов. Этот эффект является результатом главным образом изменения процессов фосфорилирования ферментов.
4. Более медленные процессы продолжаются в течение нескольких часов и дней. Они являются результатом изменения скорости процесса трансляции на рибосомах и приводят к образованию новых белков. Еще более медленные эффекты опосредованы изменением скорости транскрипции ДНК в ядре клетки. Таким способом инсулин формирует многие ферментативные механизмы клеток для выполнения свойственных им метаболических задач.
Видео физиология гормонов поджелудочной железы и регуляции уровня глюкозы — профессор, д.м.н. П.Е. Умрюхин
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Инсулин в патогенезе центральной инсулинорезистентности, сахарном диабете 2 типа и болезни Альцгеймера
Долгое время мозг считался нечувствительным к инсулину органом, однако последние исследования свидетельствуют о ключевой роли инсулиновой сигнализации в подержании гомеостаза глюкозы, контроле клеточной пролиферации и регуляции когнитивных процессов.
Авторы: Enrique Blázquez, Esther Velázquez, Verónica Hurtado-Carneiro and Juan Miguel Ruiz-Albusac
Перевод: Ирина Буянова. Редакция: Илья Левашов.
Аннотация
Инсулин может попадать в мозг как из периферических тканей, так и синтезироваться клетками центральной нервной системы (ЦНС). Молекулярные механизмы действия инсулина в ЦНС не отличаются от периферического действия гормона. Результаты последних исследований демонстрируют нейротрофический, нейромодуляторный и нейропротективный эффекты инсулина и его основополагающую роль в регуляции гомеостаза глюкозы, репродуктивной и когнитивных функций, пищевого поведения, а также пролиферации и дифференцировке клеток.
Нарушение механизмов инсулиновой сигнализации в мозге лежит в основе патогенеза центральной инсулинорезистентности, сахарного диабета 2 типа (СД2) и болезни Альцгеймера (БА). Кроме того, появляется все больше данных о связи между нарушением метаболизма глюкозы в ЦНС и риском БА.
Общими звеньями патогенеза БА и СД2 являются митохондриальная дисфункция и нарушение энергетического гомеостаза, оксидативный стресс, образование модифицированных липопротеинов низкой плотности (ЛПНП), снижение активности O-GlcNAc трансферазы, формирование амилоидных бляшек, изменение метаболизма бета-амилоидного пептида (Aβ) и повышение фосфорилирования тау-белка.
Введение
Поддержание гомеостаза глюкозы в периферических тканях – одна из основных функций инсулина. В последнее время появляется все больше данных о наличии рецепторов к инсулину (ИР) в разных отделах ЦНС и их роли в регуляции важнейших физиологических функций, включая нейрональное развитие, метаболизм глюкозы, пищевое поведение и когнитивные процессы, а также в патогенезе нейродегенеративных заболеваний (1).
Как инсулин попадает в мозг
Присутствие инсулина в нервной ткани впервые было описано в работе Havrankova et al. в 1978 году и подтверждено многочисленными исследованиями на животных (2, 3). Исследователи показали, что уровень инсулина в ЦНС не зависит от концентрации инсулина в периферических тканях и превышает его в 10-100 раз (4).
Присутствие инсулина в цереброспинальной жидкости (ЦСЖ) служит биомаркером проницаемости гематоэнцефалического барьера (ГЭБ), который определяет эффективность проникновения гормона в ЦНС и препятствует его быстрому выведению (5, 6). Считается, что основным источником инсулина в ЦНС служат специфические популяции нейронов, хотя, как отмечает Havrankova et al. (2), не стоит исключать проникновение инсулина из периферического кровотока и через ГЭБ.
Инсулин периферического происхождения
В 1967 году Margolis и Altszuler впервые высказали гипотезу о наличии механизма, опосредующего транспорт инсулина через ГЭБ (7). В эксперименте на крысах авторы работы показали, что внутривенное введение инсулина вызывает незначительное повышение его уровня в ЦСЖ. Это привело к предположению, что инсулин может проникать через ГЭБ посредством рецепторзависимого транспорта (8). Выраженное повышение уровня инсулина в крови и незначительное увеличение его концентрации в ЦСЖ в ответ на системное введение гормона указывает на наличие специальной транспортной системы, позволяющей доставлять инсулин через ГЭБ. Хотя до сих пор неизвестно, являются ли переносчики инсулина и ИР в ЦНС представителями одного семейства белков, данные многочисленных исследований демонстрируют (9) сходные физико-химические свойства (насыщаемость, специфичность, афинность, кинетику распада и иммунонейтрализацию) этих белков (10, 11). С другой стороны, активность транспортеров ГЭБ может меняться в зависимости от региональных различий в проницаемости к инсулину и уровню гормона в крови. Наиболее высокое содержание инсулина обнаружено в мосте, продолговатом мозге и гипоталамусе; самый низкий уровень гормона показан в нейронах затылочной коры и таламуса (12). Транспорт инсулина через ГЭБ регулируется различными физиологическими факторами, например, действием глюкокортикоидных гормонов (13), и обладает высокой чувствительностью к изменению энергетического гомеостаза при голодании (14), ожирении (15), гибернации (16), а также в процессе старения и у пациентов с СД и БА (17, 18).
Инсулин центрального происхождения
Биосинтез инсулина в ЦНС хорошо изучен и был продемонстрирован в ряде экспериментальных исследований.
Определение уровня С-пептида в мозге
В исследованиях post mortem показано высокое содержание иммунореактивного инсулина и С-пептида в различных структурах мозга, в несколько раз превышающее их содержание в периферической крови (19). Уровень С-пептида в сыворотке крови и структурах мозга значительно снижается после 72 часов голодания и повышается в ответ на пероральное введение глюкозы (20). Прямое доказательство связи между уровнем С-пептида и снижением количества ИР в ЦНС было получено в исследовании с участием пожилых пациентов с БА, у которых наиболее высокий уровень С-пептида был обнаружен в коре головного мозга (21).
Читайте также: Что такое активное крашение ткани
С помощью метода гибридизации in situ удалось выделить мРНК инсулина из нейронов перивентрикулярного ядра гипоталамуса крыс (22). Кроме того, мРНК инсулина II присутствует в мозге крыс на всех этапах развития, что указывает на раннюю экспрессию гена-предшественника в мозге (23). У новорожденных кроликов показано наличие мРНК инсулина в CA1 и CA3 областях гиппокампа, зубчатой извилине и клетках гранулярного слоя обонятельной луковицы (24).
Экспериментальные подходы к работе с культурами клеток мозга
Многочисленные данные о механизмах синтеза инулина в ЦНС были получены в исследованиях на культурах клеток. Показано, что инкубация нейронов в растворе циклогексимида приводила к 80%-му снижению количества инсулин-иммунореактивных клеток в мозге крыс (25). В эмбриональных нейронах мышей было показано наличие двух форм иммунореактивного инсулина (ИРИ). Под действием трипсина про-инсулин расщепляется на компоненты, схожие по структуре с панкреатическим инсулином (26). Данные иммуногистохимического анализа и гибридизации in situ демонстрируют способность эмбриональных нейронов экспрессировать и выделять инсулин-подобные мРНК и инсулин-подобные белки, схожие по структуре с инсулином периферического происхождения (27). В мозге млекопитающих синтез иммунореактивного инсулина происходит в только нейронах ЦНС, но не в астроцитах (24).
Молекулярные механизмы, участвующие в регуляции образования и секреции инсулина в ЦНС, сходны с механизмами синтеза инсулина в периферических тканях. Одним из общих звеньев центрального и периферического путей образования инсулина является деполяризация, вызванная активацией АТФ-чувствительных K+ каналов (28). Связывание гормонов или глюкозы с рецепторами на поверхности клеточных мембран вызывает деполяризацию и стимулирует высвобождение инсулина. В культурах нейронов (но не астроцитов) циклогексимид ингибирует экзоцитоз, индуцируемый деполяризацией (29). Кроме того, в мозге взрослых крыс повышение мембранного потенциала способствует высвобождению инсулина.
Эффективность экзоцитоза инсулина во многом зависит от внутриклеточного содержания уровня кальция. Кальций-зависимый экзоцитоз специфичен для синаптических везикул нервных окончаний, что подтверждает данные о синтезе инсулина в нейронах ЦНС и его хранении в составе синаптических везикул (30).
Секреция инсулина из синаптосом повышается в ответ на увеличение уровня глюкозы, а добавление ингибитора гликолитического метаболизма – йодацетиловой кислоты (ЙК) – в культуры клеток мозга вызывает снижение высвобождения ИРИ на 50%, что подчеркивает регуляторную роль метаболизма глюкозы в секреции инсулина в ЦНС (31).
Таким образом, результаты последних исследования подтверждают наличие центрального пути синтеза инсулина (32).
Влияние инсулина на эндотелиальные клетки и ГЭБ
ГЭБ образован особым типом эндотелиальных клеток (33), которые формируют физический барьер между клетками мозга и кровотоком (17). Поверхность клеток эндотелия содержит инсулин-связывающие сайты, которые выполняют транспортную и рецепторную функции (34). Эти инсулин-связывающие сайты играют ключевую роль в регуляции внутриклеточных каскадов в клетках ГЭБ и инсулиновой сигнализации, а также способствуют повышению эффективности транспорта тирозина и триптофана (35), азидотимидина (36) и лептина (37) из периферической крови в мозг.
Кроме того, инсулин оказывает регуляторное действие на экспрессию и активность некоторых транспортных белков. Так, инсулин повышает экспрессию гликопротеина-Р (170-кДа), играющего важную роль в поддержании целостности ГЭБ (38), и подавляет экспрессию и активность белка лекарственной устойчивости рака молочной железы (39). Нейрохимические изменения в капиллярах мозга в ответ на действие инсулина реализуются через снижение активности щелочной фосфатазы (40). Вместе с тем наблюдается активация элемента антиоксидантного ответа 4 и увеличение экспрессии каталитической субъединицы глутаматцистеин лигазы (41). Инсулин-зависимое снижение активности серотониновых рецепторов подсемейства 5-HT2c в сосудистом сплетении мозга указывает на роль MAP-киназного пути в реализации эффекта серотонинового рецептора (42).
.png)
Рисунок 1. Проникновение инсулина через ГЭБ опосредовано действием белков-переносчиков
Ферменты, участвующие в деградации инсулина, также в большом количестве присутствуют в нейронах коры больших полушарий, гиппокампа, мозжечка и ствола, а также в олигодендроцитах, сосудистом сплетении и эндотелиальных клетках (43). При низком уровне пептида Aβ наблюдается увеличение активности ферментов, разрушающих инсулин, что указывает на их роль в катаболизме Aβ и может стать терапевтической мишенью при разработке подходов к лечению нейродегенеративных заболеваний (44).
Механизмы действия инсулина в ЦНС
Инсулиновые рецепторы в мозге
Ген ИР расположен на хромосоме 19p13.2–19p13.3 и содержит 22 экзона, 11 из которых кодируют α и β субъединицы белка. В результате альтернативного сплайсинга +/− экзона 11 образуются две изоформы белка-предшественника – ИР-B и ИР-A, соответственно. Этот экзон кодирует небольшую последовательность аминокислот, расположенных на С-терминальном конце внеклеточного домена α-субъединицы рецептора (45). ИР-B содержатся преимущественно в инсулин-чувствительных тканях человека – скелетных мышцах, адипоцитах и печени, в то время как рецепторы типа А преобладают в структурах ЦНС (46–48).
Гетеротетрамер ИР содержит два лиганд-связывающих сайта – внеклеточные гидрофильные α-субъединицы (15 сайтов N-гликозилирования и 37 цистеиновых остатков) и две трансмембранные β-субъединицы, связанные друг с другом дисульфидными связями. β-субъединица инсулинового рецептора обладает тирозинкиназной активностью (49).
Началу активного изучения ИР в мозге в первой половине 1970-х годов предшествовало исследование на крысах, в котором было показано снижение уровня глюкозы в крови в ответ на введение 500 μU инсулина в сонную артерию (50). Кроме того, в других исследованиях на животных было показано активное связывание меченого 125I-инсулина на мембранах клеток различных тканей (51).
Метаболизм углеводов в печени регулируется преимущественно посредством холинергической активации, нежели изменением секреторной активности островковых клеток поджелудочной железы. Впервые ИР были обнаружены в структурах мозга в 1978 году (52) и присутствовали на всех исследуемых этапах развития (53). С этих пор широкое распространение ИР в ЦНС было неоднократно подтверждено in vitro и in vivo.
Использование радиоактивной метки для оценки содержания инсулина в культурах тканей показало преобладание ИР на поверхности клеток передней части гипоталамуса и значительно более низкое количество ИР на мембранах клеток задней доли гипоталамуса, таламуса и коры больших полушарий (54). Активное связывание меченого инсулина [125I] также было показано в структурах обонятельной и лимбической систем в новой коре и корковых областях, получающих афферентные сигналы от базальных ганглиев, гиппокампе, мозжечке и сосудистом сплетении мозга, что подтверждает нейромодуляторную роль инсулина в ЦНС (55). Методами авторадиографии и компьютерной денситометрии показано преобладание ИР в зонах, связанных с обонянием, пищевым поведением и автономными функциями (56). Данные гибридизации in situ демонстрируют высокий уровень мРНК ИР в клетках гранулярного слоя обонятельной луковицы, мозжечка и зубчатой извилины, а также пирамидных клетках пириформной коры, гиппокампа, сосудистого сплетения и дугообразного ядра гипоталамуса (57).
Экспрессия мРНК ИР выше у крыс Цукера с ожирением (fa/fa) по сравнению с крысами с нормальной массой тела (Fa/−) (58). Однако связывание инсулина в культуре клеток мозга нормальных крыс не отличается от связывания гормона у крыс с стрептозоцин-индуцированным диабетом, что опровергает данные о повышении экспрессии рецепторов инсулина при СД (59).
Наряду с IR, рецепторы инсулиноподобного фактора роста (ИФР1-Р) также широко распространены в ЦНС, в частности в обонятельных и сенсорных зонах мозга и областях, отвечающих за контроль автономной регуляции, включая гипофиз (60). Кроме того, характер экспрессии ИР и ИФР1-Р в мозге крыс обладает межполушарной асимметрией и различается у самцов и самок. Предполагается, что различия в пространственном распределении этих рецепторов могут лежать в основе патогенеза некоторых психических расстройств и поведенческих различий, в частности связанных с активностью гиппокампа (например, пространственное обучение и адаптивный ответ на стресс), между мужчинами и женщинами (61).
Присутствие ИР в гипоталамусе, коре больших полушарий и мозжечке в отсутствии СД подтверждено в исследованиях post-mortem (62). Динамика связывания меченого инсулина на мембранах синаптосом коры больших полушарий меняется в процессе развития. Активное связывание инсулина с ИР в мозге обнаруживается уже на 14-й неделе внутриутробного развития, постепенно снижается к 30-й неделе и достигает минимальных значений после рождения (63).
Несмотря на различия в размере (α-субъединица ИР в мозге, ИР-А, меньше по молекулярному весу, чем α-субъединица периферического ИР, ИР-В), степени гликозилирования (выше в периферических ИР) и специфичности, кинетика и фармакологические свойства ИР в ЦНС не отличаются от свойств инсулиновых рецепторов в периферических тканях (64). С другой стороны, связывание инсулина центральными и периферическими ИР запускает разные молекулярные каскады. Так, избыток инсулина в периферических тканях вызывает снижение экспрессии ИР, и не влияет на экспрессию ИР в мозге (65). Такая гетерогенность действия центральных и периферических ИР способствует независимой и специфичной регуляции клеточных каскадов в ответ на связывание одних и тех же лигандов.
Активность экспрессии ИР также определяется типом рецептора (66). Плотность ИР на мембранах нейронов меняется на каждом этапе развития мозга. В процессе активного нейрогенеза наиболее высокий уровень ИР обнаружен в таламусе, хвостатом ядре и скорлупе, а также некоторых ядрах среднего мозга и ствола. В мозге взрослых особей плотность ИР в этих структурах значительно снижается (67).
Система инсулиновой сигнализации в мозге
Инсулиновый рецептор принадлежит семейству рецепторных тирозинкиназ. Связывание инсулина с α-субъединицей ИР в нейронах и клетках глии стимулирует фосфорилирование и активацию β-субъединицы, обладающей тирозинкиназной активностью (68). У млекопитающих механизм инсулиновой сигнализации (Рисунок 2) регулируется посредством фосфорилирования тирозиновых остатков в составе некоторых белков, включая субстраты ИР (ИРС) (69) и адапторные белки (70), которые способствуют интеграции действия рецепторных тирозинкиназ и других звеньев сигнального пути (71). Связывание инсулина вызывает интернализацию ИР в составе окаймленных везикул (72). Этот процесс играет основополагающую роль в инсулиновой сигнализации. Попадая внутрь клетки, ИР деградируют под действием внутриклеточных ферментных комплексов или снова встраиваются в мембрану.
.png)
Рисунок 2. Трансдукция сигнала и реализация физиологических эффектов инсулина и ИФР-1
Действие инсулина в ЦНС реализуется преимущественно за счет ИРС-1 и ИРС-2. ИРС-1 участвует в регуляции роста и опосредует периферическое действия инсулина, в то время как ИРС-2 играет важную роль в созревании нейронных структур, контроле массы тела, гомеостазе глюкозы и репродуктивной функции самок (74). На NH2-терминальном конце ИРС белков содержится домен гомологии плекстрина (РН), за которым следует фосфотирозин-связывающий (PTB) домен. На С-терминальном конце находятся сайты фосфорилирования тирозина и серина/треонина (75).
Сайты фосфорилирования тирозина участвуют в координации сигнальных каскадов посредством связывания SH2 доменов эффекторных (фосфоинозитид-3-киназа, PI3K; фосфатаза SHP2; тирозинкиназа Fyn) или адапторных белков (SOCS1, SOCS-3, GRB2 и др.) (70, 74). Фосфорилирование серина ИРС-1/2 с-Jun N-терминальной киназой (JNK1) и другими протеинкиназами, наоборот, подавляет фосфорилирование тирозина в ответ на связывание инсулина. Это, наряду с убиквитин-опосредованной деградацией ИРС-1/2, играет важную роль в развитии инсулинорезистентности (76, 77). С другой стороны, повышение экспрессии ИРС-2 в ответ на действие цАМФ и активации CREB способствуют усилению инсулиновой сигнализации (78).
Синапс образует физический контакт между нейронами и способствует передаче сигналов между клетками. На роль ИРС сигнализации в постнаптическом аппарате нейронов указывают данные о коэкспрессии ИР и тирозинкиназного субстрата ИР p58/53 (IRSp53) в богатых синапсами молекулярном и гранулярном слоях мозжечка, а также в синапсах изолированных клеток гиппокампа (79). Фосфорилирование IRSp53 в ответ на действе инсулина (80, 81) играет важную роль в реорганизации цитоскелета, предшествующей росту нейритов (82), а также участвует в патогенезе ряда нейродегенеративных заболеваний (83). Это подтверждается результатами исследований на животных, демонстрирующих, что нокаут гена IRSp53 вызывает нарушения когнитивных функций, выявляемых в лабиринте Морриса и тесте на распознавание новых объектов (84).
Взаимодействие ИРС и PI3K приводит к активации PI3K и последующему фосфорилированию компонента плазматической мембраны инозитола PI (4,5)P2 с образованием инозитолтрифосфата PI (3,4,5)P3. Это, в свою очередь, стимулирует ассоциацию серинтреониновой PDK киназы (3-фосфоинозитол-зависимая протеинкиназа) и протеинкиназы В (PKB или Akt) с плазматической мембраной клетки, а также активацию Akt в ответ на фосфорилирование киназами PDK1 и PDK2 (85). Этот сигнальный путь подавляется действием липидной фосфатазы PTEN или SHIP2. Фосфорилирование TSC2 (комплекс туберозного склероза 2, туберин 2) киназой Akt приводит к активации мишени рапамицина млекопитающих (mTOR) – цитоплазматической протеинкиназы, играющей ключевую роль в регуляции клеточной пролиферации и метаболизма, и таким образом опосредует связь между инсулиновой сигнализацией и чувствительностью к уровню нутриентов (70, 86). Помимо IRS/PI3K/Akt сигнального пути существует периферический путь инсулиновой сигнализации, способствующий транслокации транспортера глюкозы GLUT-4 в ответ на действие инсулина и включающий ряд молекулярных субстратов ИР – Cbl и APS. Интеграция различных белков в состав липидных рафтов индуцирует слияние везикул, содержащих GLUT-4, с плазматической мембраной клетки (71, 85).
Связывание инсулина с ИР также приводит к фосфорилированию остатков тирозина в адапторных белках Gab-1/Shp2, Shc/Grb2 и SOS/Grb2, активации G-белка Ras и запуску сигнального каскада митоген-активируемой протеинкиназы (МАР), в частности MAPK/ERK киназы (MEK) и киназы, регулируемой внеклеточными сигналами (88). МАР-киназа ERK (extracellular signal-regulated kinase) активирует ряд цитоплазматических белков, включая S6 рибосомальную киназу p90rsk (89), белки цитоскелета, фосфолипазу А2 (PLA2), а также сигнальные белки – рецепторные тирозинкиназы, рецепторы эстрогена и белки семейства SOS и STAT (сигнальный белок и активатор транскрипции). Попадая в ядро, ERK фосфорилирует факторы транскрипции – белки семейства Ets – и участвует в контроле экспрессии генов (18, 70).
Нарушение экспрессии и функциональной активности ИР в процессе развития, в том числе в результате точечных мутаций F382V (нарушение транспорта компонентов ИР к поверхности клетки), R735S (инсулинорезистентность в результате ингибирования синтеза белков-предшественников), L1018A (подавление тирозинкиназной активности рецептора) и Y960F (различные функциональные нарушения) лежит в основе патогенеза некоторых функциональных нарушений мозга (49).
Действие инсулина в ЦНС
Роль инсулина в регуляции энергетического обмена, гомеостаза глюкозы и пищевого поведения
Глюкоза – основной источник энергии для клеток мозга вне условий голодания, когда источником служат кетоновые тела (90). Наряду с метаболической, глюкоза выполняет сигнальную функцию и участвует в регуляции углеводного гомеостаза в ЦНС и периферических тканях.
Направленность действия глюкозы в ЦНС определяется присутствием двух типов рецепторов на поверхности чувствительных нейронов, играющих важную роль в регуляции пищевого поведения, энергетического обмена и углеводного гомеостаза (49). Связывание глюкозы рецепторами первого типа (glucose-excited, GE) вызывает активацию клеток, в то время как действие глюкозы на рецепторы второго типа (glucose-inhibited, GI) подавляет их активность. Сенсором глюкозы в чувствительных нейронах служит глюкокиназа, которая участвует в поддержании энергетического баланса и контроле потребления пищи (91–94).
Эффективность захвата глюкозы определяется уровнем экспрессии белков транспортеров (Таблица 1) и сенсоров глюкозы (95). Транспортный белок GLUT-1 наиболее широко представлен в ЦНС. Две изоформы белка-транспортера различаются степенью гликозилирования. В астроцитах экспрессируется изоформа с молекулярным весом 45 кДа, устойчивая и гипо- и гипергликемии; экспрессия изоформы с молекулярным весом 55 кДа встречается преимущественно в эндотелии, повышается в ответ на гипогликемию и устойчива к гипергликемии.
Функция GLUT-1 в структурах ЦНС определяется специализацией клеток, поэтому инсулин может оказывать разнонаправленное действие на активность транспортных белков (49, 96). GLUT-2, совместно с глюкокиназой и рецептором сульфонилмочевины 1 (SUR1), экспрессируется преимущественно в латеральной области гипоталамуса и некоторых его ядрах – паравентрикулярном и дугообразном (97, 98, 49, 93, 99). В нейронах мозжечка, стриатума, коры больших полушарий и гиппокампа, а также в некоторых глиальных клетках и эндотелии преобладают транспортеры глюкозы типа GLUT-3, активность которых повышается при низком уровне глюкозы.
Читайте также: Маркер для восстановления цвета ткани
Таблица 1. Основные изоформы транспортеров инсулина в мозге
Глиальные клетки и эндотелий сосудов
Присутствуют в большом количестве
Нейроны, глиальные клетки, танициты
Присутствуют в ограниченном количестве
Мозжечок, стриатум, кора и гиппокамп
Нейроны, глиальные клетки и клетки эндотелия
Присутствуют в большом количестве
Обонятельная луковица, зубчатая извилина гиппокампа, гипоталамус и мозжечок
Присутствуют в большом количестве в некоторых областях r
Глюкоза, инсулин, физическая активность
Гипоталамус, мозжечок, ствол мозга, гиппокамп, зубчатая извилина, первичная обонятельная кора
Тела нейронов и апикальные дендриты
Присутствуют в ограниченном количестве
В отличие от периферических тканей, мозг долгое время считался нечувствительным к инсулину, в том числе из-за низкого уровня транспортеров GLUT-4 в структурах ЦНС. Наиболее высокий уровень GLUT-4 обнаружен на мембране и в цитоплазматическом пуле клеток обонятельной луковицы, зубчатой извилины гиппокампа, гипоталамуса и коры больших полушарий, хотя в этих структурах он не превышает содержание GLUT-1 и GLUT-3 (102). Наиболее высокий уровень GLUT-4 обнаружен в мозжечке, где его экспрессия напрямую регулируется действием инсулина (103). В ответ на внутривенное введение глюкозы и увеличение уровня инсулина повышается встраивание цитоплазматических форм GLUT-4 в мембрану клеток мозжечка, коры и гиппокампа (104). В GE и GI нейронах гипоталамуса GLUT-4 коэкспрессируется с ИР и глюкокиназой, что способствует повышению захвата глюкозы в ответ на действие инсулина (105).
Однако инсулин-зависимая регуляция не является доминирующим механизмом контроля транспорта глюкозы. Это подтверждается данными о наличии нейронального ответа на изменение уровня глюкозы даже в отсутствии инулина (97, 98, 106). Кроме того, показано, что инсулин не влияет на захват глюкозы в гиппокампе, а связывание инсулина с ИР не вызывает увеличения AS160-зависимой транслокации GLUT-4 (104).
Транспортер GLUT-8 обнаруживается только в структурах ЦНС, в частности в возбуждающих и тормозящих нейронах гиппокампа, и при нормальных физиологических условиях и в экспериментальной модели СД1 (107) экспрессируется только на мембране тел нейронов и проксимальных участках апикальных дендритов (108, 109). Хотя физиологическая роль GLUT-8 до сих пор малоизучена, чувствительность GLUT-8 к инсулину указывает на роль этого транспортера в мобилизации субстрата в условиях дефицита глюкозы (110).
Внутри клетки GLUT-8 способствует транспорту глюкозы из шероховатого эндоплазматического ретикулума (ЭР) в цитоплазму и поддержанию гомеостаза глюкозы в клетках гиппокампа, обладающих высокой чувствительностью к гипергликемии и снижению уровня инсулина (111, 112). Введение глюкозы вызывает активацию GLUT-8 и увеличение транспорта глюкозы из цитоплазмы в шероховатый ЭР, но не стимулирует встраивание белка в мембрану.
Подавление действия инсулина в гипоталамусе или связывание инсулина с нейронами дугообразного ядра снижает регуляторное влияние инсулина на синтез глюкозы в печени (113) и может вызывать нарушение контроля глюконеогенеза (114) и гипергликемию у пациентов с СД (115, 116).
Действие инсулина на активность нейронов гипоталамуса опосредовано АТФ-чувствительными K+-каналами (117), активация которых вызывает гиперполяризацию и снижение ответа на глюкозу (118). Резекция отростков блуждающего нерва, через которые клетки печени получают афферентацию от гипоталамуса, также подавляет ингибирующее действие инсулина на образование глюкозы в гепатоцитах (116). Эти данные подтверждают результаты работы Claude Bernard (119), который в 1855 году показал, что разрушение четвертого желудочка вызывает глюкозурию у мышей.
Популяции нейронов, участвующих в поддержании энергетического гомеостаза, расположены преимущественно в гипоталамических центрах голода и насыщения. Как и GLUT-2, глюкокиназа (92–94), AMPK и PASK и рецепторы орексигенных и анорексигенных молекул, которые продуцируются клетками этих ядер, выступают в качестве сенсоров энергетического обмена, генерируя интегрированный ответ на афферентную стимуляцию при изменениях энергетического гомеостаза или дефиците ресурсов. Гипоталамические центры голода и насыщения детектируют сигналы об уровне глюкозы и передают их в другие зоны мозга, что приводит к активации пищевого поведения. Активация PI3K каскада является общим звеном сигнального каскада инсулина, лептина и серотонина, и играет важную роль в патогенезе инсулинорезистентности и контроле потребления пищи. В периферических тканях инсулин стимулирует анаболические процессы, в то время как в структурах ЦНС он проявляет анорексигенные свойства и выполняет катаболическую роль (120). Кроме того, в гипоталамусе инсулин активирует JAK2 и SHT3 и усиливает действие лептина (121). Механизмы резистентности нейронов гипоталамуса к инсулину и лептину у мышей с СД2 открывают новые возможности в изучении механизмов инсулинорезистентности при СД2 (33).
Роль инсулина в регуляции полового поведения
Репродуктивная способность во многом зависит от энергетического метаболизма, и нарушение гомеостаза глюкозы может нарушать нормальную регуляцию полового поведения (122), изменяя активность гипоталамо-гипофизарно-гонадальной оси (123). Достаточный уровень энергетических ресурсов способствует нормальному функционированию репродуктивной системы и повышает выживаемость потомства.
Воздействие низких концентраций инсулина на клетки гипоталамуса стимулирует секрецию рилизинг-фактора лютеинизирующего гормона (ЛГ-РГ) и зависит от гомеостаза глюкозы (124), что подтверждает его роль в регуляции репродуктивной функции. С другой стороны, воздействие инсулина в высокой концентрации не влияет на секрецию ЛГ-РГ (124). Введение инсулина в культуру клеток коры больших полушарий также повышает частоту импульсной секреции лютеинизирующего гормона (ЛГ), в то время как при введении глюкозы такой эффект не наблюдается (125). У мышей с СД низкий уровень инсулина в ЦНС вызывает снижение высвобождения ЛГ (124), которое восстанавливается при введении инсулина в мозг или периферические ткани (126). В других исследованиях показано, что низкий уровень инсулина у крыс с СД способствует снижению секреции гонадотропин-рилизинг гормона (ГнРГ) в гипоталамусе и подавлению ответа гонадотропных клеток гипофиза на действие ГнРГ (126). Таким образом, инсулин играет важную роль в контроле импульсной секреции ГнРГ (127). Однако авторы отмечают, что секреция ЛГ зависит не только от действия инсулина, но и от содержания глюкозы. Это опосредовано детекторами глюкозы в гипоталамусе, которые также могут влиять на секрецию ГнРГ, независимо от уровня инсулина (128).
Влияние инсулина на клеточную пролиферацию и дифференцировку
Трофическая функция инсулина в ЦНС реализуется через регуляцию клеточной пролиферации, дифференцировки и роста нейритов в процессе эмбрионального развития. Внутривенное введение инсулина повышает активность орнитин декарбоксилазы в мозге новорожденных крыс, которая служит маркером нейронального развития (129). Участие инсулина в процессах пролиферации подтверждается данными о повышении количества ИР в период активной клеточной дифференцировки (130). Действие ИР на созревание (131) мозга, и также рост и регенерацию аксонов (132) реализуется посредством ИРС-2.
Нейротрофическая функция инсулина была продемонстрирована в исследованиях in vitro. В мозге крыс инсулин стимулирует пролиферацию (133), а в культуре нейронов переднего мозга курицы активирует рост и дифференцировку нейронов (134). ИР, участвующие в реализации эффекта инсулина на нейрональный рост и развитие, также найдены на мембранах глиальных клеток (135, 136), в которых направленность их действия определяется типом клеток. Кроме того, инсулин и ИФР2 стимулируют активность фактора роста нервов (ФРН), необходимого для роста нейритов (137). Действие инсулина на развитие ЦНС зависит от присутствия астроцитов (138), в которых наблюдается активная пролиферация белков сигнального каскада инсулина (139, 140).
В культуре эмбриональных клеток инсулин повышает фосфорилирование рибосомального белка S6 (136) и активность протеинкиназы-эпсилон в цитоплазме клетки (141), стимулируя рост нейритов (142, 143).
Механизм действия инсулина на нейрональное развитие также включает другие ферменты, в том числе фосфатидилинозитол-3-киназу (PI3K) (144). Активация сигнального пути PI3K/mTOR в ответ на действие инсулина повышает экспрессию белка постсинаптической плотности PSD-95 в нейронах CA1 и способствует формированию дендритных шипиков и возбуждающих синапсов в клетках гиппокампа (145, 146).
В инсулин-зависимой регуляции роста нейритов также участвует тау-белок, ассоциированный с микротрубочками. Эффект тау-белка на аксональный рост реализуется через PI3K/mTOR каскад и сопровождается повышением экспрессии мРНК и уровня тубулина (147).
Эндогенный инсулин способствует формированию нейрофиламентов (148) и участвует в регуляции пролиферации и дифференцировки плюрипотентных стволовых клеток. Недостаточность инсулина вызывает неапоптотическую клеточную гибель (130), а снижение активности PI3K/Akt каскада нарушает процесс дифференцировки стволовых клеток человека (hNSC), которые, в отличие от стволовых клеток грызунов, обладают высокой чувствительностью к инсулину и функционируют в узком диапазоне концентрации гормона (149). Таким образом, инсулин играет ключевую роль в интеграции нейрональной активности и регуляции энергетического гомеостаза, определяющих каждый этап клеточной дифференцировки (150).
Нейропротективный эффект инсулина
Нейропротективный эффект инсулина проявляется в подавлении апоптоза, инактивации β-амилоидного белка, снижении оксидативного центра и ишемии. Важную роль в реализации антиапоптотического эффекта инсулина играет активность PI3K/Akt/mTOR пути и белка p70SK. Ингибирование mTOR рампамицином снижает антиапоптотический эффект инсулина, что указывает на важную роль PI3K/Akt/mTOR пути и p70SK белка в реализации этого эффекта (151). Кроме того, инсулин снижает образование Aβ фибрилл, предотвращая клеточную гибель в результате накопления β-амилоида (152).
Образование активных форм кислорода (АФК) в результате оксидативного стресса индуцирует окисление липидов и белков, вызывая нарушения функциональной активности GLUT-3 и захвата глюкозы. Это приводит к накоплению лактата, ацидозу и митохондриальной дисфункции (153). В условиях оксидативного стресса инсулин стимулирует поглощение глюкозы и образование пирувата, способствуя восстановлению внутриклеточной концентрации АТФ (154). Кроме того, инсулин предотвращает внесинаптическое накопление глутамата и гама-аминомасляной кислоты (ГАМК) в результате снижения захвата этих нейромедиаторов при оксидативном стрессе (155) и стимулирует накопление мочевой кислоты, обладающей антиоксидантной активностью (156).
Антиишемическое действие инсулина реализуется с помощью двух основных механизмов – прямого действия на нейрональные рецепторы и повышения уровня глюкозы в периферических тканях (157). При транзиторной ишемии инсулин повышает уровень ГАМК – главного тормозного нейромедиатора в мозге – во внеклеточном пространстве (158).
Стимуляция Na+/K+ АТФазы в ответ на действие инсулина приводит к снижению внеклеточной концентрации K+ и внутриклеточной концентрации Na+, способствуя уменьшению нейрональной активности и метаболических потребностей клеток, предотвращению накопления жидкости и образованию постишемического отека тканей, а также снижению уровня лактата (159, 160).
С другой стороны, реперфузия при ишемии мозга стимулирует фосфорилирование JNK1/2, экспрессию Bcl-2 и деградацию каспазы-3 в гиппокампе крыс, что указывает на важность интегрированной активности Akt и JNK1/2 в реализации противоишемического действия инсулина.
Нейромодуляторный эффект инсулина
Нейромодуляторное действие инсулина реализуется через регуляцию активности ионных токов и контроль уровня и действия нейромедиаторов. Электрофизиологический эффект инсулина in vivo опосредован действием ГАМК, которое снижается в ответ на введение ингибиторов ИР (161). Кроме того, рецепторы ГАМК являются одним из основных субстратов Akt фосфорилирования, что подтверждает роль инсулина в регуляции плотности рецепторов ГАМК на постсинаптической мембране (162).
Изменение активности ионных каналов в ответ на действие инсулина способствует реорганизации ионных токов и изменению активности нейронов. В нейронах гипоталамуса инсулин повышает активацию K+АТФ каналов, что приводит к гиперполяризации мембраны и торможению нейрональной активности (163). Кроме того, инсулин стимулирует Na+/K+ АТФазу и вызывает увеличение внутриклеточной концентрации Ca2+, стимулируя высвобождение нейропептидов (164).
Инсулин способствует подавлению высвобождения норадреналина и повышению захвата серотонина в нейронах (165, 166), и, как следствие, увеличению эффективности метаболизма глюкозы (165, 166). В нейронах стриатума крыс с стрептозоцин-индуцированным СД (167) и при введении галоперидола (168), инсулин увеличивает плотность дофаминовых рецепторов, хотя у здоровых животных этот эффект не наблюдается. Системное введение инсулина вызывает повышение уровня дофамина и серотонина в ЦСЖ и снижение плотности
α2-адренергических рецепторов в нейронах гипоталамуса (169). Помимо этого, инсулин стимулирует захват аминокислот, необходимых для синтеза нейромедиаторов (170).
Влияние инсулина на когнитивные функции и память
Внутривенное введение, внутрицеребровентрикулярная или внутригиппокампальная инфузия инсулина связаны с улучшением когнитивных функций и памяти (171), которые сопровождаются увеличением экспрессии ИР и активацией сигнальных каскадов в гиппокампе (172). Инсулин также способствует восстановлению памяти при ишемическом поражении мозга (173).
Введение низких доз стрептозоцина в мозг вызывает центральную инсулинорезистентность подавляет активность инсулина и приводит к нарушениям памяти и поведения (174). СД1 и СД2 повышают риск когнитивных нарушений и развития БА у пожилых пациентов. В эпидемиологических исследования показано, что введение инсулина больным с СД1/СД2 и БА улучшает формирование памяти и способствует поддержанию уровня глюкозы (175). Системное введение инсулина здоровым испытуемым в условиях эугликемии и гиперинсулинемии улучшает показатели вербальной памяти и селективного внимания.
Ключевую роль в формировании кратковременной и долговременной памяти играют процессы долговременной потенциации (ДП) и долговременной депрессии (ДД) (176). ДП возникает в ответ на длительную и синхронную активацию пресинаптических и постсинаптических нейронов, способствующую поддержанию деполяризации постсинаптической мембраны в результате повышения уровня Ca2+ и консолидации. ДД является компенсаторным механизмом и вызывает снижение эффективности нейрональной передачи в ответ на интенсивное воздействие стимула. Наряду с ДП и ДД, важную роль в процессах обучения и памяти играет пластичность дендритных шипиков и реорганизация цитоскелета, опосредованные действием глутамата на AMPA и NMDA рецепторы. Активность рецепторов регулируется изменением их плотности на клеточной мембране или ковалентной модификацией белковых субъединиц. В процессе ДП наблюдается увеличение плотности AMPA рецепторов на постсинаптической мембране, в то время как ДД приводит к снижению количества рецепторов. Фосфорилирование и дефосфорилирование глутаматных рецепторов при ДП и ДД играет важную роль в контроле эффективности нейронной передачи (177).
Инсулин вызывает снижение плотности AMPA рецепторов на постсинаптической мембране, способствуя ДД. Этот процесс во многом зависит от уровня фосфорилирования ИР, активности PI3К и синтеза белка (178). Кроме того, инсулин стимулирует фосфорилирование субъединицы GluR2 AMPA рецепторов в нейронах гиппокампа, способствуя их интернализации и снижению возбудимости клеток (179).
Важную роль в реализации действия инсулина на процессы обучения и памяти играет ГАМК. Инсулин стимулирует транслокацию и экспрессию рецепторов ГАМК на постсинаптической мембране, в то время как подавление активности PI3K снижает этот эффект (180).
Стимулирующее действие ИФР1 на активность нейронов в гиппокампе крыс реализуется с участием AMPA рецепторов и PI3К (181). О роли соматотропного гормона (СГ) в ДП и формировании памяти свидетельствует повышение экспрессии NMDA рецепторов в гиппокампе (182).
Воспаление и инсулинорезистентность
У пациентов с ожирением и СД2 воспаление является ключевым звеном патогенеза инсулинорезистентности в ЦНС и периферических тканях и повышает риск развития когнитивных нарушений и БА (183). У пациентов с БА обнаружено увеличение уровня интерлейкина IL-6 в ЦСЖ (184), что указывает на роль воспаления в аккумуляции β-амилоидного пептида Aβ (185). Результаты экспериментальных исследований демонстрируют противовоспалительное действие инсулина у пациентов с БА (186).
Введение липополисахарида приводит к увеличению уровня С-реактивного белка и провоспалительных цитокинов IL-1β, IL-6 и TNF в плазме крови (187). TNFα и IL-6, в свою очередь, стимулируют активацию NFkβ и транскрипцию провоспалительных факторов TNFα, IL-6 и IL-1b (188). Это приводит к нарушению синаптической пластичности в гиппокампе и формированию пространственной памяти (189).
Инсулинорезистентность периферических тканей у пациентов с ожирением связана с увеличением уровня провоспалительных цитокинов и свободных жирных кислот. Хроническое воспаление также способствует развитию инсулинорезистености, СД2 и БА (190). В частности, повышение уровня TNFα и Aβ в мозге пациентов с гиперинсулинемией стимулирует образование амилоидных бляшек (191). Среди пациентов с ожирением и БА наблюдается снижение концентрации инсулина в мозге, что указывает на нарушение транспорта инсулина через ГЭБ и снижение чувствительности нейронов к действию гормона (192).
Читайте также: Черная кремнеземная негорючая ткань
В зависимости от типа рецепторов – TNF-R1 или TNF-R2 – TNF-α проявляет нейротоксический или нейропротективный эффект. TNF-R1 оказывает проапоптотическое действие, в то время как TNF-R2 предотвращает клеточную гибель. У пациентов с БА, диабетом и нарушением толерантности к глюкозе наблюдается увеличение уровня TNF-R1 и снижение уровня TNF-R2 (193, 194), который нормализуется после трех недель высококалорийной диеты (195). Накопление конечных продуктов гликирования, оксидативный стресс и повреждение клеточных структур способствуют нарушению когнитивных функций у пациентов с СД (196).
Нарушение каскадов инсулиновой сигнализации, в частности PI3K/Akt и киназы-3 гликогенсинтетазы GSK-3, связано с повышением воспаления и инсулинорезистентностью (197). Известно, что PI3K подавляет образование IL-12 в дендритных клетках, а GSK-3 способствует гиперфосфорилированию и регуляции метаболизма Aβ (198).
Активация ИР приводит к увеличению фосфорилирования и подавлению активности GSK-3β (199). У пациентов с БА и СД2 наблюдается повышение активности GSK-3β и увеличение фосфорилирования ИР и ИРС-1 (200). Кроме того, сигнальный каскад PI3K/Akt/GSK-3 играет важную роль в активации STAT-3 в глиальных клетках (201): ингибирование GSK-3 вызывает повышение уровня противовоспалительных цитокинов, в частности IL-10, и снижение образования провоспалительных факторов – IL-1β, IL-6 и IFN-γ (202).
Известно, что воспаление играет ключевую роль в патогенезе БА, способствуя повышению продукции провоспалительных факторов Т-лимфоцитами, моноцитами и клетками глии (203, 204). Данные исследований на животных показывают, что подавление продукции провоспалительных цитокинов позволяет снизить выраженность симптомов инсулинорезистентности (205). Хотя воспаление не участвует в развитии острой инсулинорезистентности, которая развивается в ответ на рацион с высоким содержанием жира (206), оно оказывает значительное влияние на развитие хронической инсулинорезистентности (207).
Роль TNF-α в патогенезе инсулинорезистентности реализуется через подавление тирозинкиназной активности ИР и снижение активности SOCS-3, приводя к инактивации ИРС-1 (208). Кроме того, активация JNK в ответ на повышение уровня факторов воспаления и свободных жирных кислот повышает фосфорилирование ИРС-1 и снижает эффективность инсулиновой сигнализации, способствуя увеличению уровня воспаления и развитию инсулинорезистентности. Как показывают исследования на животных, инактивация JNK и подавление воспалительного ответа позволяют избежать негативных метаболических эффектов, вызванных гиперлипидемией (210, 211, 212).
Внутрирецебровентрикулярная инфузия TNF-α стимулирует воспаление в гипоталамусе, повышение уровня инсулина и нарушение инсулиновой сигнализации в периферических тканях. Считается, что центральная инсулинорезистентность служит адаптивным механизмом при высококалорийной диете и сопровождается нарушением гомеостаза глюкозы. Повышение уровня свободных жирных кислот служит сигналом для активации высвобождения провоспалительных цитокинов, подавляющих действие инсулина (213).
У пациентов с БА показано нарушение инсулиновой сигнализации, снижение тирозинкиназной активности ИР, подавление экспрессии мРНК инсулина и ИФР1 и их рецепторов (214). Это вызывает снижение эффективности сигнальных каскадов инсулина (IR/IRS-1/PI3K) и ИРФ1 (IGF1R/IRS-2/PI3K). Уменьшение ответа на действие инсулина опосредовано фосфорилированием остатков серина (IRS-1 pS616, IRS-1 pS636/639) и инактивацией ИРС-1. При умеренных и выраженных когнитивных нарушениях наблюдается повышение маркеров инсулинорезистентности, независимо от уровня аполипопротеина E (APOE-4) (104). Снижение чувствительности к инсулину также сопровождается повышенным накоплением Aβ, который, в свою очередь, оказывает тормозящее действие на экспрессию и функциональную активность гормона (215). Пептид Aβ проявляет антагонистическое действие по отношению к инсулину, конкурентно связываясь и снижая аутофосфорилирование ИР (216, 217). Это может препятствовать реализации нейропротективного эффекта инсулина у пациентов с БА и СД2 и повышать риск когнитивных нарушений (218). Резистентность к инсулину и ИФР1 служит ранним биомаркером БА и указывает на дисфункцию ИРС-I, вызванную действием Aβ (104).
Сахарный диабет и болезнь Альцгеймера
Результаты последних исследований указывают на ключевую роль инсулина в регуляции нейрональной активности и тесную связь между БА и СД2 (219) (Рисунок 3). БА – самое распространенное нейродегенеративное заболевание, которое сопровождается потерей памяти и прогрессирующей деменцией. В настоящее время более 30 миллионов людей страдают БА, а к 2040 году ожидается рост встречаемости заболевания до 120 миллионов (197).
Важным звеном патогенеза БА является накопление амилоидных бляшек и нейрофибриллярных кубков – агрегатов гиперфосфорилированного тау-белка, а также церебральная амилоидная ангиопатия и нейродегенерация (220).
СД2 сопровождается нарушением секреции инсулина и инсулинорезистентностью. В 2010 году в мире насчитывалось 250 миллионов пациентов с сахарным диабетом, 90% из которых страдали СД2 (221). Наряду с ожирением, старение является одним из ключевых факторов риска БА и СД2.
.png)
Рисунок 3. Нарушение инсулиновой сигнализации в патогенезе болезни Альцгеймера. Aβ, β-амилоидный пептид; GLUT-3, транспортер глюкозы 3; GSK-3β, киназа-3 β гликогенсинтетазы; NFT, нейрофибриллярные клубки; PI3K, фосфатидилинозитол-3-киназа.
Инсулинорезистентность, воспаление, накопление амилоидного пептида и когнитивные нарушения являются общими звеньями патогенеза БА и СД2. Наряду с периферической инсулинорезистентностью, в патогенезе этих заболеваний также участвует резистентность к ИФР1 и нарушение функциональной активности ИРС-1 и ИРС-2.
Инсулинорезистентность в периферических тканях предшествует манифестации СД2, что представляет собой важное терапевтическое окно для своевременного лечения (222). Особый интерес вызывает вопрос о роли периферической и центральной инсулинорезистентности в патогенезе БА и СД2. Отсутствие СД2 у пациентов с БА может быть связано с поздней манифестацией симптомов диабета, хотя проявление симптомов когнитивной дисфункции также обнаруживается на поздних стадиях болезни.
Выделают четыре стадии прогрессирования БА. На первой стадии болезнь не сопровождается выраженными симптомами; на преклинической стадии заболевания обнаруживаются патофизиологические изменения на фоне отсутствия выраженных когнитивных нарушений; третья стадия предшествует развитию деменции и сопровождается умеренными когнитивными нарушениями; последняя стадия БА характеризуется тяжелой когнитивной дисфункцией и деменцией (223, 224).
Изменение уровня инсулина и нарушение метаболизма глюкозы повышают риск развития деменции (225, 226). Результаты последних исследований демонстрируют повышение экспрессии маркеров СД2 у пациентов с БА (227) и подтверждают связь между нарушением инсулиновой сигнализации и снижением когнитивных функций. В настоящее время центральная инсулинорезистентность, сопровождающаяся когнитивными нарушениями, рассматривается как отдельная форма СД – сахарный диабет 3 типа (СД3).
Однако эта форма диабета не может считаться классическим проявлением заболевания, так как у пациентов с БА не наблюдается гипергликемии, характерной для СД1 и СД2. Кроме того, в отличие от мышечной и жировой ткани и печени, инсулин не оказывает стимулирующего влияния на захват глюкозы клетками мозга (228). Центральная инсулинорезистентность может развиваться в ответ на изменение гомеостаза глюкозы в периферических тканях и сопровождаться повышением уровня Aβ и транспорта глюкозы (230). Таким образом, данные указывают на центральную роль инсулинорезистентности в манифестации БА и СД2, которой предшествует снижение чувствительности к инсулину в периферических тканях.
Переход от преддиабета к диабету длится от 10 до 15 лет и наиболее широко представлен среди пожилых людей, которые находятся в группе повышенного риска развития БА.
Преддиабет и диабет обнаруживается более чем у 81% пациентов с БА (226). Долгий период развития диабета из преддиабета объясняет, почему инсулинорезистентность у пациентов с БА не всегда сопровождается симптомами СД.
Результаты экспериментальных и эпидемиологических исследований подтверждают связь между инсулинорезистентностью при СД и риском деменции (231). Гиперинсулинемия и гипергликемия, вызванные снижением чувствительности к инсулину, способствуют образованию нейритных бляшек (233). По данным эпидемиологических исследований, с возрастом повышается вероятность коморбидного течения БА и СД, независимо от наличия сердечно-сосудистых заболеваний (234).
В то время как влияние СД2 на активность ЦНС хорошо изучено, существует гораздо меньше данных о центральных эффектах СД1 (235). Рядом исследований показана роль СД1 в нарушении процессов памяти, обучения и когнитивной гибкости (235–242). В основе этого эффекта лежит снижение количества дендритов в сером веществе мозга у пациентов с СД1 (245). Инсулиновая терапия, наоборот, способствует улучшению когнитивных функций (243, 244) и восстановлению морфологической структуры клеток (246). У пациентов с СД1 и СД2 инсулин и ИФР1 предотвращают атрофию головного мозга и когнитивную дисфункцию (247).
Однако существуют данные, опровергающие связь между БА и СД (248, 249). При прогрессирующей форме БА и отсутствии аллеля APOE-4 наблюдается снижение уровня инсулина в ЦСЖ на фоне повышения содежания гормона в крови (197). Увеличение уровня инсулина в плазме крови служит маркером инсулинорезистентности, в то время как снижение уровня инсулина в ЦСЖ указывает на нарушение метаболизма и снижение транспорта инсулина через ГЭБ.
Общими звеньями патогенеза БА и СД являются митохондриальная дисфункция, оксидативный стресс, нарушение метаболизма глюкозы, и образование модифицированных форм ЛПНП (197). Инсулин предотвращает инактивацию окислительного фосфорилирования в митохондриях и обладает протективным эффектом в отношении Aβ и окислительного стресса, в то время как при СД наблюдается снижение активности системы антиоксидантной защиты и повышение чувствительности клеток к действию токсических олигомеров амилоидного белка (250).
Независимо от наличия диабета, нарушение метаболизма глюкозы наблюдаются у большинства пациентов с БА (223), преимущественно в височно-теменной, задней поясной и лобной областях коры. Введение глюкозы стимулирует действие инсулина и способствует улучшению показателей памяти (251, 252).
Недостаточность метаболизма глюкозы связана с нарушением мембранного транспорта, энергетического гомеостаза и метаболизма тиамина. У пациентов с БА наблюдается снижение экспрессии GLUT-1 и GLUT-3 (253), особенно в нейронах коры больших полушарий и зубчатой извилине гиппокампа. Согласно результатам работы Liu et al. (254), снижение метаболизма глюкозы происходит в результате подавления экспрессии транспортных белков и O-GlcN-ацетилирования, повышения фосфорилирования тау-белка и образования нейрофибриллярных бляшек.
БА сопровождается изменением функциональной активности ключевых ферментов цикла Кребса и пентозофосфатного пути: пируватдегидрогеназного комплекса, α- кетоглутаратдегидрогеназного комплекса, транскетолазы и нуклеозид-дифосфатазы (255). Эти данные подтверждают роль митохондриальной дисфункции и изменения метаболизма тиамина в нарушении гомеостаза глюкозы в мозге при БА.
Недостаточность метаболизма глюкозы служит предиктором патофизиологических изменений, ассоциированных с СД (222). Согласно гипотезе Chen и Zhong, нарушение гомеостаза глюкозы и метаболизма тиамина и инсулинорезистентность способствуют агрегации пептида Aβ и гиперфосфорилированию тау-белка, занимая центральное место в патогенезе БА. Эти патофизиологические каскады индуцируют высвобождение провоспалительных факторов, вызывают митохондриальную дисфункцию и оксидативный стресс, повышают уровень конечных продуктов гликирования, апоптоз, эксайтотоксичность и активность протеинкиназ (223).
Общим звеном патогенеза БА и СД также является нарушение метаболизма холестерина. Гиперхолестеринемия вызывает изменение активности островковых клеток поджелудочной железы и снижение секреции инсулина (256, 257). Повышение уровня ЛПНП способствует образованию Aβ и его встраиванию в клеточные мембраны (10). Связывание Aβ с липопротеинами катализирует образование оксихолестерина, обладающего выраженным нейротоксическим действием и способствующего подавлению ERK/Akt каскада (258, 259). Гипергликемия и гиперинсулинемия стимулируют O-GlcN-ацетилирование белков каскада инсулиновой сигнализации и способствуют развитию инсулинорезистентности (260, 261). В образцах лобных областей коры пациентов с СД2 и БА наблюдается снижение активности инсулин/PI3K/Akt сигнального пути и (262) и O-GlcNAc трансферазы, наряду с повышением GSK-3β кальпаин-1-зависимой регуляции, что способствует гиперфосфорилированию белков и нейродегенерации (262).
Образование амилоидных агрегатов, изменение метаболизма Aβ и гиперфосфорилирование тау-белка служат связующим звеном в патогенезе БА и СД. В клетках островков Лангерганса могут формироваться скопления амилоидного белка IAPP, который сходен по структуре с пептидом Aβ и обладает токсическим действием на клетки поджелудочной железы (263, 224). Снижение активности шаперонов, препятствующих агрегации IAPP и Aβ, способствует образованию амилоидных и нейритных бляшек в островковых клетках и нейронах у пациентов с БА и СД (226).
Протеазы играют важную роль в метаболизме и предотвращении накопления продуктов распада Aβ. В частности, металлопептидаза IDE способствует повышению активности инсулина, ИФР и глюкагона. Нарушение инсулиновой сигнализации сопровождается выраженным увеличением уровня фосфорилированного тау-белка и пептида Aβ и способствует клеточной гибели и нейродегенерации, особенно у пациентов с СД2 (264). Повышение уровня инсулина в ЦНС оказывает стимулирующее действие на функциональную активность IDE (265, 266), а также способствует интернализации олигомеров Aβ и предотвращает их связывание с мембраной нейронов.
Важную роль в патогенезе БА играет формирование нейрофибриллярных клубков, которые представляют собой агрегаты гиперфосфорилированного тау-белка (267).
Инсулинорезистентность и периферическая гиперинсулинемия при СД2 приводят к снижению транспорта инсулина через ГЭБ, а системная инсулиновая недостаточность при СД1 повышает фосфорилирование тау-белка (235, 268).
Таким образом, центральная инсулинорезистентность вызывает повреждение нейронов гиппокампа и коры больших полушарий и служит общим звеном патогенеза СД2 и БА. Наряду с нарушением инсулиновой сигнализации, развитию БА могут способствовать вирусные, бактериальные и грибковые инфекции (269, 270, 271). Однако во всех случаях БА сопровождается прогрессирующей нейродегенерацией.
Терапевтические подходы
Понимание механизмов регуляции когнитивных и метаболических функций открывает новые возможности в терапии инсулинорезистентности при СД2 и БА. Ни один из современных терапевтических подходов, направленных на известные звенья патогенеза нейродегенеративных заболеваний – агрегаты амилоидного пептида (272), гиперфосфорилирование тау-белка (273) и метаболизм тиамина, а также использование антиоксидантов и нейропротектиных факторов (274, 275, 276), не позволяет достигнуть высокой эффективности лечения.
В последнее время все большую популярность набирают мультитаргетные подходы, нацеленные на регуляцию молекулярных каскадов, опосредующих патогенетический эффект инсулинорезистентности (276). В частности, наиболее известными молекулярными агентами, способствующими повышению секреции инсулина, являются метформин, антагонисты рецептора, активируемого пролифераторами пероксисом PPARγ, а также миметики инкретина, глюкагоноподобный пептид-1 (ГПП-1) и гастрический ингибирующий полипептид. Наряду с этим показана высокая эффективность интраназального введения инсулина и ГПП-1 в терапии СД2 и при умеренных когнитивных нарушениях (224).
Метформин, представитель класса бигуанидов, способствует поддержанию нормального уровня инсулина натощак и контролирует образование глюкозы в печени. В последнее время появляется все больше данных о способности метформина проникать через ГЭБ, повышая чувствительность к инсулину в структурах ЦНС (277) и способствуя снижению риска деменции у пациентов с СД (278).
Использование агонистов PPARγ, в частности росиглитазона, позволяет добиться значимого улучшения чувствительности к инсулину (279), увеличения функциональной активности адипоцитов, повышения транспорта триглицеридов и жирных кислот из печени и мышц, а также предотвращения накопления Aβ и снижения уровня воспаления (280, 281). Это приводит к нормализации уровня инсулина и улучшению внимания и памяти на ранних этапах патогенеза БА (282). Несмотря на высокую эффективность антагонистов PPARγ, препараты этой группы повышают риск инфаркта миокарда, что ограничивает их применение в терапии инсулинорезистентности.
ГПП-1, представитель класса инкретинов, способствует повышению секреции инсулина в зависимости от уровня глюкозы. В терапии СД широко применяются ингибиторы дипептидилпептидазы (глиптины), агонисты (эксенатид и лираглутид) и миметики ГПП-1. Как и метформин, миметики ГПП-1 способны проникать через ГЭБ и стимулировать соответствующие рецепторы в мозге. Эксенатид и лираглутид оказывают защитное действие на клетки мозга, подавляют нейродегенеративные процессы и предотвращают прогрессирование АД (283, 284, 285). Кроме того, наряду со снижением уровня олигомеров Aβ и нейритных бляшек, агонисты ГПП-1 стимулируют нейрогенез и улучшают распознавание объектов и пространственную память (283, 286, 287) и эффективны в терапии деменции, независимо от наличия СД.
Интраназальное введение инсулина способствует эффективному проникновению гормона в ЦНС и улучшению состояния пациентов с умеренными когнитивными расстройствами и СД2 (240, 288–290, 291). Дополнительной мишенью в терапии инсулинорезистентности служит недостаточность инсулиновой сигнализации в результате фосфорилирования серина и треонина IRS-1 (292).
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом