
Мышцы тела должны рассматриваться с точки зрения их развития и функции, а также топографии систем и групп, в которые они складываются.
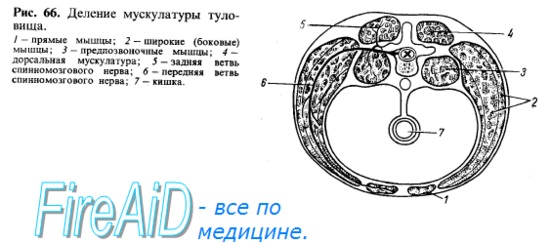
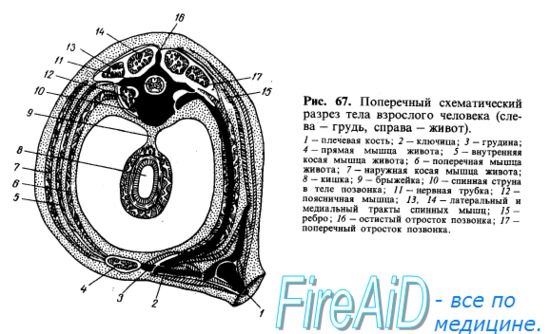
Мышцы туловища развиваются из залегающей по бокам хорды и мозговой трубки дорсальной части мезодермы, которая разделяется на первичные сегменты, или сомиты. После выделения скелеротома, идущего на образование позвоночного столба, оставшаяся дорсомедиальная часть сомита образует миотом, клетки которого (миобласты) вытягиваются в продольном направлении, сливаются друг с другом и превращаются в дальнейшем в симпласты мышечных волокон. Часть миобластов дифференцируется в особые клетки — миосателлиты, лежащие рядом с симпластами. Миотомы разрастаются в вентральном направлении и разделяются на дорсальную и вентральную части. Из дорсальной части миотомов возникает спинная (дорсальная) мускулатура туловища, а из вентральной — мускулатура, расположенная на передней и боковой сторонах туловища и называемая вентральной (рис. 66, 67).
В каждый миотом (миомер) врастают ветви соименного спинномозгового нерва (невромера). Соответственно делению миотома на 2 части от нерва отходят 2 ветви, из которых дорсальная (задняя) входит в дорсальную часть миотома, а вентральная (передняя) — в вентральную. Все происходящие из одного и того же миотома мышцы снабжаются одним и тем же спинномозговым нервом. Соседние миотомы могут срастаться между собой, но каждый из сросшихся миотомов удерживает относящийся к нему нерв. Поэтому мышцы, происходящие из нескольких миотомов (например, прямая мышца живота), иннервируются несколькими нервами. Первоначально миотомы на каждой стороне отделяются друг от друга поперечными соединительнотканными перегородками, myosepta (рис. 68). Такое сегментированное расположение мускулатуры туловища у низших животных остается на всю жизнь. У высших же позвоноч.ых и у человека благодаря более значительной дифференцировке мышечных масс сегментация значительно сглаживается, хотя следы ее и остаются как в дорсальной (короткие мышцы перекидывающиеся между позвонками), так и в вентральной мускулатуре (межреберные мышцы и прямая мышца живота). Часть мышц, развившихся на туловище, остается на месте, образуя местную, аутохтонную мускулатуру (autos — тот же самый, chthon, греч. — земля). Другая часть в процессе развития перемещается с туловища на конечности. Такие мышцы называются трункофугальными (truncus — ствол, туловище, fugo — обращаю в бегство). Наконец, третья часть мышц, возникнув на конечностях, перемещается на туловище. Это трункопетальпые мышцы (рею — стремлюсь). На основании иннервации всегда можно отличить аутохтонную (т. е. развивающуюся в данном месте) мускула гуру от сместившихся в эту область других мышц-пришельцев
Мускулатура конечностей образуется из мезенхимы почек конечностей и получает свои нервы из передних ветвей спинномозговых нервов при посредстве плечевого и пояснично-крестцового сплетений. У низших рыб (селахий) из миотомов туловища вырастают мышечные почки, которые разделяются на два слоя, расположенные с дорсальной и вентральной сторон скелета плавника. Подобным же образом у наземных позвоночных мышцы по отношению к зачатку скелета конечности первоначально располагаются дорсально и вентрально (разгибатели и сгибатели). При дальнейшей дифференцировке зачатки мышц передней конечности разрастаются и в проксимальном направлении (трункопетальные мышцы) и покрывают аутохтонную мускулатуру туловища со стороны груди и спины (mm. pectorales major et minor, m. latissimus dorsi). Кроме этой первичной мускулатуры передней конечности, к поясу верхней конечности присоединяются еще трункофугальные мышцы, т. е. производные вентральной мускулатуры. служащие для передвижения и фиксации пояса и переместившиеся на него с головы (mm. trapezius и sternocleidomastoideus) и с туловища (mm. rhomboideus, levator scapulae, serratus anterior, subclavius, omohyoideus). У пояса задней конечности вторичных мышц не развивается, так как он неподвижно связан с позвоночным столбом. Сложная дифференцировка мышц конечностей наземных позвоночных, в особенности у высших форм, объясняется функцией конечностей, превратившихся в сложные рычаги, выполняющие различного рода движения.
Мышцы головы возникают отчасти из головных сомитов, а главным образом из мезодермы жаберных дуг. Висцеральный аппарат у низших рыб состоиг из сплошного мышечного слоя (общий сжиматель), который делится по своей иннервации на отдельные участки, совпадающие с мета-мерным расположением жаберных дуг: 1-й жаберной (мандибулярной) дуге соответствует V пара черепных нервов (тройничный нерв), 2-й жаберной (гиоидной) дуге — VII пара (лицевой нерв), 3-й жаберной дуге — IX пара (языког лоточный нерв). Остальная часть общего сжимателя снабжается ветвями X пары (блуждающий нерв). Сзади общего сжимателя обособляется пучок, прикрепляющийся к поясу верхней конечности (трапециевидная мышца). Когда с переходом из воды на сушу у низших позвоночных прекратилось жаберное дыхание- приспособленное для жизни в воде, мышцы жаберного аппарата (висцеральные) распространились на череп, где превратились в жевательные и мимические мышцы, но сохранили свою связь с теми частями скелета, которые возникли из жаберных дуг. Поэтому жевательные мышцы, возникающие из челюстной дуги и мышцы дна рта, располагаются и прикрепляются на нижней челюсти и иннервируются тройничным нервом (V пара). Из мускулатуры, соответствующей 2-й жаберной дуге, происходит главным образом подкожная мускулатура шеи и головы, иннер-вируемая лицевым нервом (VII пара).
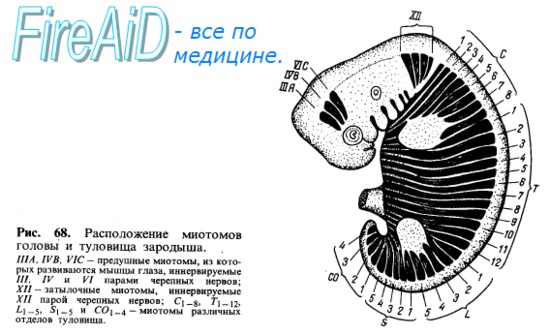
Мышцы, возникающие из материала обеих жаберных дуг, имеют двойное прикрепление и двойную иннервацию, например двубрюшная мышца, переднее брюшко которой прикрепляется к нижней челюсти (иннервация из тройничного нерва), а заднее — к подъязычной кости (иннервация из лицевого нерва). Висцеральная мускулатура, иннервируемая IX и X парами черепных нервов, у наземных позвоночных частью редуцируется, частью идет на образование мышц глотки и гортани. Трапециевидная мышца Теряет всякую связь с жаберными дугами и становится исключительно мышцей пояса верхней конечности. У млекопитающих от нее отщепляется в виде отдельной части грудино-ключично-сосцевидная мышца. Задняя ветвь блуждающего нерва, иннер-вирующая трапециевидную мышцу, превращается у высших позвоночных в самостоятельный черепной нерв — п. accessorius. Так как мозговой череп во всех своих частях представляет неподвижное образование, то на нем ожидать развития мышц нельзя. Поэтому на голове встречаются только некоторые остатки мускулатуры, образовавшейся из головных сомитов. К числу их нужно отнести мышцы глаза, происходящие из так называемых предушных миотомов (иннервация от III, IV и VI пар черепных нервов) (см. рис. 68).
Затылочные миотомы вместе с передними туловищными миотомами обычно образуют путем вентральных отростков особую поджаберную или подъязычную мускулатуру, лежащую под висцеральным скелетом. За счет этой мускулатуры, проникающей кпереди до нижней челюсти, происходят у наземных позвоночных мышцы языка, снабжаемые в силу своего происхождения из затылочных сомитов комплексом нервных волокон, образующих подъязычный нерв, который только у высших позвоночных стал настоящим черепным нервом. Остальная часть подъязычной мускулатуры (ниже подъязычной кости) представляет собой продолжение вентральной мускулатуры туловища, иннервируемой от передних ветвей спинномозговых нервов. Таким образом, для понимания расположения и фиксации мышц надо учитывать, кроме их функции, также и развитие (см. рис. 66, 67).
1. Мышечные ткани. Источники развития. Общая морфо-функциональная характеристика. Классификация. Возможности регенерации.
Мышечными тканями называют ткани, различные по строению и происхождению, но сходные по способности к выраженным сокращениям. Они обеспечивают перемещения в пространстве организма в целом, его частей и движение органов внутри организма.
Основные морфологические признаки элементов мышечных тканей- удлиненная форма, наличие продольно расположенных миофибрилл и миофиламентов- специальных органелл, обеспечивающих сократимость, расположение митохондрий рядом с сократительными элементами, наличие включений гликогена, липидов и миоглобина.
Специальные сократительные органеллы- миофиламенты или миофибриллы обеспечивают сокращение, которое возникает при взаимодействии в них двух основных фибриллярных белков- актина и миозина при обязательном участии ионов кальция . Митохондрии обеспечивают эти процессы энергией. Запас источников энергии образует гликоген липиды. Миоглобин- белок, обеспечивающий связывание кислорода и создание его запаса на момент сокращения мышцы, когда сдавливаются кровеносные сосуда.
Классификация.В соответствии с морфофункциональным принципом, в зависимости от структуры органелл сокращения, мышечные ткани подразделяют на две подгруппы.
Первая подгруппа- поперечнополосатые мышечные ткани.
Вторая подгруппа – гладкие мышечные ткани.
В соответствии с гистогенетическим принципом в зависимости от источников развития мышечные ткани подразделяются на 5 типов: мезенхимные( из десмального зачатка в составе мезенхимы), эпидермальные (из кожной эктодермы и из прехордальной пластинки), нейральные ( из нервной трубки), целомические( из миоэпикардиальной пластинки висцерального листка сомита) и соматические(миотомные).
Первые три типа относятся к подгруппе гладких мышечных тканей, четвертый и пятый- к подгруппе поперечнополосатых.
2. Гладкая мышечная ткань. Источник развития. Морфо- функциональная характеристика гладких мышечных тканей. Структурные основы сокращения. Иннервация. Регенерация.
Различают три группы гладких мышечных тканей- мезенхимные, эпидермальные и нейральные.
Стволовые клетки и клетки – предшественники в гладкой мышечной ткани на этапах эмбрионального развития пока точно не отождествлены. Они мигрируют к местам закладки органов, будучи уже детерминированными. Дифференцируясь, они синтезируют компоненты матрикса и коллагена базальной мембраны, а также эластина. У дефинитивных клеток синтетическая способности снижена, но не исчезает полностью.
Гладкий миоцит- веретеновидная клетка длиной 20-500 мкм, шириной 5-8 мкм. Ядро палочковидное, находится в ее центральной части. Когда миоцит сокращается, его ядро изгибается и даже закручивается. Филаменты актина образуют в цитоплазме трехмерную сеть, вытянутую преимущественно продольно. Концы филаментов скреплены между собой и с плазмолеммой специальными сшивающими белками. Мономеры миозина располагаются рядом с филаментами актина. Сигнал к сокращению обычно поступает по нервным волокнам. Медиатор, который выделяется из их терминалей, изменяет состояние плазмолеммы. Она образует выпячивания- кавеолы,в которых концентрируются ионы кальция. Кавеолы отшнуровываются в сторону цитоплазмы в виде пузырьков. Это влечет за собой взаимодействие миозина с актином. Актиновые филаменты смещаются друг другу навстречу, плотные пятна сближаются, усилие передается на плазмолемму, и вся клетка укорачивается. Когда поступление сигналов со стороны нервной системы прекращается, ионы кальция эвакуируются из кавеол, миозин деполимеризуется и « миофибриллы» распадаются. Сокращение прекращается. Гладкая МТ иннервируется вегетативной нервной системой, т.е. не подчиняется воле человека. Сокращение ГМТ медленное — тоническое, зато ГМТ малоутомляема. ГМТ в эмбриональном периоде развивается из мезенхимы. Вначале мезенхимные клетки имеют звездчатую, отросчатую форму, а при дифференцировке в ГМ-клетки приобретают веретеновидную форму; в цитоплазме накапливаются органоиды спецназначения — миофибриллы из актина и миозина. Регенерация ГМТ: 1. Митоз миоцитов после дедифференцировки: миоциты утрачивают сократительные белки, исчезают митохондрии и превращаются в миобласты. Миобласты начинают размножаться, а потом вновь дифференцируются в зрелые леомиоциты. 2. Возможно образование новых ГМ-клеток из малодифференцированных стволовых клеток фибробластического дифферона рыхлой с.д.Физиологическая регенерация гладкой мышечной ткани проявляется в условиях повышенных функциональных нагрузок. Наиболее отчетливо это видно в мышечной оболочке матки при беременности. Такая регенерация осуществляется не столько на тканевом, сколько на клеточном уровне: миоциты растут, в цитоплазме активизируются синтетические процессы, количество миофиламентов увеличивается. Не исключена, однако, и пролиферация клеток.
3. Скелетная поперечно-полосатая мышечная ткань. Источники развития. Морфо-функциональная характеристика миосимпласта. Структурные основы сокращения. Мышца как орган. Типы мышечных волокон. Регенерация.
Источником развития элементов скелетной поперечнополосатой мышечной ткани являются клетки миотомов- миобласты.Одни из них дифференцируются на месте и участвуют в образовании так называемых аутохтонных мышц. Другие клетки мигрируют из миотомов в мезенхиму. В ходе дифференцировки возникают две клеточные линии. Клетки одной из линий сливаются, образуя удлиненные сиппласты-мышечные трубочки.
В них происходит дифференцировка специальных органелл- миофибрилл. Клетки другой линии остаются самостоятельными и дифференцируются в миосимпластов. Основной структурной единицей скелетной мышечной ткани является мышечное волокно, состоящие из миосимпласта миосателлитоцитов, покрытых общей базальной мембраной.
Миосимпласт имеет множество продолговатых ядер, расположенных непосредственно под сарколеммой. Их количество в одном симпласте может достигать нескольких десятков тысяч. Миофибриллы заполняют основную часть миосимпласта и расположены продольно.
Типы мышечных волокон. Мышечные волокна в составе разных мышц обладают разной силой, скоростью и длительностью сокращения, а также утомляемостью. Ферменты в них обладают разной активностью и представлены в различных изомерных формах. Заметно различие в них содержания дыхательных ферментов- гликолитических и окислительных. По соотношению миофибрилл, митохондрий и миоглобина различаютбелые, красные и промежуточные волокна. По функциональным особенностям мышечные волокна подразделяют набыстрые, медленные, и промежуточные.Обычно в быстрых волокнах преобладают гликолитические процессы, они более богатыгликогеном, в них меньше миоглобина, поютому их называют такжебелыми.В медленных волокнах, напротив, выше активность окислительных ферментов, они богачемиоглобином, выглядят более красными.
Регенерация. Ядра миосимпластов делиться не могут, так как у них отсутствуют клеточные центры. Камбиальными элементами служат миосателлитоциты. Пока организм растет, они делятся, а дочерние клетки встраиваются в концы симпластов. После повреждения мышечного волокна на некотором протяжении от места травмы оно разрушается и его фрагменты фагоцитируются макрофагами. Восстановление тканей осуществляется за счет двух механизмов: компенсаторной гипертрофии самого симпласта и пролифирации миосателлитоцитов. В симпласте активизируются гранулярная эндоплазматическая сеть и аппарат Гольджи. Поврежденный конец миосимпластаутолщается, образуя мышечную почку.Миосателлитоциты, сохранившиеся рядом с повреждением, делятся. Одни из них мигрируют к мышечной почке и встраиваются в нее, другие сливаются и образуют миотубы, которые затем входят в состав вновь образованных мышечных волокон или формируют новые волокна.
Скелетная мышца как орган.Передача усилий сокращения на скелет осуществляется посредством сухожилий или прикрепления мышц непосредственно к надкостнице.На конце каждого мышечного волокна плазмолемма образует глубокие узкие впячивания.В них со стороны сухожилия или надкостницы проникают тонкиеколлагеновые волокна. Последние спирально оплетаются ретикулярными волокнами. Концы волокон направляются к базальной мембране, входят в неё, поворачивают назад и по выходе снова оплетают коллагеновые волокна соед.ткани.Между мышечными волокнами находятся тонкие прослойки рыхлой волокнистой соед.ткани— эндомизий.Более толстые прослойки рыхлой соединт.ткани окружают по несколько мышечных волокон,образуяперемизийи разделяя мышцу на пучки. Соединительную ткань ,окружающую поверхность мышцы, называютэпимизием.
4. Сердечная поперечно-полосатая мышечная ткань. Источник развития. Морфофункциональная характеристика кардиомиоцита. Структурные основы совращения. Морфофункциональная характеристика поперечно-полосатой сердечной мышечной ткани.
Гистогенез и виды клеток. Источники развития поперечно-полосатой мышечной ткани-симметричные участки висцерального листка спланхнотома в шейной части зародыша—миоэпикардиальные пластинки.Из них дифференцируются также клетки мезотелия эпикарда. В ходе гистогенеза возникает 5 видов кардиомиоцитов- рабочие, синусные переходные, проводящие, а также секреторные.
Рабочие кардиомиоциты образуют свои цепочки. Именно они, укорачиваясь, обеспечивают силу сокращения всей сердечной мышцы. Рабочие кардиомиоциты способны передавать управляющие сигналы друг другу. Синусные кардиомиоциты способны автоматически в определенном ритме сменять состояние сокращения на состояние расслабления. Синусныекардиомиоциты передают управляющие сигналыпереходнымкардиомиоцитам, а последние-проводящим.Проводящие кардиомиоциты образуют цепочки клеток, соединенных своими концами. Первая клетка в цепочке воспринимает управляющие сигналы от синусных кардиомиоцитов и передает их далее- другим проводящим кардиомиоцитов. Клетки, замыкающие цепочку, передают сигнал через переходные кардиомиоциты рабочим. Секреторные кардиомиоциты вырабатываютнатрийуретический фактор, участвующий в процессах регуляции мочеобразования и в некоторых других процессах. Все кардиомиоциты покрыты базальной мембраной.
Строение сократительных ( рабочих) кардиомиоцитов. Клетки имеют удлиненную форму, близкую к цилиндрической Их концы соединяются друг с другом, так что цепочки клеток составляют так называемые функциональные волокна. В области контактов клеток образуются так называемые вставочные диски. Ядро кардиомиоцита овальное и лежит в центральной части клетки. У полюсов ядра сосредоточены немногочисленные органеллы общего значения. Специальные органеллы, которые обеспечивают сокращение, называются миофибриллами.Кардиомиоциты соединяются друг с другом своими торцевыми концами образуются вставочные диски. Поперечные участки выступов соседних клеток соединены друг с другом интердигитациями и десмосомами. К каждой десмосоме со стороны цитоплазмы подходит миофибрилла, закрепляющаяся концом в десмоплакиновом комплексе. Таким образом, при сокращении тяга одного кардиомиоцита передается другому. Боковые поверхности выступов кардиомиоцитов объединяются нексусами. Это создает между ними метаболические связи и обеспечивает синхронность сокращений.