
Мышечные ткани — это ткани, для которых способность к сокращению является главным свойством. Мышечные ткани составляют активную часть опорно-двигательного аппарата (пассивной частью являются кости, соединения костей).
Общими свойствами всех мышечных тканей является сократимость и возбудимость. К данной группе тканей относятся гладкая, поперечнополосатая скелетная и поперечнополосатая сердечная мышечные ткани. Клетки мышечной ткани имеют хорошо развитый цитоскелет, содержат много митохондрий.

Гладкая (висцеральная) мускулатура
Эта мышечная ткань встречается в стенках внутренних органах (бронхи, кишечник, желудок, мочевой пузырь), в стенках сосудов, протоках желез. Эволюционно является наиболее древним видом мускулатуры.
Состоит из веретенообразных миоцитов — коротких одноядерных клеток. Между клетками имеются межклеточные контакты — нексусы (лат. nexus — связь). Благодаря нексусам возбуждение, возникшее в одной клетке, волнообразно распространяется на все остальные клетки.

Гладкая мышечная ткань отличается своей способностью к длительному тоническому напряжению, что очень важно для работы внутренних органов (к примеру мочевого пузыря), сокращается медленно, практически не утомляется. Скелетная мышечная ткань, которую мы изучим чуть позже, такой способностью не обладает — сокращается и утомляется быстро.
Осуществляется сокращение с помощью клеточных органоидов — миофиламентов, которые расположены в клетке хаотично и не имеют такой упорядоченной структуры, как миофибриллы в скелетной мускулатуре (все познается в сравнении, уже скоро мы их тоже изучим).
Особо заметим, что в гладкой мышечной ткани миофиламенты собираются в миофибриллы только во время сокращения. У таких временных миофибрилл не может быть регулярной организации, а значит ни у таких миофибрилл, ни у гладких миоцитов не может быть поперечной исчерченности.
Гладкая мышечная ткань сокращается непроизвольно (неподвластна воле человека). Работа гладких мышц обеспечивается вегетативной (автономной) нервной системой. К примеру невозможно по желанию сузить или расширить бронхи, кровеносные сосуды, зрачок.

Гладкая мышечная ткань называется неисчерченной, так как не обладает поперечной исчерченностью, характерной для поперечнополосатых скелетной и сердечной мышечных тканей.
Скелетная (поперечнополосатая) мышечная ткань
Скелетная мышечная ткань образует диафрагму (дыхательную мышцу), мускулатуру туловища, конечностей, головы, голосовых связок.
В отличие от гладкой мускулатуры, скелетная образована не отдельными одноядерными клетками, а длинными многоядерными волокнами, имеющими до 100 и более ядер — миосимпластами. Миосимпласт (греч. sim — вместе + plast — образованный) представляет совокупность слившихся клеток, имеет длину от нескольких миллиметров до нескольких сантиметров (соответствует длине мышцы).
Внутри миосимпласта находится саркоплазма, снаружи миосимпласт покрыт сарколеммой. Сократительные элементы — миофибриллы (лат. fibra — волоконце) — длинные тяжеобразные органеллы в миосимпласте (около 1400).

Характерная черта данной ткани — поперечная исчерченность, выражающаяся в равномерном чередовании светлых и темных полос на мышечном волокне. Это происходит потому, что границы саркомеров в соседних миофибриллах совпадают, вследствие чего все волокно приобретает поперечную исчерченность. Теперь самое время изучить микроскопическую основу мышцы — саркомер.
Саркомер (от греч. sarco — мясо (мышца) + mere — маленький)
Саркомер — элементарная сократительная единица поперечнополосатых мышц, структурная единица миофибриллы. В состав саркомера (и миофибриллы в целом) входят миофиламенты (лат. filamentum — нить) двух типов, которые обеспечивают сократимость мышечной ткани.
Саркомер состоит из актиновых (тонких) и миозиновых (толстых) филаментов, которые образованы главным образом белками актином и миозином. Сокращение происходит за счет взаимного перемещения миофиламентов: они тянутся навстречу друг другу, саркомер укорачивается (и мышца в целом).

Источником энергии для сокращения служат молекулы АТФ. К тому же невозможно представить сокращение мышц без участия ионов кальция: именно они связываются с тропонином, что приводит к изменению конформации тропомиозина (тропонин и тропомиозин — регуляторные белки между нитями актина), за счет чего становится возможно соединение актина и миозина. При сокращении мышц выделяется тепло (сократительный термогенез).
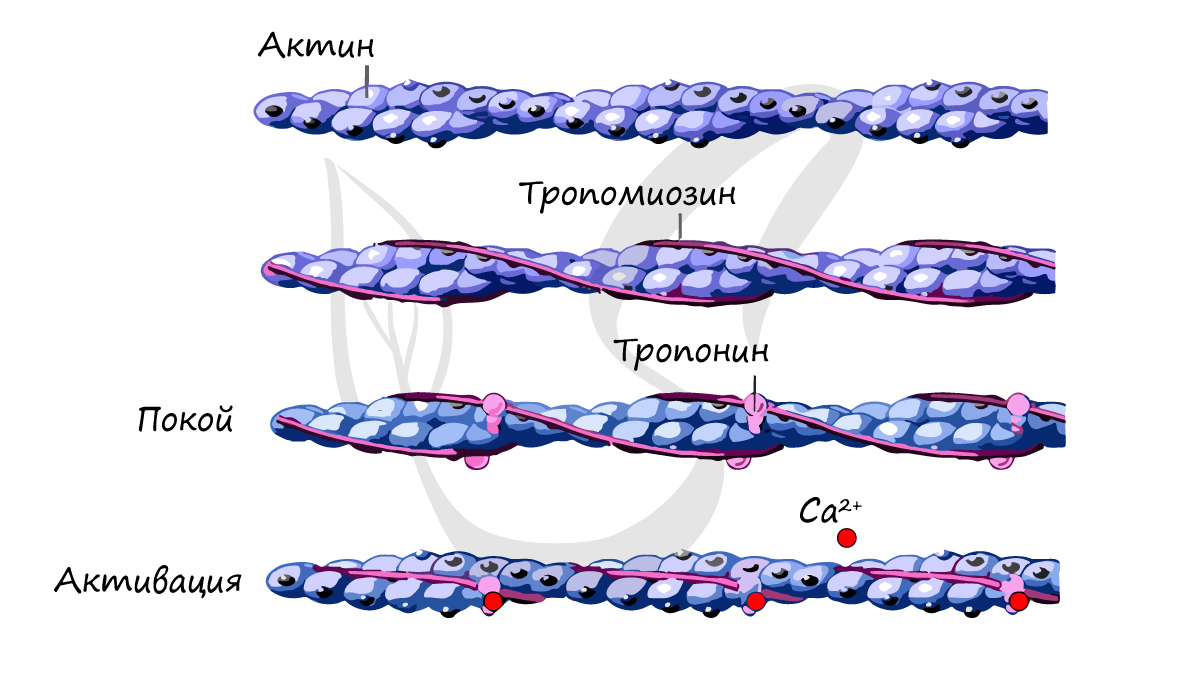
Замечу, что трупное окоченение (лат. rigor mortis) — посмертное затвердевание мышц — связано именно с ионами кальция, которые устремляются в область низкой концентрации (в саркоплазму миосимпласта), способствуя связыванию актина и миозина.
После смерти в мышце перестает синтезироваться АТФ, ее уровень быстро снижается. Как следствие этого перестает функционировать Ca-АТФаза — насос, выкачивающий ионы Ca из саркоплазмы в саркоплазматический ретикулум (мембранная органелла мышечных клеток (сходная с ЭПС), в которой запасаются ионы Ca).
В саркоплазме повышается концентрация ионов Ca — замыкаются мостики между актином и миозином, однако разомкнуться они уже не могут, в связи с чем наблюдается стойкая мышечная контрактура (лат. contractura — стягивание, сужение): конечности очень сложно разогнуть или согнуть.
Читайте также: Как люди научились делать ткань

Вернемся к скелетным мышцам. Имеется еще ряд важных моментов, о которых нужно знать.
В процесс возбуждения вовлекается изолированно один миосимпласт, соседние миосимпласты (волокна) не возбуждают друг друга, в отличие от гладких миоцитов, где возбуждение предается между соседними клетками через нексусы. Скелетные мышцы сокращаются быстро и быстро утомляются (у гладких мышц фазы сокращения и расслабления растянуты во времени, мало утомляются) .
Скелетные мышцы сокращаются произвольно: они подконтрольны нашему сознанию. К примеру, по желанию мы можем изменить скорость движения руки, темп бега, силу прыжка. Мышцы покрыты фасцией, крепятся к костям сухожилиями, и, сокращаясь, приводят в движение суставы.

Сердечная поперечнополосатая мышечная ткань
Сердечная мышечная ткань образует мышечную оболочку сердца — миокард (от др.-греч. μῦς «мышца» + καρδία — «сердце»). Миокард — средний слой сердца, составляющий основную часть его массы. При работе сердечная мышечная ткань не утомляется.

Сердечная мышечная ткань состоит из кардиомиоцитов — одиночных клеток, имеющих поперечную исчерченность. Соединяясь друг с другом, кардиомиоциты образуют функциональные волокна.
Этот тип мышечной ткани удивительным образом сочетает свойства двух предыдущих, изученных нами, тканей (возбудимость, сократимость) и имеет одно новое уникальное свойство — автоматизм.
Автоматизм — способность сердечной мышечной ткани возбуждаться и сокращаться самопроизвольно, без влияний извне. Это легко можно подтвердить, наблюдая сокращения изолированного сердца лягушки в физиологическом растворе: сокращения сердца в нем будут продолжаться несколько десятков минут после отделения сердца от организма.

Места контактов соседних кардиомиоцитов — вставочные диски (в их составе находятся нексусы), благодаря которым возбуждение одной клетки передается на соседние, таким образом волнообразно охватываются возбуждением и сокращаются новые участки миокарда.
Большое число контактов между кардиомиоцитами обеспечивает высокую эффективность и надежность проведения возбуждения по миокарду. Сокращается эта ткань непроизвольно, не утомляется.
На рисунке или микропрепарате узнать данную ткань можно по центральному положению ядер в клетках, поперечной исчерченности, наличию вставочных дисков и анастомозов (греч. anastomosis — отверстие) — мест соединений боковых поверхностей функциональных волокон (кардиомиоцитов).

В норме возбуждение проводится по проводящей системе сердца от предсердий к желудочкам (однонаправленно). Участок сердечной мышцы, в котором генерируются импульсы, определяющие частоту сердечных сокращений — водитель сердечного ритма.
Автоматизм возможен благодаря наличию в миокарде особых пейсмекерных (англ. pacemaker — задающий ритм) клеток, которые также называют водителями ритма. Они спонтанно генерируют нервные импульсы, которые охватывают весь миокард, в результате чего осуществляется сокращение. Именно благодаря водителям ритма сердце лягушки продолжает биться, будучи полностью отделенным от тела.
Ответ мышц на физическую нагрузку
Физические нагрузки приводят к гипертрофии мышц (от др.-греч. ὑπερ- чрез, слишком + τροφή — еда, пища) — в них увеличивается количество мышечных волокон, объем мышечной массы нарастает.

В условиях гиподинамии (от греч. ὑπό — под и δύνᾰμις — сила), то есть пониженной активности, мышцы уменьшаются вплоть до полной атрофии (греч. а – «не» + trophe – питание). В худшем случае волокна мышечной ткани перерождаются в соединительную ткань, после чего пациент становится обездвиженным.

Необходимо отметить, что сердечная мышечная ткань также дает ответную реакцию на чрезмерную нагрузку: сердце увеличивается в размере, нарастает масса миокарда. Причиной могут быть генетические заболевания, повышенное артериальное давление. Гипертрофия сердца — состояние, требующее вмешательства врача и наблюдения за пациентом.
В большинстве случае гипертрофия сердца обратима, а у спортсменов наблюдается так называемая физиологическая гипертрофия (вариант нормы).

Происхождение мышц
Мышцы развиваются из среднего зародышевого листка — мезодермы.

© Беллевич Юрий Сергеевич 2018-2022
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
1. Мышечные ткани. Источники развития. Общая морфо-функциональная характеристика. Классификация. Возможности регенерации.
Мышечными тканями называют ткани, различные по строению и происхождению, но сходные по способности к выраженным сокращениям. Они обеспечивают перемещения в пространстве организма в целом, его частей и движение органов внутри организма.
Читайте также: Как убрать кнопки с ткани
Основные морфологические признаки элементов мышечных тканей- удлиненная форма, наличие продольно расположенных миофибрилл и миофиламентов- специальных органелл, обеспечивающих сократимость, расположение митохондрий рядом с сократительными элементами, наличие включений гликогена, липидов и миоглобина.
Специальные сократительные органеллы- миофиламенты или миофибриллы обеспечивают сокращение, которое возникает при взаимодействии в них двух основных фибриллярных белков- актина и миозина при обязательном участии ионов кальция . Митохондрии обеспечивают эти процессы энергией. Запас источников энергии образует гликоген липиды. Миоглобин- белок, обеспечивающий связывание кислорода и создание его запаса на момент сокращения мышцы, когда сдавливаются кровеносные сосуда.
Классификация.В соответствии с морфофункциональным принципом, в зависимости от структуры органелл сокращения, мышечные ткани подразделяют на две подгруппы.
Первая подгруппа- поперечнополосатые мышечные ткани.
Вторая подгруппа – гладкие мышечные ткани.
В соответствии с гистогенетическим принципом в зависимости от источников развития мышечные ткани подразделяются на 5 типов: мезенхимные( из десмального зачатка в составе мезенхимы), эпидермальные (из кожной эктодермы и из прехордальной пластинки), нейральные ( из нервной трубки), целомические( из миоэпикардиальной пластинки висцерального листка сомита) и соматические(миотомные).
Первые три типа относятся к подгруппе гладких мышечных тканей, четвертый и пятый- к подгруппе поперечнополосатых.
2. Гладкая мышечная ткань. Источник развития. Морфо- функциональная характеристика гладких мышечных тканей. Структурные основы сокращения. Иннервация. Регенерация.
Различают три группы гладких мышечных тканей- мезенхимные, эпидермальные и нейральные.
Стволовые клетки и клетки – предшественники в гладкой мышечной ткани на этапах эмбрионального развития пока точно не отождествлены. Они мигрируют к местам закладки органов, будучи уже детерминированными. Дифференцируясь, они синтезируют компоненты матрикса и коллагена базальной мембраны, а также эластина. У дефинитивных клеток синтетическая способности снижена, но не исчезает полностью.
Гладкий миоцит- веретеновидная клетка длиной 20-500 мкм, шириной 5-8 мкм. Ядро палочковидное, находится в ее центральной части. Когда миоцит сокращается, его ядро изгибается и даже закручивается. Филаменты актина образуют в цитоплазме трехмерную сеть, вытянутую преимущественно продольно. Концы филаментов скреплены между собой и с плазмолеммой специальными сшивающими белками. Мономеры миозина располагаются рядом с филаментами актина. Сигнал к сокращению обычно поступает по нервным волокнам. Медиатор, который выделяется из их терминалей, изменяет состояние плазмолеммы. Она образует выпячивания- кавеолы,в которых концентрируются ионы кальция. Кавеолы отшнуровываются в сторону цитоплазмы в виде пузырьков. Это влечет за собой взаимодействие миозина с актином. Актиновые филаменты смещаются друг другу навстречу, плотные пятна сближаются, усилие передается на плазмолемму, и вся клетка укорачивается. Когда поступление сигналов со стороны нервной системы прекращается, ионы кальция эвакуируются из кавеол, миозин деполимеризуется и « миофибриллы» распадаются. Сокращение прекращается. Гладкая МТ иннервируется вегетативной нервной системой, т.е. не подчиняется воле человека. Сокращение ГМТ медленное — тоническое, зато ГМТ малоутомляема. ГМТ в эмбриональном периоде развивается из мезенхимы. Вначале мезенхимные клетки имеют звездчатую, отросчатую форму, а при дифференцировке в ГМ-клетки приобретают веретеновидную форму; в цитоплазме накапливаются органоиды спецназначения — миофибриллы из актина и миозина. Регенерация ГМТ: 1. Митоз миоцитов после дедифференцировки: миоциты утрачивают сократительные белки, исчезают митохондрии и превращаются в миобласты. Миобласты начинают размножаться, а потом вновь дифференцируются в зрелые леомиоциты. 2. Возможно образование новых ГМ-клеток из малодифференцированных стволовых клеток фибробластического дифферона рыхлой с.д.Физиологическая регенерация гладкой мышечной ткани проявляется в условиях повышенных функциональных нагрузок. Наиболее отчетливо это видно в мышечной оболочке матки при беременности. Такая регенерация осуществляется не столько на тканевом, сколько на клеточном уровне: миоциты растут, в цитоплазме активизируются синтетические процессы, количество миофиламентов увеличивается. Не исключена, однако, и пролиферация клеток.
3. Скелетная поперечно-полосатая мышечная ткань. Источники развития. Морфо-функциональная характеристика миосимпласта. Структурные основы сокращения. Мышца как орган. Типы мышечных волокон. Регенерация.
Источником развития элементов скелетной поперечнополосатой мышечной ткани являются клетки миотомов- миобласты.Одни из них дифференцируются на месте и участвуют в образовании так называемых аутохтонных мышц. Другие клетки мигрируют из миотомов в мезенхиму. В ходе дифференцировки возникают две клеточные линии. Клетки одной из линий сливаются, образуя удлиненные сиппласты-мышечные трубочки.
В них происходит дифференцировка специальных органелл- миофибрилл. Клетки другой линии остаются самостоятельными и дифференцируются в миосимпластов. Основной структурной единицей скелетной мышечной ткани является мышечное волокно, состоящие из миосимпласта миосателлитоцитов, покрытых общей базальной мембраной.
Читайте также: Мебель ткани кожа обивка
Миосимпласт имеет множество продолговатых ядер, расположенных непосредственно под сарколеммой. Их количество в одном симпласте может достигать нескольких десятков тысяч. Миофибриллы заполняют основную часть миосимпласта и расположены продольно.
Типы мышечных волокон. Мышечные волокна в составе разных мышц обладают разной силой, скоростью и длительностью сокращения, а также утомляемостью. Ферменты в них обладают разной активностью и представлены в различных изомерных формах. Заметно различие в них содержания дыхательных ферментов- гликолитических и окислительных. По соотношению миофибрилл, митохондрий и миоглобина различаютбелые, красные и промежуточные волокна. По функциональным особенностям мышечные волокна подразделяют набыстрые, медленные, и промежуточные.Обычно в быстрых волокнах преобладают гликолитические процессы, они более богатыгликогеном, в них меньше миоглобина, поютому их называют такжебелыми.В медленных волокнах, напротив, выше активность окислительных ферментов, они богачемиоглобином, выглядят более красными.
Регенерация. Ядра миосимпластов делиться не могут, так как у них отсутствуют клеточные центры. Камбиальными элементами служат миосателлитоциты. Пока организм растет, они делятся, а дочерние клетки встраиваются в концы симпластов. После повреждения мышечного волокна на некотором протяжении от места травмы оно разрушается и его фрагменты фагоцитируются макрофагами. Восстановление тканей осуществляется за счет двух механизмов: компенсаторной гипертрофии самого симпласта и пролифирации миосателлитоцитов. В симпласте активизируются гранулярная эндоплазматическая сеть и аппарат Гольджи. Поврежденный конец миосимпластаутолщается, образуя мышечную почку.Миосателлитоциты, сохранившиеся рядом с повреждением, делятся. Одни из них мигрируют к мышечной почке и встраиваются в нее, другие сливаются и образуют миотубы, которые затем входят в состав вновь образованных мышечных волокон или формируют новые волокна.
Скелетная мышца как орган.Передача усилий сокращения на скелет осуществляется посредством сухожилий или прикрепления мышц непосредственно к надкостнице.На конце каждого мышечного волокна плазмолемма образует глубокие узкие впячивания.В них со стороны сухожилия или надкостницы проникают тонкиеколлагеновые волокна. Последние спирально оплетаются ретикулярными волокнами. Концы волокон направляются к базальной мембране, входят в неё, поворачивают назад и по выходе снова оплетают коллагеновые волокна соед.ткани.Между мышечными волокнами находятся тонкие прослойки рыхлой волокнистой соед.ткани— эндомизий.Более толстые прослойки рыхлой соединт.ткани окружают по несколько мышечных волокон,образуяперемизийи разделяя мышцу на пучки. Соединительную ткань ,окружающую поверхность мышцы, называютэпимизием.
4. Сердечная поперечно-полосатая мышечная ткань. Источник развития. Морфофункциональная характеристика кардиомиоцита. Структурные основы совращения. Морфофункциональная характеристика поперечно-полосатой сердечной мышечной ткани.
Гистогенез и виды клеток. Источники развития поперечно-полосатой мышечной ткани-симметричные участки висцерального листка спланхнотома в шейной части зародыша—миоэпикардиальные пластинки.Из них дифференцируются также клетки мезотелия эпикарда. В ходе гистогенеза возникает 5 видов кардиомиоцитов- рабочие, синусные переходные, проводящие, а также секреторные.
Рабочие кардиомиоциты образуют свои цепочки. Именно они, укорачиваясь, обеспечивают силу сокращения всей сердечной мышцы. Рабочие кардиомиоциты способны передавать управляющие сигналы друг другу. Синусные кардиомиоциты способны автоматически в определенном ритме сменять состояние сокращения на состояние расслабления. Синусныекардиомиоциты передают управляющие сигналыпереходнымкардиомиоцитам, а последние-проводящим.Проводящие кардиомиоциты образуют цепочки клеток, соединенных своими концами. Первая клетка в цепочке воспринимает управляющие сигналы от синусных кардиомиоцитов и передает их далее- другим проводящим кардиомиоцитов. Клетки, замыкающие цепочку, передают сигнал через переходные кардиомиоциты рабочим. Секреторные кардиомиоциты вырабатываютнатрийуретический фактор, участвующий в процессах регуляции мочеобразования и в некоторых других процессах. Все кардиомиоциты покрыты базальной мембраной.
Строение сократительных ( рабочих) кардиомиоцитов. Клетки имеют удлиненную форму, близкую к цилиндрической Их концы соединяются друг с другом, так что цепочки клеток составляют так называемые функциональные волокна. В области контактов клеток образуются так называемые вставочные диски. Ядро кардиомиоцита овальное и лежит в центральной части клетки. У полюсов ядра сосредоточены немногочисленные органеллы общего значения. Специальные органеллы, которые обеспечивают сокращение, называются миофибриллами.Кардиомиоциты соединяются друг с другом своими торцевыми концами образуются вставочные диски. Поперечные участки выступов соседних клеток соединены друг с другом интердигитациями и десмосомами. К каждой десмосоме со стороны цитоплазмы подходит миофибрилла, закрепляющаяся концом в десмоплакиновом комплексе. Таким образом, при сокращении тяга одного кардиомиоцита передается другому. Боковые поверхности выступов кардиомиоцитов объединяются нексусами. Это создает между ними метаболические связи и обеспечивает синхронность сокращений.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом