
Из какого эмбрионального зачатка развивается скелетная мышечная ткань
6. Гистогенез и регенерация мышечной ткани
Гистогенез скелетной мышечной ткани . Источником развития элементов скелетной (соматической) поперечнополосатой мышечной ткани являются клетки миотомов мезодермы – миобласты. Одни из них дифференцируются на месте и участвуют в образовании так называемых аутохтонных мышц. Другие клетки мигрируют из миотомов в мезенхиму. Они уже детерминированы, хотя внешне не отличаются от других клеток мезенхимы. Их дифференцировка продолжается в местах закладки других мышц тела.
В ходе дифференцировки возникают две клеточные линии. Клетки одной из линий сливаются, образуя удлиненные симпласты – мышечные трубочки ( миотубы ). В них происходит дифференцировка специальных органелл – миофибрилл. В это время в миотубах отмечается хорошо развитая гранулярная эндоплазматическая сеть. Миофибриллы сначала располагаются под плазмолеммой, а затем заполняют большую часть миотубы . Ядра, напротив, из центральных отделов смещаются к периферии. Клеточные центры и микротрубочки при этом полностью исчезают. Гранулярная эндоплазматическая сеть редуцируется в значительной степени. Такие дефинитивные структуры называют миосимпластами .
Клетки другой линии остаются самостоятельными и дифференцируются в миосателлитоциты ( миосателлиты ). Эти клетки располагаются на поверхности миосимпластов .
Значит, из миотомов мезодермы в определенные участки мезенхимы выселяются малодифференцированные клетки — миобласты, часть из которых выстраивается в виде цепочки в стык друг к другу и в области контактов миобластов цитолеммы исчезает и образуется симпластическое образование — миотрубка , в которой ядра в виде цепочки располагаются в середине, а по периферии начинают дифференцироваться из миофиламентов миофибриллы. К миотрубке подрастают нервные волокна, образуя двигательные нервные окончания. Под влиянием эфферентной нервной импульсации начинается перестройка мышечной трубки в мышечное волокно: ядра перемещаются на периферию симпласта к плазмолемме, а миофибриллы занимают его центральную часть, из гладкой эндоплазматической сети развивается саркоплазматическая сеть, окружающая каждую миофибриллу на всем ее протяжении. Плазмолемма миосимпласта образует глубокие трубчатые впячивания — Т-канальцы. За счет деятельности зернистой эндоплазматической сети вначале миобластов, а затем и мышечных труб синтезируются и выделяются с помощью пластинчатого комплекса белки и полисахариды, из которых формируется базальная пластинка мышечного волокна.
Следует отметить, что при формировании миотрубки , а затем и дифференцировки мышечного волокна часть миобластов не входит в состав симпласта , а прилежит к нему, располагаясь под базальной пластинкой. Эти клетки носят название миосателлитов и играют важную роль в процессах физиологической и репаративной регенерации. Установлено, что закладка поперечно-полосатых скелетных мышечных волокон ( миогенез ) происходит только в эмбриональный период. В постнатальном периоде осуществляется их дальнейшая дифференцировка и гипертрофия, но количество мышечных волокон даже в условиях интенсивной тренировки не увеличивается.
Гистогенез сердечной мышечной ткани . Источники развития сердечной поперечнополосатой мышечной ткани – симметричные участки висцерального листка спланхнотома в шейной части зародыша – миоэпикардиальные пластинки. Из них дифференцируются также клетки мезотелия эпикарда. В ходе гистогенеза стволовые клетки сердечной мышечной ткани (в составе миоэпикардиальной пластинки) в процессе дифференцировки дают начало нескольким клеточным дифферонам : возникает 5 видов кардиомиоцитов – рабочие (сократительные), синусные ( ритмзадающие или пейсмекерные ), переходные, проводящие, а также секреторные.
Исходными клетками сердечной мышечной ткани являются кардиомиобласты – уплощенные клетки (крупное ядро, светлая цитоплазма с небольшим количество рибосом и митохондрий). Происходит развитие комплекса Гольджи , гранулярной ЭПС. Клетки обладают высокой способностью к делению. Кардиомиобласты дифференцируются в кардиомиоциты . В цитоплазме кардиомиоцитов увеличивается число полисом, канальцев гранулярной ЭПС, накапливаются гранулы гликогена, увеличивается объем актомиозинового комплекса. Кардиомиоциты путем контактов «конец в конец», «конец в бок» формируют клеточные комплексы – сердечные мышечные волокна.
Гистогенез гладкой мышечной ткани . Этот вид мышечной ткани расположен в стенках органов и имеет генез мезенхимного типа. Миоциты объединяются в пучки, между которыми располагаются тонкие прослойки соединительной ткани. В эти прослойки вплетаются ретикулярные и эластические волокна, окружающие миоциты . В прослойках проходят кровеносные сосуды и нервные волокна. Терминали последних оканчиваются не непосредственно на миоцитах , а между ними. Поэтому после поступления нервного импульса медиатор распространяется диффузно, возбуждая сразу многие клетки. Гладкая мышечная ткань мезенхимного происхождения представлена главным образом в стенках кровеносных сосудов и многих трубчатых внутренних органов, а также образует отдельные мелкие мышцы (цилиарные). Гладкая мышечная ткань в составе конкретных органов имеет неодинаковые функциональные свойства. Это обусловлено тем, что на поверхности органов имеются разные рецепторы к конкретным биологически активным веществам. Поэтому и на многие лекарственные препараты их реакция неодинакова. Возможно, разные функциональные свойства тканей связаны и с конкретной молекулярной организацией актиновых филаментов .
Как было сказано выше, гладкие мышцы развиваются из мезенхимы. Клетки мезенхимы начинают активно размножаться и образуют сгущения. Первым признаков начавшейся дифференциации служит возникновение в цитоплазме клеток протофибрилл . Малодифференцированные клетки располагаются рыхло, соединяются между собой отростками (как клетки мезенхимы). Миофиламенты не имеют правильной ориентации. Позднее гладкомышечные клетки становятся веретенообразными и плотно прилегают друг к другу. Количество протофибрилл в них увеличивается, и они располагаются только в направлении длинной оси клетки.
Читайте также: Прожег ткань как заделать
В активированных миоцитах увеличивается синтез сократительных белков, происходит укрупнение и гиперхроматоз ядра, гипертрофия ядрышка, увеличивается количество свободных рибосом и полисом, наблюдается активация ферментов. Клеточная регенерация осуществляется как за счет активации миоцитов малого объема (камбиальных элементов), так и за счет дифференцированных клеток (обладающих способностью к митозу).
При функциональной нагрузке (беременности) или при патологии наблюдается увеличение массы гладкомышечной ткани за счет гипертрофии (увеличения размеров) или за счет гиперплазии (увеличения количества) гладких миоцитов
Мышечная ткань эпидермального происхождения Миоэпителиальные клетки развиваются из эпидермального зачатка. Они встречаются в потовых, молочных, слюнных и слезных железах и имеют общих предшественников с их секреторными клетками. Миоэпителиальные клетки непосредственно прилежат к собственно эпителиальным и имеют общую с ними базальную мембрану. При регенерации те и другие клетки тоже восстанавливаются из общих малодифференцированных предшественников. Большинство миоэпителиальных клеток имеют звездчатую форму. Эти клетки нередко называют корзинчатыми : их отростки охватывают концевые отделы и мелкие протоки желез. В теле клетки располагаются ядро и органеллы общего значения, а в отростках – сократительный аппарат, организованный, как и в клетках мышечной ткани мезенхимного типа.
Мышечная ткань нейрального происхождения. Миоциты этой ткани развиваются из клеток нейрального зачатка в составе внутренней стенки глазного бокала. Тела этих клеток располагаются в эпителии задней поверхности радужки. Каждая из них имеет отросток, который направляется в толщу радужки и ложится параллельно ее поверхности. В отростке находится сократительный аппарат, организованный так же, как и во всех гладких миоцитах . В зависимости от направления отростков (перпендикулярно или параллельно краю зрачка) миоциты образуют две мышцы – суживающую и расширяющую зрачок.
Регенерация. В мышечной, как в других тканях, различают два вида регенерации — физиологическую и репаративную . Из всех видов мышечной ткани лучше всего регенерирует гладкая, так как ее клетки миоциты могут активно делиться митозом всю жизнь.
Физиологическая регенерация скелетной мышечной ткани проявляется в форме гипертрофии мышечных волокон, что выражается в увеличении их толщины и даже длины, увеличение числа органелл, главным образом миофибрилл, а также нарастании числа ядер, что, в конечном счете, проявляется увеличением функциональной способности мышечного волокна. Радиоизотопным методом установлено, что увеличение числа ядер в мышечных волокнах в условиях гипертрофии достигается за счет деления клеток миосателлитов и последующего вхождения в миосимпласт дочерних клеток.
Увеличение числа миофибрилл осуществляется посредством синтеза актиновых и миозиновых белков свободными рибосомами и последующей сборки этих белков в актиновые и миозиновые миофиламенты параллельно с соответствующими филаментами саркомеров . В результате этого вначале происходит утолщение миофибрилл, а затем их расщепление и образование дочерних миофибрилл. Кроме того возможно образование новых актиновых и миозиновых миофиламентов не параллельно, а встык предшествующим миофибриллам, чем достигается их удлинение. Саркоплазматическая сеть и Т-канальцы в гипертрофирующемся волокне образуются за счет разрастания предшествующих элементов.
При определенных видах мышечной тренировки может формироваться преимущественно красный тип мышечных волокон (у стайеров) или белый тип мышечных волокон (у спринтеров). Возрастная гипертрофия мышечных волокон интенсивно проявляется с началом двигательной активности организма (1-2 года), что обусловлено, прежде всего усилением нервной стимуляции. В старческом возрасте, а также в условиях малой мышечной нагрузки наступает атрофия специальных и общих органелл, истончение мышечных волокон и снижение их функциональной способности.
Репаративная регенерация развивается после повреждения мышечных волокон. При этом способ регенерации зависит от величины дефекта. При значительных повреждениях на протяжении мышечного волокна миосателлиты в области повреждения и в прилежащих участках растормаживаются, усиленно пролиферируют, а затем мигрируют в область дефекта мышечного волокна, где выстраиваются в цепочки, формируя миотрубку . Последующая дифференцировка миотрубки приводит к восполнению дефекта и восстановлению целостности мышечного волокна. В условиях небольшого дефекта мышечного волокна на его концах, за счет регенерации внутриклеточных органелл, образуются мышечные почки, которые растут навстречу друг другу, а затем сливаются, приводя к закрытию дефекта. Однако репаративная регенерация и восстановление целостности мышечных волокон могут осуществляться при определенных условиях: во-первых, при сохраненной двигательной иннервации мышечных волокон, во-вторых, если в область повреждения не попадают элементы соединительной ткани (фибробласты). Иначе на месте дефекта мышечного волокна развивается соединительно-тканный рубец.
Читайте также: Исследование кровообращения в тканях пародонта это
Советским ученым А. Н. Студитским доказана возможность аутотрансплантации скелетной мышечной ткани и даже целых мышц при соблюдении определенных условий:
- механическое измельчение мышечной ткани трансплантата, с целью растормаживания клеток-сателлитов и последующей их пролиферации;
- помещение измельченной ткани в фасциальное ложе;
- подшивание двигательного нервного волокна к измельченному трансплантату;
- наличие сократительных движений мышц-антагонистов и синергистов.
Регенерация сердечной мышечной ткани. В сердечной мышечной ткани отсутствуют камбиальные клетки, а сами кардиомиоциты утрачивают способность делиться к моменту рождения ребенка или в первые месяцы жизни. Поэтому при регенерации ткани не образуются новые кардиомиоциты и волокна. Наблюдается гипертрофия (увеличение объема) сохранившихся клеток.
Из какого эмбрионального зачатка развивается скелетная мышечная ткань
Образование любой кости происходит за счет молодых соединительнотканных клеток мезенхимного происхождения — остеобластов, которые вырабатывают межклеточное костное вещество, играющее главную опорную роль. Соответственно отмеченным 3 стадиям развития скелета кости могут развиваться на почве соединительной или хрящевой ткани, поэтому различаются следующие виды окостенения (остеогенеза).
1.Эндесмальное окостенение (en — внутри, desme — связка) происходит в соединительной ткани первичных, покровных, костей.
На определенном участке эмбриональной соединительной ткани, имеющей очертания будущей кости, благодаря деятельности остеобластов появляются островки костного вещества (точка окостенения). Из первичного центра процесс окостенения распространяется во все стороны лучеобразно путем наложения (аппозиции) костного вещества по периферии. Поверхностные слои соединительной ткани, из которой формируется покровная кость, остаются в виде надкостницы, со стороны которой происходит увеличение кости в толщину.
2.Перихондралъное окостенение (peri — вокруг, chondros — хрящ) происходит на наружной поверхности хрящевых зачатков кости при участии надхрящницы (perichondrium).
Мезенхимный зачаток, имеющий очертания будущей кости, превращается в «кость», состоящую из хрящевой ткани и представляющую собой как бы хрящевую модель кости. Благодаря деятельности остеобластов надхрящницы, покрывающей хрящ снаружи, на поверхности его, непосредственно под надхрящницей, откладывается костная ткань, которая постепенно замещает ткань хрящевую и образует компактное костное вещество.
3. С переходом хрящевой модели кости в костную надхрящница становится надкостницей (periosteum) и дальнейшее отложение костной ткани идет за счет надкостницы — периосталъное окостенение. Поэтому перихонд-ральный и периостальный остеогенезы следуют один за другим.
4.Эндохондралъное окостенение (endo, греч. — внутри, chondros — хрящ) совершается внутри хрящевых зачатков при участии надхрящницы, которая отдает отростки, содержащие сосуды, внутрь хряща. Проникая в глубь хряща вместе с сосудами, костеобразовательная ткань разрушает хрящ, предварительно подвергшийся обызвествлению (отложение в хряще извести и перерождение его клеток), и образует в центре хрящевой модели кости островок костной ткани (точка окостенения).
Распространение процесса эндохондрального окостенения из центра к периферии приводит к формированию губчатого костного вещества. Происходит не прямое превращение хряща в кость, а его разрушение и замещение новой тканью, костной.
Характер и порядок окостенения функционально обусловлены также приспособлением организма к окружающей среде. Так, у водных позвоночных (например, костистых рыб) окостеневает путем перихондрального остеогенеза только средняя часть кости, которая, как во всяком рычаге, испытывает большую нагрузку (первичные ядра окостенения). То же наблюдается и у земноводных, у которых, однако, средняя часть кости окостеневает на большем пространстве, чем у рыб. С окончательным переходом на сушу к скелету предъявляются большие функциональные требования, связанные с более трудным, чем в воде, передвижением тела по земле и большей нагрузкой на кости.
Поэтому у наземных позвоночных появляются вторичные точки окостенения, из которых у пресмыкающихся и птиц путем эндохондрального остеогенеза окостеневают и периферические отделы костей. У млекопитающих концы костей, участвующие в сочленениях, получают даже самостоятельные точки окостенения.
Такой порядок сохраняется и в онтогенезе человека, у которого окостенение также функционально обусловлено и начинается с наиболее нагружаемых центральных участков костей.
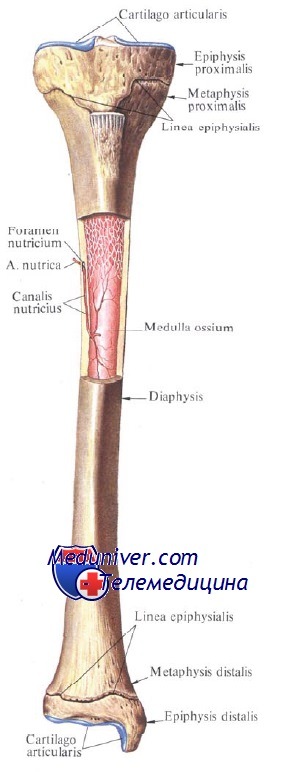
Так, сначала на 2-м месяце утробной жизни возникают первичные точки, из которых развиваются основные части костей, несущие на себе наибольшую нагрузку, т. е. тела, или диафизы, diaphysis, трубчатых костей (dia, греч.-между, phyo — расту; часть кости, растущая между эпифизами) и концы диафиза, называемые метафизами, metaphysis (meta — позади, после). Они окостеневают путем пери- и эндохондрального остеогенеза.
Затем незадолго до рождения или в первые годы после рождения появляются вторичные точки, из которых образуются путем эндохондрального остеогенеза концы костей, участвующие в сочленениях, т. е. эпифизы, epiphysis (нарост, epi — над), трубчатых костей. Возникшее в центре хрящевого эпифиза ядро окостенения разрастается и становится костным эпифизом, построенным из губчатого вещества. От первоначальной хрящевой ткани остается на всю жизнь только тонкий слой ее на поверхности эпифиза, образующий суставной хрящ.
Читайте также: Женский плащ из мембранной ткани
У детей, юношей и даже взрослых появляются добавочные островки окостенения, из которых окостеневают части кости, испытывающие тягу вследствие прикрепления к ним мышц и связок, называемые апофизами, apophysis (отросток, арo — от): например, большой вертел бедренной кости или добавочные точки на отростках поясничных позвонков, окостеневающих лишь у взрослых.
Так же функционально обусловлен и характер окостенения, связанный со строением кости. Так, кости и части костей, состоящие преимущественно из губчатого костного вещества (позвонки, грудина, кости запястья и предплюсны, эпифизы трубчатых костей и др.), окостеневают эндохондраль-но, а кости и части костей, построенные одновременно из губчатого и компактного вещества (основание черепа, диафизы трубчатых костей и др.), развиваются путем эндо- и перихондрального окостенения.
Ряд костей человека является продуктом слияния костей, самостоятельно существующих у животных. Отражая этот процесс слияния, развитие таких костей происходит за счет очагов окостенения, соответствующих по своему количеству и местоположению числу слившихся костей. Так, лопатка человека развивается из 2 костей, участвующих в плечевом поясе низших наземных позвоночных (лопатки и коракоида).
Соответственно этому, кроме основных ядер окостенения в теле лопатки, возникают очаги окостенения в ее клювовидном отростке (бывшем коракоиде). Височная кость, срастающаяся из 3 костей, окостеневает из 3 групп костных ядер. Таким образом, окостенение каждой кости отражает функционально обусловленный процесс филогенеза ее.
Рост кости
Длительный рост организма и огромная разница между размерами и формой эмбриональной и окончательной кости таковы, что делают неизбежной ее перестройку в течение роста; в процессе перестройки наряду с образованием новых остеонов идет параллельный процесс рассасывания (резорбция) старых, остатки которых можно видеть среди ново-образующихся остеонов («вставочные» системы пластинок). Рассасывание есть результат деятельности в кости особых клеток — остеокластов (clasis, греч.-ломание).
Благодаря работе последних почти вся эндохондральная кость диафиза рассасывается и в ней образуется полость (костномозговая полость). Рассасыванию подвергается также и слой перихондральной кости, но взамен исчезающей костной ткани откладываются новые слои ее со стороны надкостницы. В результате происходит рост молодой кости в толщину.
В течение всего периода детства и юности сохраняется прослойка хряща между эпифизом и метафизом, называемая эпифизарным хрящом, или пластинкой роста. За счет этого хряща кость растет в длину благодаря размножению его клеток, откладывающих промежуточное хрящевое вещество. Впоследствии размножение клеток прекращается, эпифизарный хрящ уступает натиску костной ткани и метафиз сливается с эпифизом — получается синостоз (костное сращение).
Таким образом, окостенение и рост кости есть результат жизнедеятельности остеобластов и остеокластов, выполняющих противоположные функции аппозиции и резорбции — созидания и разрушения. Поэтому на примере развития кости мы видим проявление диалектического закона единства и борьбы противоположностей. «Жить значит умирать» Источник
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом
- Правообладателям
- Политика конфиденциальности