
Как изменяется возбудимость ткани во время возбуждения
В возбудимой ткани невозможно вызвать новый потенциал действия, пока мембрана возбужденного волокна остается деполяризованной в связи с развитием предыдущего потенциала действия. Это связано с тем, что вскоре после возникновения потенциала действия натриевые каналы (или кальциевые каналы, или оба типа каналов) инактивируются, и возбуждающий сигнал любой силы, действующий на ткань в этот момент, не может открыть инактивационные ворота. Единственным условием для их повторного открытия является возврат мембранного потенциала к исходному или близкому к исходному уровню. Затем в течение небольшой доли секунды инактивационные ворота каналов открываются, и становится возможным развитие нового потенциала действия.
Период, в течение которого нельзя вызвать второй потенциал действия даже сильным стимулом, называют абсолютным рефрактерным периодом. Для крупных миелинизированных нервных волокон этот период составляет примерно 1/2500 сек. Легко рассчитать, что такое волокно может максимально передавать около 2500 имп/сек.
Кроме факторов, повышающих возбудимость нервного волокна, существуют так называемые мембраностабилизирующие факторы, способные снижать возбудимость. Например, высокая концентрация ионов кальция во внеклеточной жидкости уменьшает проницаемость мембраны для ионов натрия, снижая возбудимость. В связи с этим ионы кальция называют стабилизатором.
Местные анестетики. К наиболее важным стабилизаторам относят многие вещества, используемые в клинике в качестве местных анестетиков, в состав которых входят прокаин и тетракаин. Большинство из них действуют непосредственно на активационные ворота натриевых каналов, затрудняя их открытие, что сопровождается снижением возбудимости мембран. После снижения возбудимости до уровня, при котором отношение амплитуды потенциала действия к порогу возбуждения (называемое фактором надежности) опускается ниже 1,0, импульсы по анестезированному нерву не проходят.
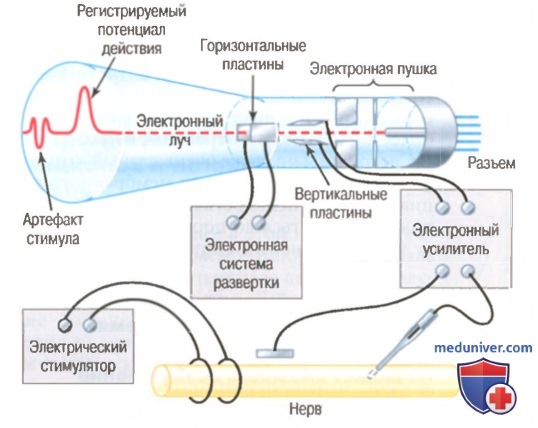
Катодно-лучевой осциллоскоп для регистрации быстрых потенциалов действия.
Катодный осциллограф. Ранее в этой главе мы отмечали, что изменения мембранного потенциала во время генерации потенциала действия происходят чрезвычайно быстро. Действительно, развитие большей части комплекса потенциала действия в крупных нервных волокнах занимает менее 1/1000 сек. На некоторых рисунках данной главы показан электроизмерительный прибор, регистрирующий эти изменения потенциалов. Однако ясно, что реакции любого прибора, способного регистрировать потенциалы действия, должны быть чрезвычайно быстрыми. Для практических целей единственным широко используемым прибором, способным точно реагировать на быстрые изменения мембранного потенциала, является катодно-лучевой осциллоскоп.
На рисунке показаны основные компоненты катодно-лучевого осциллоскопа. Катодно-лучевая трубка состоит из электронной пушки и флюоресцентного экрана, который «бомбардируется» электронами. При ударе электронов о поверхность экрана флюоресцентный материал светится. Если луч электронов перемещается по экрану, вместе с ним движется яркое пятно света, оставляя на экране флюоресцирующую линию.
Кроме электронной пушки и флюоресцентной поверхности, катодно-лучевая трубка снабжена двумя парами электрически заряженных пластин. Одна пара расположена с обеих сторон от электронного луча, а другая — сверху и снизу. Соответствующие электронные усилители изменяют напряжение на этих пластинах таким образом, что электронный луч отклоняется вверх или вниз в ответ на электрические сигналы, приходящие от регистрирующих электродов. Под влиянием внутреннего электронного блока осциллоскопа луч электронов перемещается по экрану горизонтально с постоянной скоростью. При этом регистрируется кривая, которую видно на экране катодно-лучевой трубки рисунка, с временной разверткой по горизонтали и изменениями потенциалов, регистрируемых отводящими электродами, по вертикали.
На левом конце кривой виден небольшой артефакт стимула, связанный с электрическим стимулом, который используется для вызова потенциала действия, справа на кривой — сам потенциал действия.
Видео физиология возбуждения тканей (потенциал покоя, потенциал действия) — профессор, д.м.н. П.Е. Умрюхин
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Изменения возбудимости при возбуждении
Возникновение в нервном или мышечном волокне потенциала действия сопровождается многофазными изменениями возбудимости . Для их изучения нерв или мышцу подвергают действию двух коротких, но сильных электрических стимулов, следующих друг за другом с определенным интервалом. Регистрация возникающих в ответ на эти раздражения потенциалов действия позволила установить важные факты ( рис. 136 ).
Рис. 136. Изменения возбудимости нервного волокна в различные фазы развития потенциала действия и следовых изменений мембранного потенциала (по Л. Моргану). Для наглядности длительность первых двух фаз на каждой кривой несколько увеличена. Пунктирной линией на рис. А обозначен потенциал покоя, а на рис. б — исходный уровень возбудимости.
Периоду возникновения и развития пика потенциала действия соответствует полное исчезновение возбудимости, получившее название обсолютной рефрактерности. В эту фазу второе раздражение не способно вызвать новый потенциал действия, как бы сильно ни было это раздражение.
Длительность абсолютной рефрактерности варьирует у различных возбудимых тканей в широких пределах. В быстро проводящих нервных волокнах теплокровных животных ее продолжительность составляет около 0,5 мсек. В волокнах сердечной мышцы, характеризующихся очень длительным платообразным потенциалом действия , абсолютная рефрактерная фаза продолжается около 250—300 мсек.
Вслед за абсолютной рефрактерностью начинается фаза относительной рефрактерности. В нервных волокнах она длится 4—8 мсек. В это время возбудимость постепенно возвращается к первоначальному уровню, имевшемуся до начала первого раздражения. В период относительной рефрактерности нервное или мышечное волокно способно ответить на сильное раздражение, но амплитуда потенциала действия оказывается резко сниженной.
Лишь по мере увеличения длительности интервала между первым и вторым раздражением амплитуда потенциала действия на второе раздражение постепенно повышается. Рис. 137 иллюстрирует эту закономерность.
Рис. 137. Изменения потенциалов действия нервного волокна в фазу относительной рефрактерности (по Брюкке, Ирли Форбсу). На нерв наносятся многократно два раздражения максимальной силы с интервалами1,4; 1,6; 2,0; 2,6; 3,6 и 4,4 мсек. (Потенциалы действия на первые раздражения слились)
Согласно ионной теории возбуждения Ходжкина — Хаксли, абсолютная рефрактерность обусловлена изменениями свойств мембраны, сходными с теми, которые развиваются при катодической депрессии и аккомодации, а именно: мембрана утрачивает способность отвечать повышением натриевой проницаемости на действие раздражителя (инактивация) и в то же время повышается ее проницаемость для ионов Кֺ. Оба эти процесса препятствуют возникновению потенциала действия на новое раздражение.
Абсолютная рефрактерная фаза резко удлиняется при охлаждении нервных и мышечных волокон. Это позволяет предполагать, что в основе указанных выше изменении ионной проницаемости мембраны лежат какие-то химические процессы.
Как изменяется возбудимость ткани во время возбуждения
В основе всех физиологических реакций лежит способность живых клеток реагировать на раздражитель. Раздражитель – любое изменение внешней или внутренней среды, которое действует на клетку или многоклеточную систему (ткань, организм).
Раздражители
По природе раздражители подразделяют на:
физические (звук, свет, температура, вибрация, осмотическое давление), особое значение для биологических систем имеют электрические раздражители;
химические (ионы, гормоны, нейромедиаторы, пептиды, ксенобиотики);
информационные (голосовые команды, условные знаки, условные стимулы).
По биологическому значению раздражители подразделяют на:
адекватные – раздражители, для восприятия которых биологическая система имеет специальные приспособления;
неадекватные – раздражители, не соответствующие природной специализации рецепторных клеток, на которые они действуют.
Раздражитель вызывает возбуждение только в том случае, если он достаточно силен. Порог возбуждения – минимальная сила раздражителя, достаточная для того, чтобы вызвать возбуждение клетки. Выражение «порог возбуждения» имеет несколько синонимов: порог раздражения, пороговая сила раздражителя, порог силы.
Возбуждение как активная реакция клетки на раздражитель
Реакция клетки на внешнее воздействие (раздражение) отличается от реакции небиологических систем следующими особенностями:
энергией для реакции клетки служит не энергия раздражителя, а энергия, образующаяся в результате метаболизма в самой биологической системе;
сила и форма реакции клетки не определяется силой и формой внешнего воздействия (если сила раздражителя выше пороговой).
В некоторых специализированных клетках реакция на раздражитель проявляется особенно интенсивно. Такую интенсивную реакцию называют возбуждением. Возбуждение – активная реакция специализированных (возбудимых) клеток на внешнее воздействие, проявляющаяся в том, что клетка начинает выполнять присущие ей специфические функции.
Возбудимая клетка может находиться в двух дискретных состояниях:
состоянии покоя (готовность к реагированию на внешнее воздействие, совершение внутренней работы);
состоянии возбуждения (активное выполнение специфических функций, совершение внешней работы).
В организме существует 3 типа возбудимых клеток:
нервные клетки (возбуждение проявляется генерацией электрического импульса);
мышечные клетки (возбуждение проявляется сокращением);
секреторные клетки (возбуждение проявляется выбросом в межклеточное пространство биологически активных веществ).
Возбудимость – способность клетки переходить из состояния покоя в состояние возбуждения при действии раздражителя. Разные клетки имеют различную возбудимость. Возбудимость одной и той же клетки меняется в зависимости от ее функционального состояния.
Возбудимая клетка в состоянии покоя
Мембрана возбудимой клетки поляризована. Это означает, что имеется постоянная разность потенциалов между внутренней и наружной поверхностью клеточной мембраны, которую называют мембранный потенциал (МП). В состоянии покоя величина МП составляет –60…–90 мВ (внутренняя сторона мембраны заряжена отрицательно относительно наружной). Значение МП клетки в состоянии покоя называют потенциалом покоя (ПП). МП клетки можно измерять, разместив один электрод внутри, а другой снаружи клетки (рис. 1 А ) .
Рис. 1. Схема регистрации мембранного потенциала клетки (А); мембранный потенциал клетки в состоянии покоя и его возможные изменения (Б).
Уменьшение МП относительно его нормального уровня (ПП) называют деполяризацией , а увеличение – гиперполяризацией . Под реполяризацией понимают восстановление исходного уровня МП после его изменения (см. рис. 1 Б).
Электрические и физиологические проявления возбуждения
Рассмотрим различные проявления возбуждения на примере раздражения клетки электрическим током (рис. 2).
Рис. 2. Изменение мембранного потенциала клетки (А) при действии электрического тока различной силы (Б).
При действии слабых (подпороговых) импульсов электрического тока в клетке развивается электротонический потенциал. Электротонический потенциал (ЭП) – сдвиг мембранного потенциала клетки, вызываемый действием постоянного электрического тока . ЭП есть пассивная реакция клетки на электрический раздражитель; состояние ионных каналов и транспорт ионов при этом не изменяется. ЭП не проявляется физиологической реакцией клетки. Поэтому ЭП не является возбуждением.
При действии более сильного подпорогового тока возникает более пролонгированный сдвиг МП – локальный ответ. Локальный ответ (ЛО) – активная реакция клетки на электрический раздражитель, однако состояние ионных каналов и транспорт ионов при этом изменяется незначительно. ЛО не проявляется заметной физиологической реакцией клетки. ЛО называют местным возбуждением , так как это возбуждение не распространяется по мембранам возбудимых клеток.
При действии порогового и сверхпорогового тока в клетке развивается потенциал действия (ПД). ПД характеризуется тем, что значение МП клетки очень быстро уменьшается до 0 (деполяризация), а затем мембранный потенциал приобретает положительное значение (+20…+30 мВ), т. е. внутренняя сторона мембраны заряжается положительно относительно наружной. Затем значение МП быстро возвращается к исходному уровню. Сильная деполяризация клеточной мембраны во время ПД приводит к развитию физиологических проявлений возбуждения (сокращение, секреция и др.). ПД называют распространяющимся возбуждением , поскольку, возникнув в одном участке мембраны, он быстро распространяется во все стороны.
Механизм развития ПД практически одинаков для всех возбудимых клеток. Механизм сопряжения электрических и физиологических проявлений возбуждения различен для разных типов возбудимых клеток (сопряжение возбуждения и сокращения, сопряжение возбуждения и секреции).
Устройство клеточной мембраны возбудимой клетки
В механизмах развития возбуждения участвуют 4 вида ионов: K+ , Na+ , Ca++ , Cl – (ионы Ca++ участвуют в процессах возбуждения некоторых клеток, например кардиомиоцитов, а ионы Cl – важны для развития торможения). Мембрана клетки, представляющая собой липидный бислой, непроницаема для этих ионов. В мембране существуют 2 типа специализированных интегральных белковых систем, которые обеспечивают транспорт ионов через клеточную мембрану: ионные насосы и ионные каналы.
Ионные насосы и трансмембранные ионные градиенты
Ионные насосы (помпы) – интегральные белки, которые обеспечивают активный перенос ионов против градиента концентрации. Энергией для транспорта служит энергия гидролиза АТФ. Различают Na+ / K+ помпу (откачивает из клетки Na+ в обмен на К+ ), Ca++ помпу (откачивает из клетки Ca++ ), Cl– помпу (откачивает из клетки Cl – ).
В результате работы ионных насосов создаются и поддерживаются трансмембранные ионные градиенты:
концентрация Na+, Ca++, Cl – внутри клетки ниже, чем снаружи (в межклеточной жидкости);
концентрация K+ внутри клетки выше, чем снаружи.
Ионные каналы
Ионные каналы – интегральные белки, которые обеспечивают пассивный транспорт ионов по градиенту концентрации. Энергией для транспорта служит разность концентрации ионов по обе стороны мембраны (трансмембранный ионный градиент).
Неселективные каналы обладают следующими свойствами:
пропускают все типы ионов, но проницаемость для ионов K+ значительно выше, чем для других ионов;
всегда находятся в открытом состоянии.
Селективные каналы обладают следующими свойствами:
пропускают только один вид ионов; для каждого вида ионов существует свой вид каналов;
могут находиться в одном из 3 состояний: закрытом, активированном, инактивированном.
Избирательная проницаемость селективного канала обеспечивается селективным фильтром , который образован кольцом из отрицательно заряженных атомов кислорода, которое находится в самом узком месте канала.
Изменение состояния канала обеспечивается работой воротного механизма , который представлен двумя белковыми молекулами. Эти белковые молекулы, так называемые активационные ворота и инактивационные ворота, изменяя свою конформацию, могут перекрывать ионный канал.
В состоянии покоя активационные ворота закрыты, инактивационные ворота открыты (канал закрыт) (рис. 3). При действии на воротную систему сигнала активационные ворота открываются и начинается транспорт ионов через канал (канал активирован). При значительной деполяризации мембраны клетки инактивационные ворота закрываются и транспорт ионов прекращается (канал инактивирован). При восстановлении уровня МП канал возвращается в исходное (закрытое) состояние.
Рис. 3. Состояния селективного ионного канала и условия перехода между ними.
В зависимости от сигнала, который вызывает открытие активационных ворот, селективные ионные каналы подразделяют на:
хемочувствительные каналы – сигналом к открытию активационных ворот является изменение конформации ассоциированного с каналом белка-рецептора в результате присоединения к нему лиганда;
потенциалчувствительные каналы – сигналом к открытию активационных ворот является снижение МП (деполяризация) клеточной мембраны до определенного уровня, который называют критическим уровнем деполяризации (КУД).
Механизм формирования потенциала покоя
Мембранный потенциал покоя образуется главным образом благодаря выходу К+ из клетки через неселективные ионные каналы. Утечка из клетки положительно заряженных ионов приводит к тому, что внутренняя поверхность мембраны клетки заряжается отрицательно относительно наружной.
Мембранный потенциал, возникающий в результате утечки К+ , называют «равновесным калиевым потенциалом» (Ек). Его можно рассчитать по равнению Нернста
где R – универсальная газовая постоянная,
Т – температура (по Кельвину),
F – число Фарадея,
[К+] нар – концентрация ионов К+ снаружи клетки,
[К+] вн – концентрация ионов К+ внутри клетки.
ПП, как правило, очень близок к Ек, но не точно равен ему. Эта разница объясняется тем, что свой вклад в формирование ПП вносят:
поступление в клетку Na+ и Cl– через неселективные ионные каналы; при этом поступление в клетку Cl– дополнительно гиперполяризует мембрану, а поступление Na+ – дополнительно деполяризует ее; вклад этих ионов в формирование ПП невелик, так как проницаемость неселективных каналов для Cl– и Na + в 2,5 и 25 раза ниже, чем для К+ ;
прямой электрогенный эффект Na+ /К+ ионного насоса, возникающий в том случае, если ионный насос работает асимметрично (количество переносимых в клетку ионов K+ не равно количеству выносимых из клетки ионов Na+).
Механизм развития потенциала действия
В потенциале действия выделяют несколько фаз (рис. 4):
фаза деполяризации;
фаза быстрой реполяризации;
фаза медленной реполяризации (отрицательный следовый потенциал);
фаза гиперполяризации (положительный следовый потенциал).
Рис. 2.4. Изменение мембран-ного потенциала, интенсивности калиевого и натриевого трансмембранного тока и возбудимости клетки в разные фазы потенциала действия.
Д – фаза деполяризации, Рб – фаза быстрой реполяризации, Рм – фаза медленной реполяризации, Г – фаза гиперполяризации;
Н – период нормальной возбудимости, Ра – период абсолютной рефрактерности, Ро – период относительной рефрактерности, Н+ – период супернормальной возбудимости, Н- – период субнормальной возбудимости
Фаза деполяризации. Развитие ПД возможно только при действии раздражителей, которые вызывают деполяризацию клеточной мембраны. При деполяризации клеточной мембраны до критического уровня деполяризации (КУД) происходит лавинообразное открытие потенциалчувствительных Na+-каналов. Положительно заряженные ионы Na+ входят в клетку по градиенту концентрации (натриевый ток), в результате чего мембранный потенциал очень быстро уменьшается до 0, а затем приобретает положительное значение. Явление изменения знака мембранного потенциала называют реверсией заряда мембраны.
Фаза быстрой и медленной реполяризации . В результате деполяризации мембраны происходит открытие потенциалчувствительных К+ -каналов. Положительно заряженные ионы К+ выходят из клетки по градиенту концентрации (калиевый ток), что приводит к восстановлению потенциала мембраны. В начале фазы интенсивность калиевого тока высока и реполяризация происходит быстро, к концу фазы интенсивность калиевого тока снижается и реполяризация замедляется.
Фаза гиперполяризации развивается за счет остаточного калиевого тока и за счет прямого электрогенного эффекта активировавшейся Na+ / K+ помпы.
Овершут – период времени, в течение которого мембранный потенциал имеет положительное значение.
Пороговый потенциал – разность между мембранным потенциалом покоя и критическим уровнем деполяризации. Величина порогового потенциала определяет возбудимость клетки – чем больше пороговый потенциал, тем меньше возбудимость клетки.
Изменение возбудимости клетки при развитии возбуждения
Если принять уровень возбудимости клетки в состоянии физиологического покоя за норму, то в ходе развития цикла возбуждения можно наблюдать ее колебания. В зависимости от уровня возбудимости выделяют следующие состояния клетки (см. рис. 4).
Супернормальная возбудимость ( экзальтация ) – состояние клетки, в котором ее возбудимость выше нормальной. Супернормальная возбудимость наблюдается во время начальной деполяризации и во время фазы медленной реполяризации. Повышение возбудимости клетки в эти фазы ПД обусловлено снижением порогового потенциала по сравнению с нормой.
Абсолютная рефрактерность – состояние клетки, в котором ее возбудимость падает до нуля. Никакой, даже самый сильный, раздражитель не может вызвать дополнительного возбуждения клетки. Во время фазы деполяризации клетка невозбудима, поскольку все ее Na+ -каналы уже находятся в открытом состоянии.
Относительная рефрактерность – состояние, в котором возбудимость клетки значительно ниже нормальной; только очень сильные раздражители могут вызвать возбуждение клетки. Во время фазы реполяризации каналы возвращаются в закрытое состояние и возбудимость клетки постепенно восстанавливается.
Субнормальная возбудимость характеризуется незначительным снижением возбудимости клетки ниже нормального уровня. Это уменьшение возбудимости происходит вследствие возрастания порогового потенциала во время фазы гиперполяризации.