
Поверхность альвеолярного эпителия в лёгких покрыта сурфактантом, необходимым для обеспечения дыхания и адекватной иммунной защиты. Лёгочный сурфактант состоит из липидов (90%) и ряда белков, имеющих различные функции. Сурфактантные белки представлены белками SP-A, SP-D, SP-B и SP-C. В данном обзоре обсуждаются основные функции сурфактантных белков.
Ключевые слова
Обзор
Лёгкие выполняют две главные функции в организме: обеспечение дыхания и функционирование механизмов иммунной защиты. Корректное выполнение данных функций связано с лёгочным сурфактантом.
Сурфактант в лёгких синтезируется альвеолярными клетками II типа и секретируется в альвеолярное пространство. Сурфактант покрывает поверхность альвеолярного эпителия и состоит из липидов (90 %) и белков (10 %), составляя липопротеидный комплекс. Липиды представлены в основном фосфолипидами. Дефицит и/или качественные изменения состава легочного сурфактанта описаны при туберкулезе, респираторном дистресс-синдроме новорожденных, пневмонии и других заболеваниях. 1.
Сурфактантные белки представлены белками SP-A, (Surfactant Protein A, 5,3%), SP-D (0,6 %), SP-B (0,7 %), и SP-C (0,4%). 4.
Функции гидрофильных белков SP-A и SP-D связаны с иммунной защитой в легких. Эти белки связывают липополисахарид грамотрицательных бактерий и агрегируют различные микроорганизмы, влияют на активность тучных, дендритных клеток, лимфоцитов и альвеолярных макрофагов. SP-A ингибирует созревание дендритных клеток, тогда как SP-D увеличивает способность альвеолярных макрофагов к захвату и презентации антигенов, стимулируя адаптивный иммунитет [7,8].
Сурфактантный белок А является наиболее обильным белком лёгочного сурфактанта. Он обладает выраженными иммуномодулирующими свойствами. Белок SP-A воздействует на рост и жизнеспособность микроорганизмов, повышая проницаемость их цитоплазматической мембраны. Более того, SP-A стимулирует хемотаксис макрофагов, влияет на пролиферацию клеток иммунного ответа и на продукцию цитокинов, повышает продукцию реактивных оксидантов, повышает фагоцитоз клеток, подвергшихся апоптозу и стимулирует фагоцитоз бактерий. SP-A человека состоит из двух генных продуктов — SP-A1 и SP-A2, структура и функция которых различна. Наиболее важное различие в структуре SP-A1 и SP-A2- аминокислотная позиция 85 коллагеноподобного региона белка SP-A, где SP-A1 имеет цистеин, а SP-A2 – аргинин. Функциональные различия между SP-A1 и SP-A2 включают их способность стимулировать фагоцитоз, ингибировать секрецию сурфактанта.[7]. Во всех этих случаях SP-A2 обладает большей активностью, чем SP-A1. [8].
Функции гидрофобных белков SP-B и SP-C связаны с обеспечением возможности дыхания. Они снижают поверхностное натяжение в альвеолах и способствуют равномерному распределению сурфактанта на поверхности альвеол. [9].
Как называется белок транспортирующий кислород из легочных альвеол в ткани
После поступления свежего воздуха в альвеолы начинается следующий этап дыхательного процесса: диффузия кислорода из альвеол в кровь и диффузия двуокиси углерода в обратном направлении — из крови в альвеолы. Процесс диффузии представляет собой беспорядочное движение молекул, прокладывающих себе дорогу через дыхательную мембрану и жидкости во всех направлениях. Однако в физиологии дыхания нас интересуют не только основные механизмы диффузии, но и ее скорость, что представляет собой намного более сложную проблему и потребует более глубоких знаний в области физики диффузии и обмена газов.
Физические основы диффузии и парциальные давления газов
Все газы, представляющие интерес для физиологии дыхания, являются простыми молекулами, которые свободно перемещаются в смеси. Этот процесс называют диффузией. Это справедливо и для газов, растворенных в жидкостях и тканях тела.
Для процесса диффузии необходимо наличие источника энергии. Энергия производится кинетическим движением самих молекул. При температуре выше абсолютного нуля молекулы находятся в постоянном движении. Это значит, что свободные молекулы, не связанные с другими молекулами, двигаются линейно на высокой скорости до встречи с другими молекулами. После столкновения их движение получит новое направление — до следующего столкновения. Таким образом, молекулы находятся в быстром и случайном движении среди себе подобных.
Читайте также: Все части листа состоят из клеток тканей да или нет
а) Диффузия газа одном направлении. Влияние градиента концентрации. Если в емкости или в растворе концентрация одного газа в одной зоне высокая, а в другой — низкая (для облегчения понимания просим вас изучить рисунок ниже), то суммарная диффузия газа будет направлена от зоны с высокой концентрацией в зону с низкой концентрацией: на рисунке в зоне А находится больше молекул, способных двигаться в направлении зоны Б, чем молекул, которые могут переместиться в обратном направлении, поэтому диффузия в каждом из направлений пропорциональна концентрации молекул, что на рисунке демонстрирует длина стрелок.
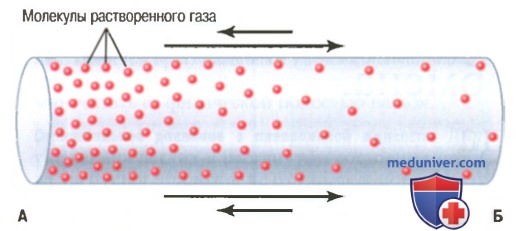
Диффузия кислорода из одной зоны (А) в другую (Б). Разница в длине стрелок представляет величину конечной диффузии
б) Давление газов в газовой смеси. Парциальные давления отдельных газов. Давление создается множественными ударами движущихся молекул о поверхность, поэтому давление газа на поверхности дыхательных ходов и альвеол пропорционально суммарной силе ударов о поверхность всех молекул данного газа в данный момент, т.е. давление газа прямо пропорционально концентрации молекул газа.
В физиологии дыхания мы имеем дело со смесями газов, состоящих главным образом из кислорода, азота и двуокиси углерода. Скорость диффузии каждого из них прямо пропорциональна давлению, создаваемому только этим газом, и это давление называют парциальным давлением данного газа. Далее приводим объяснение концепции парциального давления.
Воздух состоит примерно из 79% азота и 21% кислорода. Общее давление этой смеси на уровне моря равно 760 мм рт. ст. Из приведенного ранее объяснения молекулярных основ возникновения давления ясно, что доля каждого газа в давлении их смеси находится в прямой пропорции с его концентрацией, поэтому 79% из 760 мм рт. ст. давления воздуха создается азотом (600 мм рт. ст.) и 21% — кислородом (160 мм рт. ст.). Таким образом, парциальное давление азота в смеси составляет 600 мм рт. ст., парциальное давление кислорода — 160 мм рт.ст., а общее давление (760 мм рт. ст.) является суммой отдельных парциальных давлений. Парциальное давление отдельных газов обозначают PCO2, PO2, PN2, PH2O, PHe и т.д.
Видео физиология газообмена в легких и транспорта газов кровью — профессор, д.м.н. П.Е. Умрюхин
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Как называется белок транспортирующий кислород из легочных альвеол в ткани
После диффузии кислорода из альвеол в капиллярную кровь его дальнейший транспорт в капилляры периферических тканей совершается почти полностью в связанной с гемоглобином форме. Наличие в эритроцитах гемоглобина позволяет крови транспортировать в 30-100 раз больше кислорода, чем могло бы транспортироваться в виде газа, растворенного в водной составляющей крови.
В клетках тканей тела кислород реагирует с разными веществами, формируя большое количество двуокиси углерода, который потом входит в капилляры ткани и транспортируется обратно в легкие. Двуокись углерода также связывается с разными химическими веществами, находящимися в крови, что увеличивает транспорт двуокиси углерода в 15-20 раз.
В статьях на сайте представлены физические и химические принципы транспорта кислорода и двуокиси углерода в крови и тканевой жидкости как с количественной, так и качественной стороны.
а) Транспорт кислорода из легких в ткани тела. Газы могут переместиться из одной точки в другую путем диффузии и причиной такого передвижения всегда является наличие градиента парциального давления между этими точками. Так, кислород диффундирует в легких из альвеол в капиллярную кровь, потому что парциальное давление кислорода (Рог) в альвеолах больше, чем в крови легочных капилляров. В других тканях тела PO2 в капиллярной крови выше, чем в тканях, и это заставляет кислород диффундировать в ткани.
Читайте также: Птицы из ткани с перьями
В метаболических процессах клеток кислород используется для образования двуокиси углерода, в результате внутриклеточное давление двуокиси углерода (PCO2) поднимается до высоких значений, что приводит к диффузии двуокиси углерода в тканевые капилляры. Когда кровь доходит до легких, двуокись углерода диффундирует из крови в альвеолы, т.к. Рсог в крови легочных капилляров выше, чем в альвеолах. Таким образом, транспорт кислорода и двуокиси углерода кровью зависит как от диффузии, так и от кровотока. Далее рассмотрим количественную сторону факторов, определяющих эти явления.
![]()
Поглощение кислорода капиллярной кровью в легких
б) Диффузия кислорода из альвеол в капиллярную кровь легких. В верхней части рисунка выше изображена альвеола, расположенная рядом с легочным капилляром, и показана диффузия молекул кислорода из альвеолярного воздуха в кровь. PO2 в альвеолярной газовой смеси составляет 104 мм рт. ст., а PO2 в венозной крови, входящей в легочный капилляр через его артериальный конец, составляет только 40 мм рт. ст., т.к. большое количество кислорода было поглощено из крови во время прохождения ее через периферические ткани. Таким образом, начальная разница в парциальном давлении, являющаяся причиной диффузии кислорода в легочные капилляры, составляет 104 — 40, или 64 мм рт. ст. На графике в нижней части рисунка виден резкий подъем PO2 крови во время прохождения ее через капилляр; к моменту прохождения 1/3 длины капилляра Р02 в крови составляет около 104 мм рт. ст., т.е. почти достигает Р02 в альвеолярном воздухе.
Поглощение кислорода кровью в легких во время физической нагрузки. При тяжелой физической нагрузке потребление кислорода может оказаться в 20 раз выше нормы. При этом из-за повышения сердечного выброса при такой нагрузке время прохождения легочного капилляра кровью может сократиться более чем в 2 раза. Однако в силу существования большого фактора надежности для диффузии кислорода через легочную мембрану кровь ко времени выхода из капилляра все же насыщается кислородом почти до максимального уровня. Это объясняется следующим.
Во-первых, во время физической нагрузки диффузионный объем кислорода возрастает почти в 3 раза. Это происходит главным образом из-за увеличения площади поверхности капилляров, участвующих в процессе диффузии, а также из-за приближения вентиляционно-перфузионного коэффициента в верхних частях легких к идеальной величине.
Во-вторых, на рисунке выше показано, что при отсутствии физической нагрузки кровь достигает почти полного насыщения кислородом уже после прохождения первой трети легочного капилляра и во время прохождения следующих двух третей обычно в нее добавляется очень мало кислорода. Можно сказать, что в покое кровь остается в легочных капиллярах в 3 раза дольше, чем это необходимо для полного насыщения ее кислородом, поэтому во время физической нагрузки кровь может полностью или почти полностью насыщаться кислородом и после сокращения времени пребывания в капиллярах.
Видео физиология газообмена в легких и транспорта газов кровью — профессор, д.м.н. П.Е. Умрюхин
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Как называется белок транспортирующий кислород из легочных альвеол в ткани
Кровообращение выполняет одну из важнейших функций переноса кислорода от легких к тканям, а углекислого газа — от тканей к легким. Потребление кислорода клетками тканей может изменяться в значительных пределах, например при переходе от состояния покоя к физической нагрузке и наоборот. В связи с этим кровь должна обладать большими резервами, необходимыми для увеличения ее способности переносить кислород от легких к тканям, а углекислый газ в обратном направлении.
Транспорт кислорода.
При 37 С растворимость 02 в жидкости составляет 0,225 мл • л-1 • кПа-1 (0,03 мл/л/мм рт. ст.). В условиях нормального парциального давления кислорода в альвеолярном воздухе, т. е. 13,3 кПа или 100 мм рт.ст., 1 л плазмы крови может переносить только 3 мл 02, что недостаточно для жизнедеятельности организма в целом. В покое в организме человека за минуту потребляется примерно 250 мл кислорода. Чтобы тканям получить такое количество кислорода в физически растворенном состоянии, сердце должно перекачивать за минуту огромное количество крови. В эволюции живых существ проблема транспорта кислорода была более эффективно решена за счет обратимой химической реакции с гемоглобином эритроцитов. Кислород переносится кровью от легких к тканям организма молекулами гемоглобина, которые содержатся в эритроцитах.
Читайте также: Ткань для мешка гост 30090 93
Гемоглобин способен захватывать кислород из альвеолярного воздуха (соединение называется ок-сигемоглобином) и освобождать необходимое количество кислорода в тканях. Особенностью химической реакции кислорода с гемоглобином является то, что количество связанного кислорода ограничено количеством молекул гемоглобина в эритроцитах крови. Молекула гемоглобина имеет 4 места связывания с кислородом, которые взаимодействуют таким образом, что зависимость между парциальным давлением кислорода и количеством переносимого кислорода с кровью имеет S-образную форму, которая носит название кривой насыщения или диссоциации оксигемоглобина (рис. 10.18). При парциальном давлении кислорода 10 мм рт. ст. насыщение гемоглобина кислородом составляет примерно 10 %, а при Р02 30 мм рт. ст. — 50—60 %. При дальнейшем увеличении парциального давления кислорода от 40 мм рт. ст. до 60 мм рт. ст. происходит уменьшение крутизны кривой диссоциации оксигемоглобина и процент его насыщения кислородом возрастает в диапазоне от 70—75 до 90 % соответственно. Затем кривая диссоциации оксигемоглобина начинает занимать практически горизонтальное положение, поскольку увеличение парциального давления кислорода с 60 до 80 мм рт. ст. вызывает прирост насыщения гемоглобина кислородом на 6 %. В диапазоне от 80 до 100 мм рт. ст. процент образования оксигемоглобина составляет порядка 2. В результате кривая диссоциации оксигемоглобина переходит в горизонтальную линию и процент насыщения гемоглобина кислородом достигает предела, т. е. 100. Насыщение гемоглобина кислородом под влиянием Р02 характеризует своеобразный молекулярный «аппетит» этого соединения к кислороду.
Значительная крутизна кривой насыщения гемоглобина кислородом в диапазоне парциального давления от 20 до 40 мм рт. ст. способствует тому, что в ткани организма значительное количество кислорода может диффундировать из крови в условиях фадиента его парциального давления между кровью и клетками тканей (не менее 20 мм рт. ст.). Незначительный процент насыщения гемоглобина кислородом в диапазоне его парциального давления от 80 до 100 мм рт. ст. способствует тому, что человек без риска снижения насыщения артериальной крови кислородом может перемещаться в диапазоне высот над уровнем моря до 2000 м.
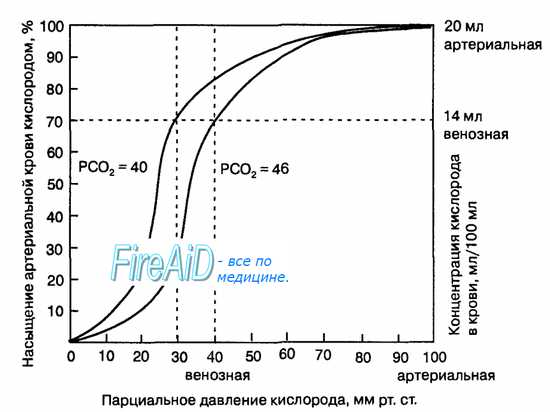
Рис. 10.18. Кривая диссоциации оксигемоглобина. Пределы колебания кривой при РС02 = 40 мм рт. ст. (артериальная кровь) и РС02 = 46 мм рт. ст. (венозная кровь) показывают изменение сродства гемоглобина к кислороду (эффект Ходена).
Общие запасы кислорода в организме обусловлены его количеством, находящимся в связанном состоянии с ионами Fe2+ в составе органических молекул гемоглобина эритроцитов и миоглобина мышечных клеток.
Один грамм гемоглобина связывает 1,34 мл 02. Поэтому в норме при концентрации гемоглобина 150 г/л каждые 100 мл крови могут переносить 20,0 мл 02.
Количество 02, которое может связаться с гемоглобином эритроцитов крови при насыщении 100 % его количества, называется кислородной емкостью гемоглобина. Другим показателем дыхательной функции крови является содержание 02 в крови (кислородная емкость крови), которое отражает его истинное количество, как связанного с гемоглобином, так и физически растворенного в плазме. Поскольку в норме артериальная кровь насыщена кислородом на 97 %, то в 100 мл артериальной крови содержится примерно 19,4 мл 02.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом