
С тех пор процессы пролиферации и дифференцировки клеток ушли далеко вперед, создав настоящее чудо — вас, человека. У растения жизнь начинается точно так же — с одной маленькой клетки, из которой в дальнейшем будут развиваться ткани и органы самых разных форм. Главная заслуга роста растения принадлежит образовательной ткани.
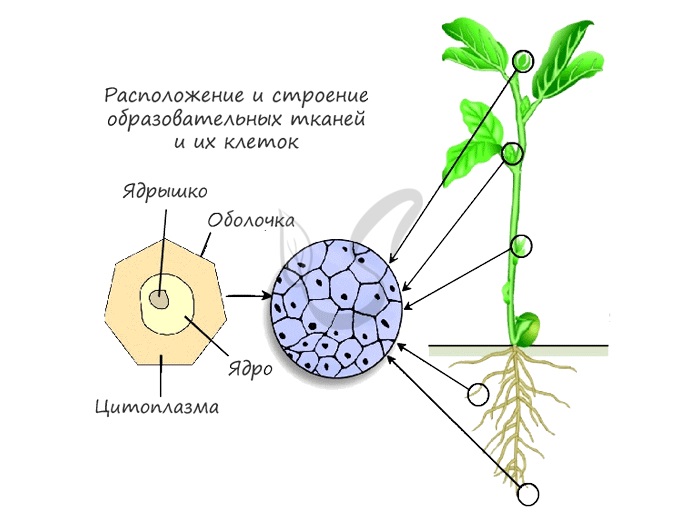
Как вы видите, на картинке схематично изображено месторасположение образовательной ткани. Главным образом это:
- Кончик побега — конус нарастания в почках
- Кончик корня — зона деления, прикрытая корневым чехликом для защиты
- Камбий — обеспечивает рост растения в ширину
- Основание междоузлий и черешков листьев — это также зоны активного роста растения
Именно в этих местах и происходит деление клеток и рост растения. Важно отметить, что сезонные изменения активности клеток камбия являются причиной возникновения годичных колец древесины. Внешний вид годичных колец обусловлен хронологической закономерностью: весной больше образуется проводящей ткани (более тонкая и рыхлая внутри), а осенью — механическая (толстая, более твердая). Именно поэтому годичные кольца на спиле дерева выглядят как чередование колец, отличающихся друг от друга.
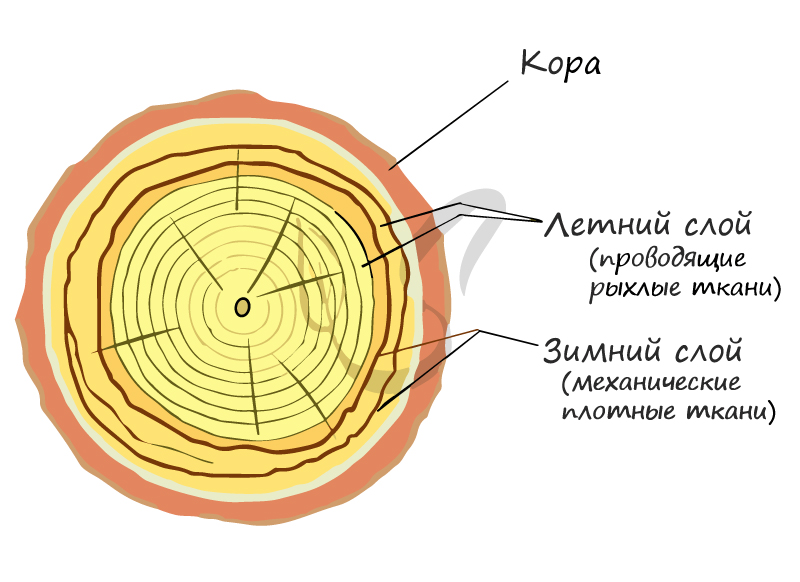
На внешний вид годичных колец оказывают весьма сильное влияние условия внешней среды. Так, при дефиците трофического компонента (питательных веществ), к примеру, у растений, растущих на болоте, годичные кольца выглядят тоньше своих обычных размеров.
Ветер также оказывает существенное влияние: при его постоянном действии происходит перераспределение древесины по стволу. Оказывая действие на крону, ветер смещает центр тяжести дерева, что сказывается на его нижележащих отделах. Они начинают компенсаторно утолщаться для предотвращения слома дерева. При постоянно дующем ветре ствол сильно искривляется, а форма кроны становится флагообразной.

Тема камбия и форм стволов растений весьма занимательна, и все-таки мы должны разобраться в строении самой образовательной ткани. Она представлена живыми мелкими быстро делящимися клетками с относительно крупным ядром. Объем цитоплазмы небольшой, она вязкая по консистенции, оболочка клетки тонкая. Это уязвимые клетки, которые растение оберегает по-своему, подобно тому, как животные оберегают только что появившееся потомство.

Другое название образовательных тканей — меристемы (с др.-греч. — «μεριστός» — делимый). По времени возникновения различают первичные и вторичные меристемы.
Первичные меристемы — закладываются в эмбриогенезе
1) Вставочные меристемы (интеркалярные) — в виде отдельных участков в зоне активного роста в разных частях растения. Такие ткани можно найти в основании междоузлий у злаков, черешков листьев у многих растений. У злаковых наблюдается быстрый рост стебля за счет множественного расположения данной ткани на стебле — «вставочный рост».
2) Прокамбий — основа будущего камбия, перицикла, окружающего проводящие ткани в один или несколько слоёв (у голосеменных). В корнях перицикл является корнеродным слоем, так как в корне с него начинается формирование осевого цилиндра, наружным слоем которого он является. В нём закладываются придаточные и боковые корни, что имеет принципиальное значение для формирования корневой системы растения.
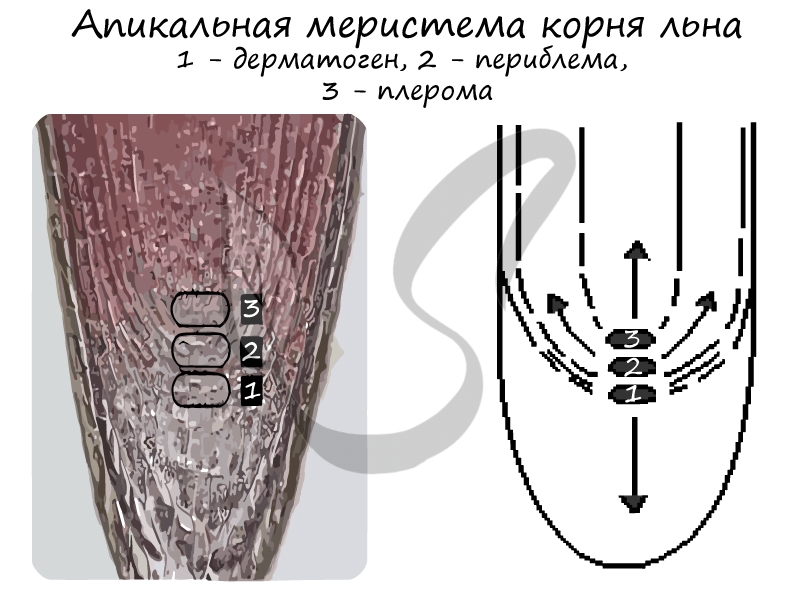
3) Верхушечные (апикальные) — формируются на верхушках стеблей и кончиках корней. В периферической части корня различают три слоя:
- Дерматоген — в дальнейшем преобразующийся в первичную покровно-всасывающую ризодерму (эпиблему или ризодерму)
- Периблема — образующая ткани первичной коры
- Плерома — внутренний слой ткани центрального осевого цилиндра
Вторичные меристемы — закладываются в постэмбриональном развитии
Камбий и феллоген (пробковый камбий) — занимают боковое положение по отношению к оси органа, обеспечивают рост вширь. Растения часто повреждаются, их задевают животные, нарушая целостность тканей и органов. На этот случай в группе вторичных меристем есть раневые меристемы, дающие начало защитной ткани в местах повреждения растения.

Топографическая классификация меристем
Спешу заверить, это отнюдь не сложная классификация, которой нужно бояться. Речь пойдет о взгляде на те же образовательные ткани с другой стороны. В переводе с греч. τόπος — место. Мы рассмотрим меристемы в соответствии с их месторасположением на растении.
- Верхушечная или апикальная (лат. apex — вершина) — расположена на кончике корня и конусе нарастания побега
- Боковая или латеральная (лат. latus — бок): камбий – обеспечивает рост стебля и корня в толщину
- Краевая или маргинальная (лат. margo — край) меристема даёт начало листовой пластинке
- Вставочная или интеркалярная (лат. inter — между и calaris — вставочный, добавочный) — расположена преимущественно у основания стеблевых междоузлий между зонами дифференцированных тканей.
Читайте также: Характеристики элементов проводящей ткани
© Беллевич Юрий Сергеевич 2018-2022
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Образовательные ткани (меристемы)
Меристема(образовательная ткань) обусловливает рост растений в длину и толщину в связи с делением клеток и их дифференциацией. Различают первичную и вторичную меристемы.
Первичная меристема появляется в самом начале роста проростка из клеток зародыша в виде конуса нарастания стебля и корня.
В первичной меристеме закладывается прокамбий. При этом клетки меристемы делятся преимущественно продольными перегородками, концы их заостряются. Так возникает прокамбиальный тяж, состоящий из прозенхимных меристематических клеток, из которых впоследствии дифференцируются проводящие и механические ткани или вторичная меристема – камбий.
Вторичной называют меристему, возникшую из какой-либо уже дифференцированной ткани.
Ткани, которые образуются из первичной меристемы, называют первичными, а из вторичной меристемы – вторичными.
Итак, рост растений обеспечивают меристемы в связи с делением клеток. Митоз – основной способ деления соматических клеток.
Митоз является составной частью митотического цикла, который проходит каждая делящаяся клетка. Митотический цикл состоит из интерфазы и митоза, тесно связанных между собой.
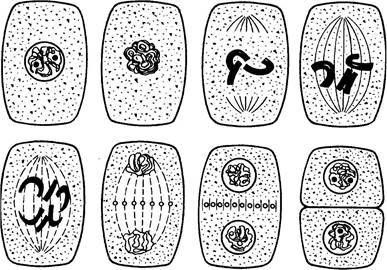 |
Рис. 3.1. Схема деления клетки
Интерфаза – наиболее продолжительная часть митотического цикла. В этой фазе происходят важные биохимические процессы, подготавливающие клетку к делению, – редупликация ДНК, накопление веществ и энергии. В интерфазе различают 3 периода: предсинтетический – рост и подготовка к удвоению ДНК, синтетический – синтез ДНК и постсинтетический – подготовка к построению веретена и накопление энергии.
Митоз в свою очередь делят на 4 фазы: профазу, метафазу, анафазу и телофазу. В профазе начинается спирализация хромосом: они становятся более короткими, увеличивается их толщина. Ядрышко, хорошо видимое в начале профазы, к ее концу исчезает. Подвергается деструкции ядерная оболочка и хромосомы оказываются в цитоплазме. К концу профазы начинает формироваться по полюсам клетки ахроматиновое веретено. Метафаза характеризуется наличием ахроматинового веретена. Часть его нитей связаны с центромерами хромосом. К центромере одной хромосомы подходит по одной нити от каждого полюса. Хромосомы выстраиваются в экваториальной плоскости клетки, там, где произойдет ее разделение на две дочерние клетки. Каждая хромосома состоит из двух хроматид. В метафазе между ними появляется щель и хроматиды слегка обособляются друг от друга, но остаются соединенными в области центромеры. В анафазе сестринские хроматиды полностью разъединяются и расходятся к полюсам клетки. На каждом полюсе оказывается по равноценному набору дочерних хромосом. Каждая из них состоит из одной хроматиды. В телофазе осуществляется реконструкция ядра в дочерних клетках. Хромосомы деспирализуются, образуется ядрышко, восстанавливается ядерная оболочка. Между двумя формирующимися дочерними клетками распределяется все живое содержимое материнской клетки. Появляется межклеточная пластинка в экваториальной плоскости. Каждая дочерняя клетка достраивает недостающую оболочку.
После четырех фаз деления начинается интерфаза – перерыв между следующими друг за другом митотическими делениями ядра. В этот период осуществляется рост клетки, сопровождающийся увеличением цитоплазмы и накоплением пластического и энергетического материала, необходимого для обеспечения последующего деления клетки. В интерфазу в клетке, готовой перейти к очередному делению, происходит удвоение количества наследственного материала. В результате каждая хромосома вновь состоит из двух хроматид. Только после этого ядро снова может делиться.
Комплекс взаимосвязанных и согласованных во времени процессов, происходящих при подготовке клетки к митотическому делению (в интерфазе) и на протяжении самого деления, называют митотическим циклом.
Образовательные ткани. Процесс деления клетки.
1. Митоз и его значение для растения.
2. Профаза. 3. Метафаза. 4. Анафаза. 5. Телофаза.
6. Понятие о тканях. Общие особенности тканей растений.
7. Меристемы, их цитологические особенности и значение.
Обычно деление клетки, как у животных, так и у растений, происходит путём сложного деления, называемого митозом.
Читайте также: Ткань для гобеленовой вышивки
Митоз – основной способ деления эукариотических клеток. Он подразделяется на ряд фаз: профаза, метафаза, анафаза, телофаза.
Профаза (2n4с). Это самая продолжительная фаза митоза. Её началом принято считать момент, когда заканчивается спирализация хромосом в ядре. В это время хромосомы становятся различимы в световой микроскоп. Становится заметно, что каждая хромосома состоит из двух хроматид, соединённых первичной перетяжкой, или центромерой. Затем разрушается ядерная оболочка и исчезает ядрышко.
Метафаза (2n4с). Хромосомы выстраиваются на экваторе клетки совершенно случайным образом, образуя метафазную, или экваториальную, пластинку. Хорошо видно, что каждая хромосома состоит из двух сестринских хроматид, слегка обособленных друг от друга по длине хромосомы, но соединённых в области центромеры. От хромосом тянутся плазматические нити к двум точкам, расположенных в противоположных частях клетки. Эти точки называются полюсами клетки.
Анафаза (2n4с). Центромеры хромосом разрушаются, и хроматиды, составляющие хромосому, становятся дочерними хромосомами. Все дочерние хромосомы одновременно расходятся к полюсам клетки.
Телофаза – последняя стадия митоза (2n4с®2n2с). Все процессы в телофазе напоминают прокручивающуюся назад профазу: хромосомы деспирализуются, формируется ядерная оболочка, появляются ядрышки. Затем наступает цитокинез – деление цитоплазмы. У растительных клеток она делится от периферии к центру.
В результате митоза из одной клетки образуются две диплоидные клетки совершенно одинаковые по содержанию наследственного материала, а значит, имеющие одинаковое строение и выполняющие одинаковые функции.
Период между предыдущим и последующим делением клетки называется интерфазой. В это время клетка самообновляется, в ней происходит активный обмен веществ, накопление белков, а главное, синтез ДНК. Именно в интерфазе количество ДНК увеличивается в два раза (2n4с®2n2с). Это позволит дочерним клеткам получить одинаковую наследственную информацию.
Появление тканей у высших растений – крупнейший ароморфоз, позволивший растениям адаптироваться к обитанию в более жёсткой, чем вода, внешней среде.
Ткани растений не столь специализированы как ткани животных, т. е. они многофункциональны и могут состоять из элементов, имеющих разное строение.
В ходе онтогенеза ткани могут давать начало другим тканям или перерождаться. Это объясняется колоссальной способностью растительных тканей к регенерации.
В растении в зависимости от степени дифференцировки выделяют два вида тканей: образовательные ткани (меристемы) и постоянные ткани. Клетки постоянных тканей достигли окончательной дифференцировки. К ним относятся проводящие, механические, покровные, основные и выделительные ткани.
Ткани, состоящие из одного типа клеток, получили название простых, а состоящие из разных типов – сложных, или комплексных.
Меристемы занимают ничтожный объём в теле растения – 0,1% общей массы. Инициальные клетки меристем задерживаются на эмбриональной стадии развития в течение всей жизни растения, а их производные постепенно дифференцируются и превращаются в клетки различных постоянных тканей.
Меристемы присущи только растениям. Они обеспечивают обновление и рост всего организма в ходе всего онтогенеза. В этой связи возраст разных тканей и органов в пределах одного организма неодинаков. Эта особенность растений (рост в течение всей жизни) обусловлена их прикреплённым образом жизни и автотрофным типом питания.
Клетки меристем очень слабо дифференцированы, поэтому им присущи следующие черты:
1) клетки мелкие и плотно упакованные (межклетников нет)
2) клеточные стенки тонкие первичные
3) цитоплазма вязкая, в ней активно идут процессы биосинтеза белка и дыхания
4) практически все органоиды слабо дифференцированы
5) нет единой центральной вакуоли, вакуолярная система представлена группой мелких пузырьков
6) клетки способны к многократным делениям
В меристеме можно выделить две группы клеток:
- инициальные клетки – небольшая группа клеток (у высших растений 8-10), способных в течение всей жизни растения делиться митозом. Для растений, обитающих в условиях сезонного климата, инициальные клетки находятся либо в состоянии активной жизнедеятельности (в вегетационный период), либо в состоянии покоя (неблагоприятный период).
- производные клетки – образовались от инициальных, делятся митозом 3-5 раз, а затем эти клетки переходят к процессу растяжения и дифференцировки, постепенно превращаясь в постоянные ткани.
Топографическая классификация меристем
- апикальные (верхушечные) меристемы располагаются на верхушках побегов и корней, обеспечивая нарастание их в длину.
Вакуоли многочисленные, мелкие, но под световым микроскопом обычно заметны. Пластид и митохондрий мало, и они мелкие. Эргастические вещества отсутствуют.
Если растение теряет апикальную меристему, то рост органа в длину прекращается, но это может стимулировать рост боковых органов.
- вставочные (интеркалярные) расположены у снования междоузлий побегов, у основания тычинок, в основании листьев злаков.
- боковые (латеральные) располагаются параллельно боковым поверхностям осевых органов (стеблей и корней) и обеспечивают их утолщение. К боковым меристемам относят прокамбий, камбий, перицикл, феллоген.
Читайте также: Как почистить утюг от пригоревшей ткани в домашних карандашом
Латеральные меристемы располагаются параллельно боковым поверхностям осевых органов, нередко образуя цилиндры, на поперечных срезах имеющих вид колец. Гланейшие латеральные меристемы – камбий и феллоген. Эти меристемы обеспечивают нарастание стеблей и стволов в толщину, образуя вторичные ткани и формируя вторичное тело растения. Камбий даёт начало вторичным производным тканям – вторичной ксилеме и флоэме, а феллоген – перидерме.
Клетки боковых меристем различны по величине и форме. Они примерно соответствуют клеткам тех постоянных тканей, которые из них в дальнейшем возникают. Так, в камбии встречаются как паренхимные, так и прозенхимные инициали. Из паренхимных инициалей возникают паренхимные элементы проводящих тканей, а из прозенхимных – проводящие элементы.
- раневые меристемы образуются в местах повреждения тканей и органов и дают начало каллусу. Это особая ткань, состоящая из однородных паренхимных клеток, прикрывающих место поражения.
Классификация меристем по происхождению
Первичная меристема – возникает на самых первых этапах онтогенеза растений, берут своё начало от делящейся зиготы. К ним относятся апикальные меристемы, прокамбий, и интеркалярные меристемы. Апикальные меристемы функционируют в течение всей жизни растения, все остальные меристемы живут определённый промежуток времени и в итоге полностью превращаются в постоянные ткани.
Вторичные меристемы – формируются на более поздних этапах онтогенеза, чем первичные. Они возникают частично из первичных меристем, а частично из постоянных тканей. К ним относятся камбий, феллоген и раневые меристемы.
Способы закладывания межклеточной перегородки.
Если межклеточная перегородка закладывается под прямым углом к поверхности органа – это антиклинальное деление клетки. При этом образуются структуры пластинчатого типа (листовые пластинки цветковых растений).
Если межклеточная перегородка закладывается параллельно ближайшей поверхности органа – перериклинальное деление. Оно характерно для латеральных меристем, за счёт таких делений происходит утолщение осевых органов.
Если межклеточная перегородка закладывается касательно окружности – это тангециальное деление клетки. Клеточные деления при этом происходят, по сути, во всех плоскостях, и в результате образуется массив меристемы, напоминающий шар. Так формируется эндосперм семян.
Типы роста клеток меристем в процессе дифференцировки
Симпластический – рост оболочек смежных клеток происходит согласованно и их связь через плазмодесмы не нарушается (характерен для органов, удлиняющихся в период первичного роста).
Интрузивный (интерпозиционный) – рост отдельных клеток не согласован, одни клетки могут внедряться в пространство, образовавшееся между другими клетками (камбиальные инициали, склеренхимные волокна, трахеиды).
Ткани – это системы клеток, структурно и функционально сходные друг с другом и обычно имеющие общее происхождение. Тканей нет только у печёночных мхов.
Большинство клеток зародыша обладают меристематической активностью, т. е. способностью к делению. Часть этих клеток сохраняет эту способность в течение всей жизни. Это инициальные клетки, или инициали.
Клетки, постоянно возникающие в процессе деления инициалей, называются производными. Производные клетки делятся обычно один или два раза и начинают дифференцироваться. Дифференцировка касается, прежде всего, качественных особенностей клеток, но происходит на фоне различных количественных процессов, в частности роста. В результате меристемы дают начало постоянным тканям, клетки которых лишены способности к делению. Как правило, дифференцировка не обратима, и только когда специализация зашла не слишком далеко, клетки постоянных тканей могут вновь приобретать меристематическую активность. На ход дифференцировки оказывают влияние фитогормоны.
Внутреннее строение листа формируется только за счёт первичных меристем, все ткани листа первичные.
Основную массу листа, располагающуюся между верхней и нижней эпидермой, занимает хлоренхима, которая в листе получила особое название – мезофилл.
Цитологические особенности мезофилла: клеточные стенки всегда тонкие (либо первичные, либо с незначительными вторичными изменениями), наличие плазмодесм, наличие хлоропластов (их количество и размеры зависят от освещенности).
Система разветвлённых проводящих пучков снабжает мезофилл водой и растворами минеральных солей, а также обеспечивает отток органических веществ, образовавшихся в процессе фотосинтеза.
Поможем написать любую работу на аналогичную тему
Образовательные ткани. Процесс деления клетки.
Образовательные ткани. Процесс деления клетки.