
Кнооп в 1904 г. выдвинул гипотезу β-окисления жирных кислот на основании опытов по скармливанию кроликам различных жирных кислот, в которых один атом водорода в концевой метальной группе (у ω-углеродного атома) был замещен фенильным радикалом (С6Н5-).
Кнооп высказал предположение, что окисление молекулы жирной кислоты в тканях организма происходит в β-положении; в результате происходит последовательное отсечение от молекулы жирной кислоты двууглеродных фрагментов со стороны карбоксильной группы.
Жирные кислоты, входящие в состав естественных жиров животных и растений, принадлежат к ряду с четным числом углеродных атомов. Любая такая кислота, отщепляя по паре углеродных атомов, в конце концов проходит через стадию масляной кислоты, которая после очередного β-окисления должна дать ацетоуксусную кислоту. Последняя затем гидролизуется до двух молекул уксусной кислоты.
Теория β-окисления жирных кислот, предложенная Кноопом, не потеряла своего значения и до настоящего времени и является в значительной мере основой современных представлений о механизме окисления жирных кислот.
Современные представления об окислении жирных кислот
Установлено, что окисление жирных кислот в клетках происходит в митохондриях при участии мультиферментного комплекса. Известно также, что жирные кислоты первоначально активируются при участии АТФ и HS-KoA; субстратами на всех последующих стадиях ферментативного окисления жирных кислот служат КоА-эфиры этих кислот; выяснена также роль карнитина в транспорте жирных кислот из цитоплазмы в митохондрии.
Процесс окисления жирных кислот складывается из следующих основных этапов.
Активация жирных кислот и их проникновение из цитоплазмы в митохондрии. Образование «активной формы» жирной кислоты (ацил-КоА) из коэнзима А и жирной кислоты является эндергоническим порцессом протекающим за счет использования энергии АТФ:

Реакция катализируется ацил-КоА-синтетазой. Существует несколько таких ферментов: один из них катализирует активацию жирных кислот, содержащих от 2 до 3 углеродных атомов, другой- от 4 до 12 атомов, третий — от 12 и более атомов углерода.
Как уже отмечалось, окисление жирных кислот (ацил-КоА) происходит в митохондриях. В последние годы было показано, что способность ацил-КоА проникать из цитоплазмы в митохондрии резко возрастает в присутствии азотистого основания — карнитина (γ-триметиламино-β-гидроксибутирата). Ацил-КоА, соединяясь с карнитином, при участии специфического цитоплазматического фермента (карнитин-ацил-КоА-трансферазы) образует ацилкарнитин (эфир карнитина и жирной кислоты), который обладает способностью проникать внутрь митохондрии:

После прохождения ацилкарнитина через мембрану митохондрии происходит обратная реакция — расщепление ацилкарнитина при участии HS-KoA и митохондриальной карнитин-ацил-КоА-трансферазы:

При этом карнитин возвращается в цитоплазму клетки, а ацил-КоА подвергается в митохондриях окислению.
Первая стадия дегидрирования. Ацил-КоА в митохондриях прежде всего подвергается ферментативному дегидрированию;
при этом ацил-КоА теряет два атома водорода в α- и β-положении, превращаясь в КоА-эфир ненасыщенной кислоты:

По-видимому, существует несколько ФАД-содержащих ацил-КоА-дегидрогеназ, каждая из которых обладает специфичностью по отношению к ацил-КоА с определенной длиной углеродной цепи.
Стадия гидратации. Ненасыщенный ацил-КоА (еноил-КоА) при участии фермента еноил-КоА-гидратазы присоединяет молекулу воды. В результате образуется β-гидроксиацил-КоА:

Вторая стадия дегидрирования. Образовавшийся β-гидроксиацил-КоА затем дегидрируется. Эту реакцию катализируют НАД-зависимые дегидрогеназы. Реакция протекает по следующему уравнению:

Тиолазная реакция. В этой реакции β-кетоацил-КоА взаимодействует с коэнзимом А. В результате происходит расщепление β-кетоацил-КоА и образуется укороченный на два углеродных атома ацил-КоА и двууглеродный фрагмент в виде ацетил-КоА. Данная реакция катализируется ацетил-КоА-ацилтрансфе-разой (или тиолазой):

Образовавшийся ацетил-КоА подвергается окислению в цикле трикарбоновых кислот (цикле Кребса), а ацил-КоА, укоротившийся на два углеродных атома, снова многократно проходит весь путь β-окисления вплоть до образования бутирил-КоА (4-углеродное соединение), который в свою очередь окисляется до двух молекул ацетил-КоА (см. схему).
Читайте также: Сумка из фольгированной ткани

Например, в случае пальмитиновой кислоты (С16) повторяются 7 циклов окисления. Запомним, что при окислении жирной кислоты, содержащей n углеродных атомов, происходит n/2 — 1 циклов β-окисления (т. е. на один цикл меньше, чем n/2 , так как при окислении бутирил-КоА сразу происходит образование двух молекул ацетил-КоА) и всего получится n/2 молекул ацетил-КоА.
Следовательно, суммарное уравнение р-окисления пальмитиновой кислоты можно написать так:
Пальмитоил-КоА + 7 ФАД + 7 НАД + 7Н2O + 7HS-KoA —> 8 Ацетил — КоА + 7 ФАДН2 + 7 НАДН2.
Баланс энергии. При каждом цикле β-окисления образуются 1 молекула ФАДН2 и 1 молекула НАДН2. Последние в процессе окисления в дыхательной цепи и сопряженного с ним фосфорилирования дают: ФАДН2 — две молекулы АТФ и НАДН2 — три молекулы АТФ , т. е. в сумме за один цикл образуется 5 молекул АТФ. В случае окисления пальмитиновой кислоты проходит 7 циклов β-окисления (16/2 — 1 = 7), что ведет к образованию 5X7 = 35 молекул АТФ. В процессе β-окисления пальмитиновой кислоты образуется — молекул ацетил-КоА, каждая из которых, сгорая в цикле трикарбоновых кислот, дает 12 молекул АТФ , а 8 молекул дадут 12X8 = 96 молекул АТФ.
Таким образом, всего при полном окислении пальмитиновой кислоты образуется 35+96=131 молекула АТФ. Однако с учетом одной молекулы АТФ, потраченной в самом начале на образование активной формы пальмитиновой кислоты (пальмитоил-КоА), общий энергетический выход при полном окислении одной молекулы пальмитиновой кислоты в условиях животного организма составит 131-1 = 130 молекул АТФ (заметим, что при полном окислении одной молекулы глюкозы образуется лишь 36 молекул АТФ).
Подсчитано, что если изменение свободной энергии системы (ΔG) при полном сгорании одной молекулы пальмитиновой кислоты составляет 9797 кДж, а богатая энергией концевая фосфатная связь АТФ характеризуется величиной около 34,5 кДж, то выходит, что примерно 45% всей потенциальной энергии пальмитиновой кислоты при ее окислении в организме может быть использовано для ресинтеза АТФ, а оставшаяся часть, по-видимому, теряется в виде тепла.
Окисление ненасыщенных жирных кислот
Окисление ненасыщенных жирных кислот в принципе происходит так же, как и окисление насыщенных жирных кислот. Однако здесь имеются некоторые особенности. Двойные связи природных ненасыщенных жирных кислот (олеиновой, линолевой и т. д.) имеют цис-конфигурацию, а в КоА-эфирах ненасыщенных кислот, являющихся промежуточными продуктами при β-окислении насыщенных жирных кислот, двойные связи имеют транс-конфигурацию. Кроме того, последовательное удаление двууглеродных фрагментов при окислении ненасыщенных жирных кислот до первой двойной связи дает Δ 3,4 -ацил-КоА, а не Δ 2,3 -ацил-КоА, который является промежуточным продуктом при β-окислении насыщенных жирных кислот:

Оказалось, что в тканях существует фермент, который осуществляет перемещение двойной связи из положения 3-4 в положение 2-3, а также изменяет конфигурацию двойной связи из цис- в транс-. Этот фермент получил название Δ 3,4 -цис-Δ 2,3 -трансеноил-КоА-изомеразы. Ниже приводится путь окисления олеиновой кислоты, иллюстрирующей назначение дополнительного фермента. При окислении жирных кислот, имеющих две и более ненасыщенные связи, требуется еще один дополнительный фермент β-гидроксиацил-КоА-эпимераза.
Как происходит окисление жирных кислот в тканях
а) Расщепление жирных кислот до ацетилкоэнзима А путем бета-окисления. Молекула жирной кислоты расщепляется в митохондрии путем постепенного отщепления двууглеродных фрагментов в виде ацетилкоэнзима А (ацетил-КоА). Этот процесс, ход которого представлен на рисунке ниже, называют процессом бета-окисления.
Бета-окисление жирных кислот в результате дает ацетилкоэнзим А
Обратите внимание, что первый этап бета-окисления представляет собой взаимодействие молекулы жирной кислоты с коэнзимом А (КоА) с образованием ацил-КоА жирной кислоты. В уравнениях 2, 3 и 4 бета-углерод (второй углерод справа) ацил-КоА жирной кислоты взаимодействует с молекулой кислорода, вследствие этого бета-углерод окисляется.
Читайте также: Какие методы определения волокнистого состава тканей вы знаете
В правой части уравнения 5 двууглеродная часть молекулы отщепляется, образуя ацетил-КоА, выделяющийся во внеклеточную жидкость. В то же время другая молекула КоА взаимодействует с концом оставшейся части молекулы жирной кислоты, вновь формируя ацил-КоА жирной кислоты. Сама молекула жирной кислоты в это время становится короче на 2 атома углерода, т.к. первый ацетил-КоА уже отделился от ее терминали.
Затем эта укоротившаяся молекула ацил-КоА жирной кислоты выделяет еще 1 молекулу ацетил-КоА, что приводит к укорочению исходной молекулы жирной кислоты еще на 2 атома углерода. Кроме высвобождения молекул ацетил-КоА из молекул жирных кислот в ходе этого процесса выделяются 4 атома углерода.
б) Окисление ацетил-КоА. Образующиеся в митохондриях в ходе процесса бета-окисления жирных кислот молекулы ацетил-КоА немедленно поступают в цикл лимонной кислоты и, взаимодействуя прежде всего с щавелево-уксусной кислотой, образуют лимонную кислоту, которая затем последовательно окисляется посредством хемоосмотическои. системы окисления митохондрий. Чистый выход реакции цикла лимонной кислоты на 1 молекулу аце-тил-КоА составляет:

Таким образом, после начального расщепления жирной кислоты с образованием ацетил-КоА окончательное их расщепление осуществляется так же, как расщепление ацетил-КоА, образовавшегося из пировиноградной кислоты в процессе метаболизма глюкозы. Образующиеся при этом атомы водорода окисляются той же системой окисления митохондрий, которая используется в процессе окисления углеводов, с образованием большого количества аденозинтрифосфата.
в) При окислении жирных кислот образуется огромное количество АТФ. На рисунке выше показано, что 4 атома водорода, высвобождающиеся при отделении ацетил-КоА от цепочки жирной кислоты, выделяются в виде ФАДН2, НАД-Н и Н+, поэтому при расщеплении 1 молекулы стеариновой кислоты образуется, кроме 9 молекул ацетил-КоА, еще 32 атома водорода. В процессе расщепления каждой из 9 молекул ацетил-КоА в цикле лимонной кислоты выделяются еще 8 атомов водорода, что в итоге дает 72 атома водорода.
Суммарно при расщеплении 1 молекулы стеариновой кислоты выделяются 104 атома водорода. Из этого общего количества 34 атома выделяются, будучи связанными с флавопротеинами, а остальные 70 высвобождаются в форме, связанной с никотинамидадениндинуклеотидом, т.е. в виде НАД-Н+ и Н+.
Окисление водорода, связанного с этими двумя типами веществ, осуществляется в митохондриях, но они вступают в процесс окисления в разных точках, поэтому окисление каждого из 34 атомов водорода, связанных с флавопротеинами, приводит к выделению 1 молекулы АТФ. Еще 1,5 молекулы АТФ синтезируется из каждых 70 НАД+ и Н+. Это дает к 34 еще 105 молекул АТФ (т.е. всего 139) при окислении водорода, отщепляющегося при окислении каждой молекулы стеариновой кислоты.
Дополнительно 9 молекул АТФ образуются в цикле лимонной кислоты (помимо АТФ, получаемой при окислении водорода), по 1 на каждую из 9 молекул метаболизирующегося ацетил-КоА. Итак, при полном окислении 1 молекулы стеариновой кислоты образуются в сумме 148 молекул АТФ. С учетом того, что при взаимодействии стеариновой кислоты с КоА на начальной стадии метаболизма этой жирной кислоты расходуются 2 молекулы АТФ, чистый выход АТФ составляет 146 молекул.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
— Вернуться в оглавление раздела «Физиология человека.»
Окисление жирных кислот
Окисление жирных кислот протекает в печени, почках, скелетных и сердечных мышцах, в жировой ткани.
Ф.Кнооп высказал предположение, что окисление молекулы жирной кислоты в тканях организма происходит в b-окислении. В результате от молекулы жирной кислоты отщепляются двууглеродные фрагменты со стороны карбоксильной группы. Процесс b-окисления жирных кислот складывается из следующих этапов:
Читайте также: Ткань сделанная из натурального сырья это ответ
Активация жирных кислот. Подобно первой стадии гликолиза сахаров перед b-окислением жирные кислоты подвергаются активации. Эта реакция протекает на наружной поверхности мембраны митохондрий при участии АТФ, коэнзима А (НS-КоА) и ионов Mg 2+ . Реакция катализируется ацил-КоА-синтетазой:
В результате реакции образуется ацил-КоА, являющийся активной формой жирной кислоты.
Транспорт жирных кислот внутрь митохондрий.Коэнзимная форма жирной кислоты, в равной мере как и свободные жирные кислоты, не обладает способностью проникать внутрь митохондрий, где, собственно, и протекает их окисление, переносчиком активированных жирных кислот через внутреннюю митохондриальную мембрану служит карнитин (g-триметиламино-b-оксибути-рат):
После прохождения ацилкарнитина через мембрану митохондрий происходит обратная реакция – расщепления ацилкарнитина при участии НS-КоА и митохондриальной карнитин-ацилтрансферазы:
Ацил-КоА в митохондрии подвергается процессу b-окисления.
Этот путь окисления связан с присоединением атома кислорода к углеродному атому жирной кислоты, находящемуся в b-положении:
При b-окислении происходит последовательное отщепление от карбоксильного конца углеродной цепи жирной кислоты двууглеродных фрагментов в форме ацетила-КоА и соответствующее укорачивание цепи жирной кислоты:
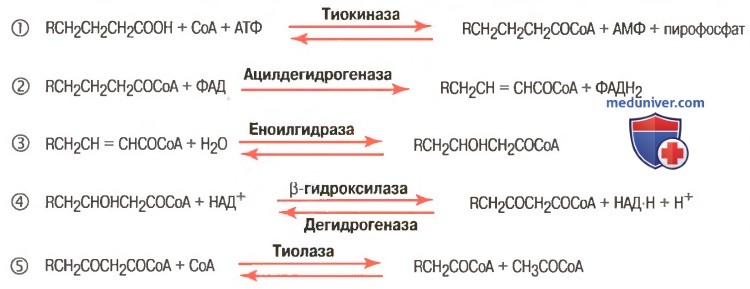
В матриксе митохондрии ацил-КоА распадается в результате повторяющейся последовательности четырех реакций (рис.8).
1) окисление с участием ацил-КоА-дегидрогеназы (ФАД-зависимой дегидрогеназы);
2) гидратация, катализируемой еноил-КоА-гидратазой;
3) второго окисления под действием 3-гидроксиацетил-КоА-дегидрогеназы (НАД-зависимой дегидрогеназы);
4) тиолиза с участием ацетил-КоА-ацилтрансферазы.
Совокупность этих четырех последовательностей реакций составляет один оборот b-окисления жирной кислоты (см. рис. 8).
Образовавшийся ацетил-КоА подвергается окислению в цикле Кребса, а ацетил-КоА, укоротившийся на два углеродных атома, снова многократно проходит весь путь b-окисления вплоть до образования бутирил-КоА (4-углеродное соединение), на последнем этапе b-окисления распадается на две молекулы ацетил-КоА.
При окислении жирной кислоты, содержащей n углеродных атомов, происходит n/2-1 цикл b-окисления (т.е. на один цикл меньше, чем n/2, так как при окислении бутирил-КоА сразу происходит образование двух молекул ацетил-КоА) и всего получится n/2 молекул ацетил-КоА.
Например при окислении пальмитиновой кислоты (С16) повторяется 16/2-1=7 циклов b-окисления и образуется 16/2=8 молекул ацетил-КоА.
Рисунок 8 – Схема b-окисления жирной кислоты
Баланс энергии.При каждом цикле b-окисления образуется одна молекула ФАДН2 (см. рис. 8; реакция 1) и одна молекула НАДН+Н + (реакция 3). Последняя в процессе окисления дыхательной цепи и сопряженного с ним фосфорилирования дают: ФАДН2 – 2 молекулы АТФ и НАДН+Н + – 3 молекулы АТФ, т.е. в сумме за один цикл образуется 5 молекул АТФ. При окислении пальмитиновой кислоты образуется 5*7=35 молекул АТФ. В процессе b-окисления пальмитиновой кислоты образуется 8 молекул ацетил-КоА, каждая из которых, «сгорая» в цикле Кребса, дает 12 молекул АТФ, а 8 молекул дадут 12*8=96 молекул АТФ.
Таким образом, всего при полном b-окислении пальмитиновой кислоты образуется 35+96=131 молекула АТФ. С учетом одной молекулы АТФ, потраченной в самом начале на стадии активации жирной кислоты, общий энергетический выход при полном окислении одной молекулы пальмитиновой кислоты составит 131-1=130 молекул АТФ.
Однако, образовавшийся в результате b-окисления жирных кислот ацетил-КоА, может не только окисляться до СО2, Н2О, АТФ, вступая в цикл Кребса, но использоваться на синтез холестерина , а также углеводов в глиоксилатном цикле.
Глиоксилатный путь специфичен только для растений и бактерий, у животных организмов он отсутствует. Данный процесс синтеза углеводов из жиров подробно описан в методическом указании «Взаимосвязь процессов обмена углеводов, жиров и белков» (см. п. 2.1.1, с. 26).
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом