
Какая из перечисленных тканей обладает возбудимостью

Сходство нервной и мышечной тканей состоит в том, что они обладают свойством
Свойства нервной ткани — возбудимость и проводимость; мышечной ткани — возбудимость и сократимость мышц под действием возбуждения.
Возбудимость — это свойство ткани генерировать потенциал действия на раздражение. К возбудимым клеткам относятся только те, которые генерируют потенциал действия. Это мышечные и нервные клетки. Потенциал действия проводится по мембранам нервных и мышечных клеток, с его помощью передается информация и обеспечивается управление деятельностью клеток организма.
Раздражимостью называется способность живых организмов и образующих их систем (органов, тканей, клеток) реагировать на внешнее воздействие изменением своих физико-химических и физиологических свойств. Раздражимость является универсальным проявлением жизнедеятельности всех без исключения биологических систем.
Возбудимостью называется способность организма, органа, ткани или клетки отвечать на раздражение активной специфической реакцией — возбуждением (генерацией нервного импульса, сокращением, секрецией и др. ).
Раздражимость и возбудимость характеризуют в сущности одно и то же свойство биологической системы — способность отвечать на внешние воздействия. Однако термин возбудимость используется для определения специфических реакций, имеющих более позднее филогенетическое происхождение. Возбудимость является, следовательно, высшим проявлением более общего свойства раздражимости тканей.
Проводимость — это способность ткани и клетки проводить возбуждение. Процессы возбуждения и торможения нервных клеток (электрические явления) обеспечивают выполнение их функций. Особенностью нервной ткани является то, что возбуждение не остается на месте его возникновения, а передается по нервным волокнам.
Почему в этом вопросе нельзя ответить проводимость? Разве нервная и мышечная ткань не обладают проводимостью? Объясните, пожалуйста.
Согласна, т.к. возбуждение и проведение возбуждения «пограничные понятия».
Под проводимостью следует понимать способность ткани проводить возбуждение в виде потенциала действия. В основе проведения возбуждения по мышечной ткани лежат циркулярные или круговые токи, которые формируются в результате перезарядки мембраны. Однако, проведение возбуждения по мышечному волокну отличается от проведения возбуждения по нервной ткани тем, что потенциал действия в этом случае должен достигнуть мышечных сократительных элементов — актиновых и миозиновых нитей, которые находятся в глубине клетки. Проведение возбуждения к сократительным элементам мышечного волокна обеспечивается мембранно-миофибриллярной связью.
НО, всё же в школьных учебниках нам даются свойства тканей именно так:
Свойства нервной ткани — возбудимость и проводимость.
А физиологическими свойствами мышечной ткани являются возбудимость и сократимость. Сократимость можно определить как способность мышцы укорачиваться, уменьшая свою длину или напрягаться без ее изменения.
Мышечные ткани
Мышечные ткани — это ткани, для которых способность к сокращению является главным свойством. Мышечные ткани составляют активную часть опорно-двигательного аппарата (пассивной частью являются кости, соединения костей).
Общими свойствами всех мышечных тканей является сократимость и возбудимость. К данной группе тканей относятся гладкая, поперечнополосатая скелетная и поперечнополосатая сердечная мышечные ткани. Клетки мышечной ткани имеют хорошо развитый цитоскелет, содержат много митохондрий.

Гладкая (висцеральная) мускулатура
Эта мышечная ткань встречается в стенках внутренних органах (бронхи, кишечник, желудок, мочевой пузырь), в стенках сосудов, протоках желез. Эволюционно является наиболее древним видом мускулатуры.
Состоит из веретенообразных миоцитов — коротких одноядерных клеток. Между клетками имеются межклеточные контакты — нексусы (лат. nexus — связь). Благодаря нексусам возбуждение, возникшее в одной клетке, волнообразно распространяется на все остальные клетки.

Гладкая мышечная ткань отличается своей способностью к длительному тоническому напряжению, что очень важно для работы внутренних органов (к примеру мочевого пузыря), сокращается медленно, практически не утомляется. Скелетная мышечная ткань, которую мы изучим чуть позже, такой способностью не обладает — сокращается и утомляется быстро.
Осуществляется сокращение с помощью клеточных органоидов — миофиламентов, которые расположены в клетке хаотично и не имеют такой упорядоченной структуры, как миофибриллы в скелетной мускулатуре (все познается в сравнении, уже скоро мы их тоже изучим).
Особо заметим, что в гладкой мышечной ткани миофиламенты собираются в миофибриллы только во время сокращения. У таких временных миофибрилл не может быть регулярной организации, а значит ни у таких миофибрилл, ни у гладких миоцитов не может быть поперечной исчерченности.
Гладкая мышечная ткань сокращается непроизвольно (неподвластна воле человека). Работа гладких мышц обеспечивается вегетативной (автономной) нервной системой. К примеру невозможно по желанию сузить или расширить бронхи, кровеносные сосуды, зрачок.

Гладкая мышечная ткань называется неисчерченной, так как не обладает поперечной исчерченностью, характерной для поперечнополосатых скелетной и сердечной мышечных тканей.
Скелетная (поперечнополосатая) мышечная ткань
Скелетная мышечная ткань образует диафрагму (дыхательную мышцу), мускулатуру туловища, конечностей, головы, голосовых связок.
В отличие от гладкой мускулатуры, скелетная образована не отдельными одноядерными клетками, а длинными многоядерными волокнами, имеющими до 100 и более ядер — миосимпластами. Миосимпласт (греч. sim — вместе + plast — образованный) представляет совокупность слившихся клеток, имеет длину от нескольких миллиметров до нескольких сантиметров (соответствует длине мышцы).
Внутри миосимпласта находится саркоплазма, снаружи миосимпласт покрыт сарколеммой. Сократительные элементы — миофибриллы (лат. fibra — волоконце) — длинные тяжеобразные органеллы в миосимпласте (около 1400).

Характерная черта данной ткани — поперечная исчерченность, выражающаяся в равномерном чередовании светлых и темных полос на мышечном волокне. Это происходит потому, что границы саркомеров в соседних миофибриллах совпадают, вследствие чего все волокно приобретает поперечную исчерченность. Теперь самое время изучить микроскопическую основу мышцы — саркомер.
Саркомер (от греч. sarco — мясо (мышца) + mere — маленький)
Саркомер — элементарная сократительная единица поперечнополосатых мышц, структурная единица миофибриллы. В состав саркомера (и миофибриллы в целом) входят миофиламенты (лат. filamentum — нить) двух типов, которые обеспечивают сократимость мышечной ткани.
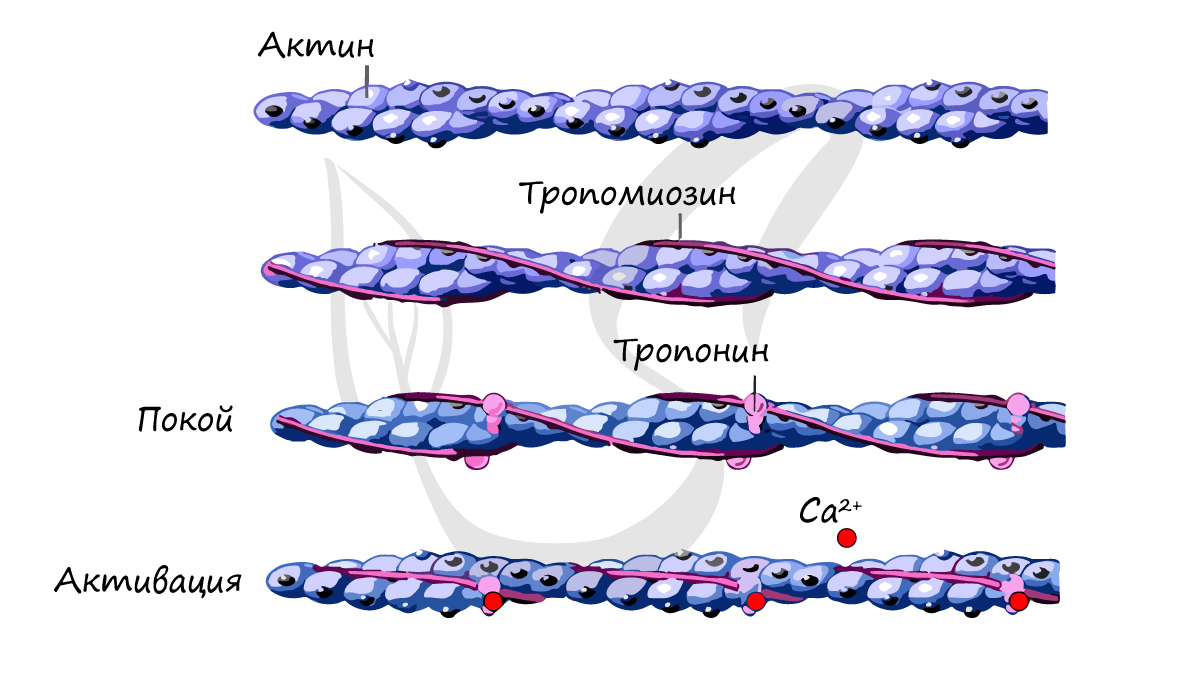
Саркомер состоит из актиновых (тонких) и миозиновых (толстых) филаментов, которые образованы главным образом белками актином и миозином. Сокращение происходит за счет взаимного перемещения миофиламентов: они тянутся навстречу друг другу, саркомер укорачивается (и мышца в целом).

Источником энергии для сокращения служат молекулы АТФ. К тому же невозможно представить сокращение мышц без участия ионов кальция: именно они связываются с тропонином, что приводит к изменению конформации тропомиозина (тропонин и тропомиозин — регуляторные белки между нитями актина), за счет чего становится возможно соединение актина и миозина. При сокращении мышц выделяется тепло (сократительный термогенез).
Замечу, что трупное окоченение (лат. rigor mortis) — посмертное затвердевание мышц — связано именно с ионами кальция, которые устремляются в область низкой концентрации (в саркоплазму миосимпласта), способствуя связыванию актина и миозина.
После смерти в мышце перестает синтезироваться АТФ, ее уровень быстро снижается. Как следствие этого перестает функционировать Ca-АТФаза — насос, выкачивающий ионы Ca из саркоплазмы в саркоплазматический ретикулум (мембранная органелла мышечных клеток (сходная с ЭПС), в которой запасаются ионы Ca).
В саркоплазме повышается концентрация ионов Ca — замыкаются мостики между актином и миозином, однако разомкнуться они уже не могут, в связи с чем наблюдается стойкая мышечная контрактура (лат. contractura — стягивание, сужение): конечности очень сложно разогнуть или согнуть.

Вернемся к скелетным мышцам. Имеется еще ряд важных моментов, о которых нужно знать.
Читайте также: Ткань для валяния это
В процесс возбуждения вовлекается изолированно один миосимпласт, соседние миосимпласты (волокна) не возбуждают друг друга, в отличие от гладких миоцитов, где возбуждение предается между соседними клетками через нексусы. Скелетные мышцы сокращаются быстро и быстро утомляются (у гладких мышц фазы сокращения и расслабления растянуты во времени, мало утомляются) .
Скелетные мышцы сокращаются произвольно: они подконтрольны нашему сознанию. К примеру, по желанию мы можем изменить скорость движения руки, темп бега, силу прыжка. Мышцы покрыты фасцией, крепятся к костям сухожилиями, и, сокращаясь, приводят в движение суставы.

Сердечная поперечнополосатая мышечная ткань
Сердечная мышечная ткань образует мышечную оболочку сердца — миокард (от др.-греч. μῦς «мышца» + καρδία — «сердце»). Миокард — средний слой сердца, составляющий основную часть его массы. При работе сердечная мышечная ткань не утомляется.

Сердечная мышечная ткань состоит из кардиомиоцитов — одиночных клеток, имеющих поперечную исчерченность. Соединяясь друг с другом, кардиомиоциты образуют функциональные волокна.
Этот тип мышечной ткани удивительным образом сочетает свойства двух предыдущих, изученных нами, тканей (возбудимость, сократимость) и имеет одно новое уникальное свойство — автоматизм.
Автоматизм — способность сердечной мышечной ткани возбуждаться и сокращаться самопроизвольно, без влияний извне. Это легко можно подтвердить, наблюдая сокращения изолированного сердца лягушки в физиологическом растворе: сокращения сердца в нем будут продолжаться несколько десятков минут после отделения сердца от организма.

Места контактов соседних кардиомиоцитов — вставочные диски (в их составе находятся нексусы), благодаря которым возбуждение одной клетки передается на соседние, таким образом волнообразно охватываются возбуждением и сокращаются новые участки миокарда.
Большое число контактов между кардиомиоцитами обеспечивает высокую эффективность и надежность проведения возбуждения по миокарду. Сокращается эта ткань непроизвольно, не утомляется.
На рисунке или микропрепарате узнать данную ткань можно по центральному положению ядер в клетках, поперечной исчерченности, наличию вставочных дисков и анастомозов (греч. anastomosis — отверстие) — мест соединений боковых поверхностей функциональных волокон (кардиомиоцитов).

В норме возбуждение проводится по проводящей системе сердца от предсердий к желудочкам (однонаправленно). Участок сердечной мышцы, в котором генерируются импульсы, определяющие частоту сердечных сокращений — водитель сердечного ритма.
Автоматизм возможен благодаря наличию в миокарде особых пейсмекерных (англ. pacemaker — задающий ритм) клеток, которые также называют водителями ритма. Они спонтанно генерируют нервные импульсы, которые охватывают весь миокард, в результате чего осуществляется сокращение. Именно благодаря водителям ритма сердце лягушки продолжает биться, будучи полностью отделенным от тела.
Ответ мышц на физическую нагрузку
Физические нагрузки приводят к гипертрофии мышц (от др.-греч. ὑπερ- чрез, слишком + τροφή — еда, пища) — в них увеличивается количество мышечных волокон, объем мышечной массы нарастает.

В условиях гиподинамии (от греч. ὑπό — под и δύνᾰμις — сила), то есть пониженной активности, мышцы уменьшаются вплоть до полной атрофии (греч. а – «не» + trophe – питание). В худшем случае волокна мышечной ткани перерождаются в соединительную ткань, после чего пациент становится обездвиженным.

Необходимо отметить, что сердечная мышечная ткань также дает ответную реакцию на чрезмерную нагрузку: сердце увеличивается в размере, нарастает масса миокарда. Причиной могут быть генетические заболевания, повышенное артериальное давление. Гипертрофия сердца — состояние, требующее вмешательства врача и наблюдения за пациентом.
В большинстве случае гипертрофия сердца обратима, а у спортсменов наблюдается так называемая физиологическая гипертрофия (вариант нормы).

Происхождение мышц
Мышцы развиваются из среднего зародышевого листка — мезодермы.

© Беллевич Юрий Сергеевич 2018-2021
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Какая из перечисленных тканей обладает возбудимостью
tsyganskiymedvet.ru
Главная страница
Моя работа
Категории
Владельцы сайта
Возбудимые ткани
1. Возбудимые ткани, их характеристика
2. Биоэлектрические потенциалы в возбудимых тканях
3. Основные свойства возбудимых тканей
5. Строение скелетных мышц
6. Свойства скелетных мышц
7. Сила, работа, утомление мышц
1. Возбудимые ткани, их характеристика
Раздражимость (или реактивность) – это способность органов и тканей, клеток активно отвечать на воздействие какой-либо формой деятельности – обменом, ускорением деления, выбросом секрета, движением, электрическим импульсом. Ответ на раздражение является важным свойством организма.
Возбудимость – одна из форм проявления раздражимости связанная с деятельностью специализированных клеток и тканей. Способные к возбуждению клетки – мышечные, нервные, железистые – называют возбудимыми. Для возникновения возбуждения необходимо раздражение возбудимой ткани, т.е. воздействие на живую ткань раздражителя.
Раздражитель – это агент внешней или внутренней среды организма, который действуя на клетки, ткани, органы и организм в целом вызывает возбуждение.
1. По энергетической природе:
физические – механические, температурные, электрические, лучевые;
химические – кислоты, щёлочи, яды;
биологические – животные, растения, бактерии вирусы и др.
внешние (различные формы раздражения окружающей организм материи);
внутренние (изменение химического состава крови, лимфы, тканевой, спинномозговой жидкости).
пороговые – раздражители минимальной силы (энергии), необходимой для раздражения клетки. Такая сила называется пороговой (порогом). Чем ниже порог возбуждения, тем выше возбудимость (у нерва порог возбуждения ниже, чем у мышцы и особенно железы);
подпороговые – раздражители, сила которых меньше пороговой, называются;
сверхпороговые – раздражители, более сильные, чем пороговые.
4. По биологическому значению для ткани, организма:
адекватные – естественные раздражители, действующие на данную ткань в природных условиях;
неадекватные – раздражители, действию которых ткань в естественных условиях не подвергается. К ним относятся физические, механические, электрические раздражители.
При отсутствии достаточных раздражителей в клетке наблюдается состояние физиологического покоя при условии текущего метаболизма. Реакции клетки под воздействием внешних факторов или других клеток организма, выражающиеся в снижении метаболизма, роста, возбудимости, называются торможением.
Возбуждение – реакция возбудимой клетки на действие раздражителя, проявляющаяся в совокупности физических, физико-химических и функциональных изменений. Различают общие и специфические признаки возбуждения:
общие признаки – изменение мембранного потенциала (генерация потенциала действия);
специфические признаки – переход от состояния физиологического покоя к свойственной деятельности: для нервной ткани – это генерация распространяющегося импульса, синтез и разрушение медиаторов; для мышечной – сокращение; для железистой – секреция.
Для возникновения возбуждения наряду с силой и временем действия раздражителя имеет значение быстрота нарастания силы раздражителя. Так, при медленном сдавливании пинцетом нерва возбуждение не наступает, а быстром ударе по нерву моментально возникает ответная реакция, т.к. возникло возбуждение. Скорость нарастания силы раздражителя называется градиентом раздражения. Высокий градиент раздражения характеризует мгновенное нарастающее по силе раздражение до своей пороговой величины. Чем ниже градиент возбуждения, тем ниже возбудимость раздражаемых тканей. Приспособление ткани к медленно нарастающей силе раздражения называют аккомодацией. Это объясняется тем, что за длительное время действия раздражителя в клетке происходят изменения, повышающие порог возбудимости. У разных видов тканей скорость аккомодации неодинакова: высокая у двигательных нервов, мала у сердечной мышцы, гладких мышц желудка, кишечника.
Читайте также: Как сплести из ниток ткань
2. Биоэлектрические потенциалы в возбудимых тканях
Первые сведения о способности живых тканей генерировать электричество были получены во второй половине Х V III в. на примере рыб, имеющих электрические органы. Существование же истинного «животного электричества» как проявления процессов жизнедеятельности тканей было установлено итальянцем Л. Гальвани, опубликовавшим в 1971 г. «Трактат о силах электричества при мышечном движении». В опытах на нервно-мышечном препарате лягушки ученый обнаружил, что при замыкании между нервом и мышцей цепи из двух металлических проводников происходит сокращение мышц, что толковалось как проявление разряда электричества «предсуществовавшего» в живой ткани. Однако физик А. Вольта объяснил это явление как результат возникновения постоянного тока в цепи из двух разнородных металлов (железо, медь), где препарат служит проводником-электролитом. В 1794 г. Л. Гальвани предложил новый вариант опыта без металлических проводников: надрезал мышцу и набрасывал нерв стеклянными крючками таким образом, чтобы часть его касалась повреждённого участка, и другая часть – неповреждённого. В момент набрасывания нерва мышца сокращалась. Как доказал позже Э. Дюбуа-Реймон, мышца сокращалась из-за электрического тока, источником которого была разность потенциалов между повреждённым и неповреждённым участком мышцы (повреждённый участок заряжен отрицательно, целый – положительно. К концу ХIХ и в начале ХХ стало известно, что электрические потенциалы образуются во всех живых тканях, а возникновение и распространение возбуждения связано с изменением электрического заряда на поверхности клеточной мембраны и внутри клетки.
В настоящее время для исследования электрических явлений клетки применяют микроэлектроды (стеклянные пипетки с тонким кончиком – 0,5 мкм), заполненные электролитом. Роль проводника электрического тока играет электролит, а изолятора – стекло. Вводя кончик электрода, присоедин ё нного к регистрирующей установке, в клетку, регистрируют е ё потенциал. Благодаря данной методике в конце 50-х годов ХХ столетия А. Ходжкин, А. Хаксли, Б. Кац ( A . Hodgkin , A . Huxley , В. Katz , Великобритания) сформулировали современную мембранную теорию возникновения биоэлектрических потенциалов, получившую название теории натрий-калиевого насоса. В создание теории свой вклад также внесли В.Ю. Чаговец (Россия, 1896) и Ю. Бернштейн (Россия, 1902).
Мембрана клетки состоит из двойного слоя фосфолипидов, интегральных, трансмембранных и периферических белков, а также поверхностных гликолипидов и гликопротеинов.
В мембране существуют каналы, проницаемые для воды, для малых молекул водорастворимых веществ и для малых ионов. Особое значение имеют каналы, проницаемые для ионов Nа + , К + , Cl – , Са 2+ . В мембране обнаружены специфические (селективные) натриевые, калиевые, хлорные и кальциевые каналы, то есть каналы избирательно пропускающие названные ионы. Эти каналы могут быть открытыми или закрытыми, то есть обладают воротными механизмами. В покое практически все натриевые каналы мембраны клетки закрыты, а большинство калиевых каналов – открыты. Различная концентрация ионов по обе стороны мембраны всех клеток приводит к появлению трансмембранной разности электрического потенциала – мембранного потенциала.
Мембранный потенциал покоя
Мембранный потенциал (МП) покоя – разность электрических потенциалов между внутренней и наружной поверхностью мембраны в состоянии покоя, т.е. при отсутствии электрического или химического раздражителя (сигнала).
Потенциал покоя у разных клеток имеет величину от 50 до 90 милливольт (мВ).
Согласно современной мембранной теории потенциал покоя возникает за счет пассивного и активного движения ионов через мембрану.
Пассивное движение ионов осуществляется по градиенту концентрации и не требует затрат энергии. Клеточная мембрана в состоянии покоя более проницаема для ионов калия. Ионов калия в цитоплазме примерно в 30 раз больше, чем во внеклеточной жидкости. Они находятся в свободном состоянии и по градиенту концентрации диффундируют через клеточную мембрану во внеклеточную жидкость, в ней они не рассеиваются, а удерживаются на внешней поверхности мембраны внутриклеточными анионами.
Внутри клетки содержатся в основном анионы органических кислот: аспарагиновой, уксусной, пировиноградной и др. Содержание неорганических анионов в клетке сравнительно небольшое. Анионы не могут проникать через мембрану и остаются в клетке, располагаясь на внутренней поверхности мембраны. Так как ионы калия имеют положительный заряд, а анионы – отрицательный, то внешняя поверхность мембраны при этом заряжается положительно, а внутренняя – отрицательно.
Ионов натрия приблизительно в 10 раз больше во внеклеточной жидкости, чем в клетке, проницаемость их через мембрану незначительна. В результате пассивного движения ионов натрия и калия по концентрационному градиенту должен был бы наступить момент, когда их концентрации в клетке и внеклеточной жидкости выровнялись и разность потенциалов исчезла. Однако этого не происходит, так как в мембране существует специальный молекулярный механизм, который обеспечивает активный перенос ионов натрия и калия в сторону их повышенной концентрации и поддерживает ионную асимметрию.
Активный перенос ионов осуществляется в результате деятельности ионного Na + ,K + -насоса. Работа этого насоса заключается в перемещении ионов натрия и калия против градиента концентрации (в сторону повышенной концентрации). На каждые три иона натрия выводимые из клетки, в клетку поступают два иона калия. Na + ,K + -насос выполняет двоякую функцию: во-первых он создает и поддерживает трансмембранный градиент концентраций натрия и калия, и, во-вторых, генерирует разность потенциалов, суммирующуюся с потенциалом покоя, создаваемым пассивным движением ионов через мембрану.
Для работы насоса необходима энергия макроэргических связей АТФ. Расщепление АТФ происходит под влиянием фермента аденозинтрифосфотазы. Активность этого фермента угнетают протоплазматические яды, например, при действии цианидов прекращается работа Na + ,K + -насоса, в результате концентрация ионов по обе стороны мембраны выравнивается и клетка гибнет.
Все клетки возбудимых тканей (нервные, скелетных мышц, гладкомышечные клетки, кардиомиоциты) при действии различных раздражителей пороговой силы способны переходить в состояние возбуждения. Обязательный признак возбуждения – изменение электрического состояния клеточной мембраны. Под влиянием раздражения заряд мембраны начинает быстро уменьшаться от нуля и затем вновь появляется, но только с обратным знаком. Происходит перезарядка, или реверсия, заряда мембраны: внутренняя ее сторона заряжается положительно, а наружная – отрицательно. Затем вновь восстанавливаются первоначальные заряды мембраны. Пикообразное колебание потенциала, возникающее в результате кратковременной перезарядки мембраны и последующего восстановления ее исходного заряда называется потенциалом действия. Такое колебание потенциала продолжается 1-5 миллисекунд (мс). Величина потенциала действия нерва и скелетной мышцы составляет 115-120 мВ. В потенциале действия различают пик длительностью 0,5-0,8 мс и следовые потенциалы.
Пик потенциала действия связан с процессом возбуждения, он имеет восходящую и нисходящую фазы. Во время восходящей фазы вначале исчезает исходная поляризация мембраны, поэтому её называют фазой деполяризации, затем изменяется знак потенциала – реверсия. Во время нисходящей фазы заряд мембраны возвращается к уровню покоя, и её называют реполяризацией. При реполяризации отмечают следовые потенциалы: следовую деполяризацию и следовую гиперполяризацию. Следовые потенциалы связаны с восстановительными процессами после окончания возбуждения.
Современная мембранная теория объясняет механизм возникновения потенциала действия следующим образом. При действии раздражителей пороговой или сверхпороговой силы наступает деполяризация клеточной мембраны. Когда деполяризация достигает некоторой критической величины, одновременно за доли миллисекунды открывается максимальное количество натриевых каналов, проницаемость мембраны для ионов натрия возрастает в 500 раз по сравнению с покоем. В результате этого ионы натрия из внеклеточной жидкости, где их концентрация повышена, лавинообразно устремляются внутрь клетки и уносят с наружной поверхности мембраны положительные заряды, значительно увеличивая их концентрацию в клетке; внутренняя поверхность мембраны приобретает положительный заряд. Вследствие уменьшения положительных зарядов наружная поверхность мембраны за счет внеклеточных анионов заряжается отрицательно. Но увеличение натриевой проницаемости очень кратковременно и длится примерно 0,5-1,5 мс, после чего проницаемость мембраны для ионов натрия вновь понижается до исходного уровня в результате уменьшения количества открытых натриевых каналов. Вслед за этим проницаемость мембраны для ионов калия повышается в 10-15 раз. Они начинают усиленно выходить из клетки и, унося из нее положительные заряды, восстанавливают исходную разность потенциалов, то есть мембрана опять приобретает снаружи положительный заряд, а изнутри – отрицательный.
Читайте также: Как вывести с ткани деготь
Следовательно, потенциал действия возникает в результате прохождения ионных потоков через мембрану: движение ионов натрия внутрь клетки приводит к перезарядке мембраны, а выход из клетки ионов калия восстанавливает исходный потенциал покоя. Потоки приблизительно равны по величине, но сдвинуты во времени.
3. Основные свойства возбудимых тканей
Для возбудимых клеток и тканей характерны раздражимость, возбудимость, проводимость, функциональная подвижность (лабильность).
1. Раздражимость – общее свойство любой живой ткани, клетки реагировать на раздражение изменением обмена веществ и энергии. Следует учитывать, что изменения носят местный характер, т.е. реагирует только участок ткани, который подвергается раздражению.
2. Возбудимость – это свойство нервной, мышечной или железистой клетки отвечать на действие раздражителей изменениями ионной проницаемости мембраны и генерировать потенциал действия. Для возникновения состояния возбуждения необходимы определенные условия:
а) сила действующего раздражителя должна быть пороговой;
б) достаточное время действия раздражителя (полезное время). Чем сильнее раздражитель, тем меньше времени потребуется для возбуждения ткани.
в) действие раздражителя с большой силой нарастания (градиентом).
Для изучения возбуждения, как формы раздражимости, у живых организмов электрический ток имеет преимущество, так как легко дозируется по силе, деятельности, характеру, вызывает возбуждение, не повреждая, и после прекращения раздражения не оставляет необратимых изменений (ток образуется и в живых тканях, поэтому действие электрического тока близко к природным механизмам).
Для характеристики возбудимости тканей в электрофизиологии приняты следующие термины:
реобаза – пороговая сила раздражителя – электрического тока;
полезное время – время, необходимое для того, чтобы раздражитель пороговой силы вызвал возбуждение;
хронаксия – наименьшее время действия удвоенной реобазы (измеряют в миллисекундах или сигмах). Чем меньше хронаксия, т.е. сила тока, способная вызвать возбуждение, тем выше возбудимость ткани. Хронаксия – величина переменная, зависящая от структуры ткани, её физиологического состояния. У скелетных мышц величина хронаксии больше, чем у двигательных нервов. Самая большая хронаксия, измеряемая сотыми и десятыми долями секунды, – у гладких мышц кишечника, желудка, матки.
Изменение возбудимости ткани при возбуждении.
Для возбудимости тканей характерны фазовые изменения в зависимости от возникновения и развития импульсов. Во время деполяризации мембраны, при действии возбудителя, и появлении местного возбуждения происходит кратковременное повышение возбудимости. Когда местное возбуждение переходит в распространяющееся, возникает потенциал действия, а возбудимость ткани резко снижается. Возникает состояние, когда ткань после раздражения временно не реагирует на повторное раздражение любой силы – абсолютная рефрактерность (совпадает с пиком потенциала действия) – от лат. refrakta – невосприимчивость. Длительность фазы – в нервных волокнах – 1-10, в мышцах – до 30 мс. Период восстановления ткани после абсолютной рефрактерности, когда возбудимость ещё понижена называется относительной рефрактерностью (совпадает с фазой реполяризации и переходом его в следовую поляризацию). Длительность периода – в нерве – 20, в мышце – 50 мс.
Затем наступает следующий период – фаза экзальтации (от лат. exsaltatio – очень возбужденный) – период повышенной возбудимости, совпадающий с периодом следовой поляризации. Длительность – в нерве – 20, в мышцах – 50 мс. Со следовой гиперполяризацией совпадает фаза субнормальности, во время которой возбудимость ткани несколько снижена по сравнению с величиной возбудимости в состоянии физиологического покоя.
Очевидно, что фазы изменения возбудимости определены развитием и изменением потенциала действия в живой ткани. Изменение возбудимости зависит от действия натрий-калиевого насоса.
Самый частый ритм раздражений, на которые возбудимая ткань отвечает таким же частым ритмом возбуждения, отражает функциональное состояние ткани или лабильность. В 1928 году А.А. Ухтомский создал представление об усвоении ритма, согласно которому лабильность может изменяться под действием разных условий, в частности зависеть от обмена веществ в ткани. Понижение и повышение физиологической лабильности в связи с деятельностью, проявляющееся более высоким или низким ритмом возбуждения названо ученым усвоением ритма.
При изучении влияния на живую ткань частоты ритмов раздражения Н.Е. Введенский установил ритмы, наиболее выгодные в рабочем отношении, так называемый оптимум. При этом ритме раздражений новый импульс возникает в момент фазы экзальтации предшествующего импульса, в результате мышца сокращается максимально. Показатели оптимального ритма меньше максимального, Так максимальный ритм для двигательного нерва лягушки – 500, а оптимальный 75-150 импульсов, для мышечных волокон максимальный 150 импульсов/с, оптимальный – 20 – 50 в 1 с. Пессимум ритма раздражения возникает, когда частота раздражений настолько высока, что превышает меру лабильности. Это вызвано тем, что раздражение действует в фазу абсолютной рефрактерности предыдущего импульса. Фаза рефрактерности может углубиться, лабильность будет продолжать снижаться и наступит торможение. Перечисленные явления характерны не только для двигательных реакций, но и для условно-рефлекторных, для секреторных реакций, деятельности сердечно-сосудистой системы.
Н.Е. Введенский в классическом труде «Возбуждение, торможение и наркоз» изложил каким образом возбуждение переходит в торможение. Ученый воздействовал на среднюю часть нерва нервно-мышечного препарата лягушки эфиром, теплом, холодом, хлористым калием, сильным электрическим током, т.е. вызывал альтерацию ткани. По обеим сторонам участка накладывал электроды индукционной катушки. Электрическую активность изучал при помощи гальванометра. Ответная реакция мышцы на раздражения была неоднозначной, что дало основание выделить несколько последующих стадий:
1 стадия – уравнительная или трансформирующая. При действии перечисленных веществ различной природы, отличающихся по силе и частоте раздражения, мышца начинает отвечать одинаковыми сокращениями;
2 стадия – парадоксальная, сопровождалась сильными сокращениями мышцы при слабых или редких раздражениях, а при действии сильных раздражителей, наоборот, не сокращалась или почти не реагировала;
3 стадия – парабиотическая, сопровождалась состоянием, когда при воздействии на нерв раздражителя любой силы и частоты мышца не сокращалась, развивалось торможение, заканчивающееся состоянием, при котором возбудимость и проводимость, как проявления жизни, отсутствуют.
Эти последовательные функциональные изменения были названы Введенским – стадиями парабиотического процесса. При удалении раздражающих веществ с нерва парабиоз прекращается, функциональные свойства участка альтерируемой ткани восстанавливаются, проходя те же стадии в обратном порядке. Если воздействие раздражителей длительное, нерв отмирает.
5. Строение скелетных мышц
У всех высших животных мышцы являются важнейшими исполнительными, т.е. рабочими органами (эффекторами). У позвоночных различают мышцы: