
Какая неинкапсулированная лимфоидная ткань входит в состав иммунной системы
Непосредственно под мукозным эпителием слизистых оболочек в тесной связи с эпителиальными клетками располагаются лимфоциты пейеровых бляшек кишечника, лимфоидных фолликулов аппендикса, миндалин глотки, лимфоидных фолликулов подслизистого слоя верхних дыхательных путей и бронхов, мочеполового тракта. Все эти лимфоидные скопления получили собирательное название — мукозно-ассоциированная лимфоидная ткань.
В лимфоидных тканях, ассоциированных со слизистыми оболочками, проходят все стадии специфического иммунного ответа в тех случаях, когда антиген проникает в организм через слизистые оболочки. В мукозно-ассоциированных лимфоидных тканях активированные В-лимфоциты дифференцируются в плазматические клетки, продуцирующие специфические антитела, относящиеся к классу иммуноглобулинов A (IgA). Иммуноглобулины А, пройдя через эпителиальные клетки, где они присоединяют секреторный компонент, выходят на поверхность слизистых оболочек в форме секреторного иммуноглобулина A (SIgA), который обеспечивает местную антибактериальную и антивирусную защиту.
В качестве одного из периферических органов иммунной системы мукозно-ассоциированная лимфоидная ткань является местом:
1) распознавания антигена,
2) антигензависимой пролиферации и дифференцировки Т- и В-лимфоцитов,
3) активации Т- и В-лимфоцитов,
4) продукции цитокинов,
5) продукции и секреции специфических антител — иммуноглобулинов.
Какая неинкапсулированная лимфоидная ткань входит в состав иммунной системы
Концепция общей иммунной системы слизистых оболочек. Под общей иммунной системой слизистых оболочек подразумевают компоненты специфической защиты, реализуемые на внешних слизистых покровах и функционирующие в известной степени независимо от системной иммунологической реактивности. Эта система включает лимфоидную ткань кишечника (GALT-gut associated lymphoid tissue), лимфоидную ткань бронхов (BALT-bronchus associated lymphoid tissue), иммунокомпетентные клетки глотки, слюннных желез, респираторного тракта, молочной железы и гениталий.
Замечательная особенность иммунной системы слизистых оболочек — наличие больших количеств секреторного IgA, что является уникальной характеристикой слизистых покровов. Сывороточные иммуноглобулины и компоненты также присутствуют на различных слизистых покровах и во внешних секретах, но их роль в обеспечении локального защитного механизма менее существенна.
Суть концепции общности иммунной системы слизистых оболочек состоит в том, что выработка секреторных антител происходит не только в местах антигенного воздействия на слизистый покров, но и в отдаленных секреторных участках. Бронхиальный лимфоэпителий, пейеровы бляшки и другие организованные лимфоидные фолликулы изобилуют антиген-реактивными предшественниками В-клеток, коммитированных, главным образом, к синтезу секреторного IgA. Непосредственное антигенное раздражение — необходимое условие для первоначальной активации и пролиферации таких клеток.
Сенсибилизированные антигеном клетки при повторном контакте с тем же антигеном быстро дифференцируются в IgA-иммунобласты, пролиферирующие и мигрирующие сначала в регионарные лимфоузлы, а затем, через грудной лимфатический проток, в кровеносное русло. Во время нахождения в системе кровообращения эти клетки оседают на слизистых оболочках глаз, глотки, гениталий, слюнных и молочных желез, субэпителиальном участке слизистой оболочки верхнего и нижнего отделов респираторного тракта и в lamina propria кишечника.
Специфическая рециркуляция лимфоцитов, стимулированных в лимфоидной ткани слизистых, объясняется тем, что они имеют на своей поверхности молекулы, которые взаимодействуют со специфическими молекулами, размноженными только на поверхности клеток эндотелия венул лимфоидной ткани слизистых, что обеспечивает избирательную рециркуляцию лимфоцитов.
Здесь репопулировавшие клетки начинают активный синтез специфического секреторного IgA, играющего ведущую роль в функционировании общей иммунной системы слизистых оболочек.
Эта концепция имела многочисленные экспериментальные подтверждения. Было установлено, что клетки пейеровых бляшек кролика при введении их облученным аллогенным реципиентам избирательно распределялись в lamina propria и продуцировали антитела преимущественно класса IgA, тогда как клетки, полученные из периферических лимфоузлов доноров, осуществляли специфический homing в периферических лимфоузлах реципиентов и продуцировали антитела преимущественно класса IgG. Сходные исследования выявили, что homing бронхиальных лимфоцитов у реципиентов происходил в BALT.
Оральная иммунизация защищала лабораторных животных от интраназального заражения PC-вирусом. Интраназальное введение дендритных клеток лошадей, праймированных инактивированным герпесвирусом, приводило к появлению иммунитета у мышей.
Иммунизация мышей аналогичным образом инактивированной вакциной против гриппа сообщала устойчивость к интраназальному заражению. Двукратное введение в желудок мышей цельновирионной инактивированной сорбированной вакцины сопровождалось образованием специфических IgA и IgG в легких. При парентеральном введении вакцины в смывах легких IgA не обнаружены.
Энтеральная иммунизация против респираторно-синцитиальной инфекции индуцировала антитела в респираторном тракте, но полную защиту его обеспечивала интраназальная иммунизация. Однако, несмотря на иммунологическую реакцию отдаленных слизистых, наиболее выраженный иммунный ответ образуется в том месте, где слизистая оболочка непосредственно контактирует с вирусным антигеном. Важность локального антигенного стимулирования была убедительно продемонстрирована в экспериментах с вирусом полиомиелита. Уровень специфических секреторных IgA был максимальным в тех участках кишечника, где происходил непосредственный контакт вирусного антигена со слизистой оболочкой.
Живую вакцину Роковак против рота- и коронавирусных энтеритов новорожденным телятам вводили орально перед приемом молозива. После вакцинации заболеваемость и гибель телят от вирусных энтеритов сократились примерно в 4 раза.
В ряде опытов у животных, вакцинированных интраназально, наблюдали образование местных IgA-антител не только в носоглотке, но также в кишечнике и во влагалище. Однако при перректальной вакцинации в кишечнике появлялись IgA- и IgG-антитела в высоких титрах, но не обнаруживались во влагалище. Эти данные указывают, что для индукции местных IgA-антител во влагалище или кишечнике необходима вакцинация именно через эти органы.
Несмотря на то, что исследования по иммунитету слизистых покровов были сосредоточены в основном на изучении секреторных иммуноглобулинов и их роли в функционировании указанной системы, в ряде исследовательских работ указано на участие факторов клеточного иммунитета и, в первую очередь, Т-лимфоцитов в локальном иммунном ответе. Так, установлена миграция и homing Т-лимфоцитов после их антигенной сенсибилизации, а также показана роль Т-хелперов в регуляции местного IgA-ответа.
Представляют интерес данные интравагинальной иммунизации мышей живой вакциной против герпеса. Развивающийся при этом иммунитет может быть перенесен сингенным животным лимфоцитами генитальных лимфоузлов, но не лимфоцитами других лимфоидных органов иммунизированных доноров. Трансплантированные лимфоциты мигрировали преимущественно в ткани половых органов реципиентов, что свидетельствует о важной роли в местном иммунитете вирусиндуцированных Тц-клеток.
Однако имеются наблюдения, когда не были отмечены различия в защите животных от инфекции несмотря на наличие или отсутствие местных IgA-антител после вакцинации.
Лекция № 2 (иммунология) Иммунная система человека.
Для осуществления специфической функции надзора за генетическим постоянством внутренней среды, сохранения биологической и видовой индивидуальности в организме человека существует иммунная система. Эта система достаточно древняя, ее зачатки обнаружены еще у круглоротых.
Принцип действия иммунной системы основан на распознавании «свой-чужой», а также постоянной рециркуляции, воспроизводстве и взаимодействии ее клеточных элементов.
Структурно-функциональные элементы иммунной системы
Иммунная система — это специализированная, анатомически обособленная лимфоидная ткань.
Она разбросана по всему организму в виде различных лимфоидных образований и отдельных клеток. Суммарная масса этой ткани составляет 1—2 % от массы тела.
Анатомо-физиологический принцип устройства иммунной системы — органно-циркуляторный.
В анатомическом плане иммунная система подразделена на центральные и периферические органы.
К центральным органам иммунитета относятся
Инкапсулированные органы: селезёнка, лимфатические узлы.
Неинкапсулированная лимфоидная ткань.
Лимфоидная ткань слизистых оболочек (MALT — Mucosal–Аssociated Lymphoid Tissue). В том числе:
Читайте также: Подкладка для ткани с мембраной
Лимфоидная ткань, ассоциированная с ЖКТ (GALT — Gut–Associated Lymphoid Tissue) — миндалины, аппендикс, пейеровы бляшки, а также субпопуляция внутриэпителиальных лимфоцитов слизистой оболочки ЖКТ.
Лимфоидная ткань, ассоциированная с бронхами и бронхиолами (BALT — Bronchus–Associated Lymphoid Tissue), а также внутриэпителиальные лимфоциты слизистой оболочки дыхательной системы.
Лимфоидная ткань, ассоциированная с женскими половыми путями (VALT — Vulvovaginal–Associated Lymphoid Tissue), а также внутриэпителиальные лимфоциты их слизистой оболочки.
Лимфоидная ткань, ассоциированная с носоглоткой (NALT — Nose–Associated Lymphoid Tissue), а также внутриэпителиальные лимфоциты её слизистой оболочки.
Субпопуляции лимфоцитов печени, которые в качестве лимфоидного барьера «обслуживают» кровь воротной вены, несущей все всосавшиеся в кишечнике вещества.
Лимфоидная подсистема кожи (SALT – Skin-Associated Lymphoid Tissue) диссеминированные внутриэпителиальные лимфоциты и региональные лимфатические узлы и сосуды лимфодренажа.
Лимфоидная подсистема мозга, включающая различные субпопуляции лимфоцитов и других иммуноцитов.
Периферическая кровь — транспортно–коммуникационный компонент иммунной системы.
Таким образом, вполне оправдано выделение локальных иммунных подсистем слизистых оболочек, также как и мозга, печени, кожи и других тканей.
В каждой ткани популяции лимфоцитов и других иммуноцитов имеют свои особенности. Более того, миграция лимфоцитов в определённую ткань зависит от экспрессии на мембране так называемого homing–Рц (home — дом, место «прописки» лимфоцита).
С функциональной точки зрения можно выделить следующие органы иммунной системы:
воспроизводства и селекции клеток иммунной системы (костный мозг, тимус);
контроля внешней среды или экзогенной интервенции (лимфоидные системы кожи и слизистых);
контроля генетического постоянства внутренней среды (селезенка, лимфатические узлы, печень, кровь, лимфа).
Основными функциональными клетками являются 1) лимфоциты. Их число в организме достигает 10 12 . Кроме лимфоцитов, к числу функциональных клеток в составе лимфоидной ткани относят
2) мононуклеарные и гранулярные лейкоциты, тучные и дендритные клетки. Часть клеток сосредоточена в отдельных органах иммунной системы, другие — свободно перемещаются по всему организму.
Неинкапсулированные лимфатические фолликулы в рыхлой волокнистой соединительной ткани

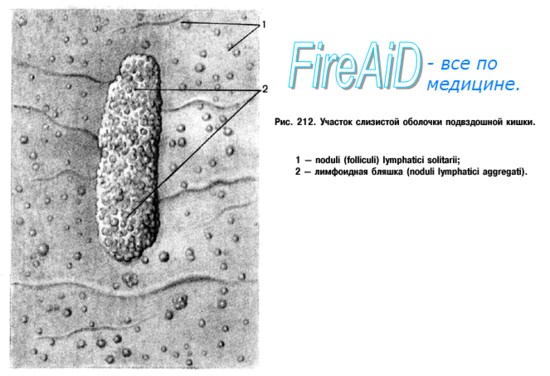
В рыхлой волокнистой соединительной ткани, расположенной под пограничным эпителием (эпителием слизистой полых органов – дыхательной, пищеварительной, мочеполовой трубок и в других областях организма) встречаются неинкапсулированные лимфоидные фолликулы, которые, как правило, находятся в тесной связи с эпителиальной тканью (расположены непосредственно под ней). Фолликулы имеют округлую форму, диаметр – до 1 мм. Большинство лимфатических фолликулов является дискретными, но в некоторых областях организма они образуют скопления, сливаясь в единую массу. Так, в области языка, глотки и носоглотки скопления лимфоидных фолликулов формируют миндалины, в рыхлой волокнистой соединительной ткани кишечника – пэйеровы бляшки. Основными структурами лимфатических фолликулов являются дифференцированные малые лимфоциты и производные В-лимфоцитов – плазматические клетки, которые способны покидать фолликул с периферической его области и заселять окружающую рыхлую волокнистую соединительную ткань.
Рис. Лимфоидный фолликул слизистой тонкого кишечника

Рис. Скопление лимфоидных фолликулов в небных миндалинах человека
Лимфатические фолликулы начинают формироваться еще в эмбриональном периоде, но количество их резко увеличивается сразу после рождения и в последующей постнатальной жизни в результате встречи лимфоцитов с антигенами, проникающими под эпителиальный пласт в соединительную ткань. Следовательно, стимулом для формирования лимфатических фолликулов является контакт малых лимфоцитов (в основном В-лимфоцитов) с антигенами, в результате чего лимфоциты начинают пролиферировать, В-лимфоциты дают начало плазматическим клеткам. Пролиферация и дифференцировка В-лимфоцитов после антигенной стимуляции и являются причиной формирования лимфатического фолликула. В связи с отмеченным, у животных, выращенных в стерильных условиях (т.н. «безмикробных животных») количество лимфатических фолликулов очень мало.
Функциональное значение лимфатических фолликулов состоит в том, что они представляют собой второй (после пограничного эпителия) эшелон защиты организма от проникающих под эпителиальный пласт антигенов.
Лимфатические узлы
Лимфатические узлы являются периферическими органами иммунной системы, в которых осуществляется
контакт лимфоцитов с антигенами, сопровождающийся активацией лимфоцитов. Процесс активации лимфоцитов, в свою очередь, включает следующие реакции:
антигензависимую пролиферацию (клонирование) определенных лимфоцитов, детерминированных на данный антиген,
последующую дифференцировку Т- и В-лимфоцитов в эффекторные клетки. В частности, В-лимфоциты превращаются в плазматические клетки, а Т-лимфоциты – в Т-хелперы или Т-киллеры (цитотоксические лимфоциты)
образование клеток-памяти (Т- или В-клеток памяти в результате активации лимфоцитов после контакта с определенным антигеном)
очистка лимфы от инородных частиц и антигенов (на 95-99%), связанная с активацией лимфоцитов и присутствием макрофагов, а также освобождение лимфы от избытка воды, белков, жиров, обогащение ее антителами и лимфоцитами
депонирование лимфы и некоторые другие процессы.
Следовательно, лимфатические узлы служат местом лимфоцитопоэза (причем не первичного (антигеннезависимого) из кроветворных стволовых клеток, которое осуществляется в центральных органах иммунной системы (красном костном мозге и тимусе), а вторичного антигензависимого образования активированных Т- и В-лимфоцитов из малодифференцированных лимфоцитов-предшественников, коммитированных на данный антиген), иммунной защиты и депонирования лимфы.
Лимфатические узлы имеют овально-округлую форму, диаметр – 0,5-1 см. Они располагаются по ходу крупных лимфатических сосудов, образуя большие скопления в подкрыльцовой впадине, паху, вдоль крупных сосудов шеи, в грудной клетке и брюшной полости, особенно по ходу крупных сосудов, в брыжейке и некоторых других областях организма. Следовательно, лимфатические узлы, в отличие от лимфатических фолликулов, локализованы не там, где образуется лимфа, а вдоль главных ветвей грудного и правого лимфатического протоков, через посредство которых лимфа возвращается в кровь. Обычно у лимфатических узлов с одной стороны имеется вдавление, называемое воротами. В области ворот в узел входят артерии и нервы, выходят вены и выносящие (эфферентные) лимфатические сосуды. Противоположная вдавлению сторона узла имеет выпуклую форму и называется наружной его стороной; она соответствует месту проникновения в лимфатический узел приносящих (аффрентных) лимфатических сосудов. Таким образом, лимфа входит в лимфатический узел со стороны наружной его поверхности, проходит через него и выходит по выносящим лимфатическим сосудам в области ворот узла.
Лимфатический узел покрыт соединительнотканной капсулой, содержащей большое количество жировых клеток. От капсулы узла со стороны его ворот отходят, направленные внутрь многочисленные соединительнотканные трабекулы, достигающие наружной поверхности узла, образующие в совокупности соединительнотканнную его строму и выполняющие опорную и трофическую функции.
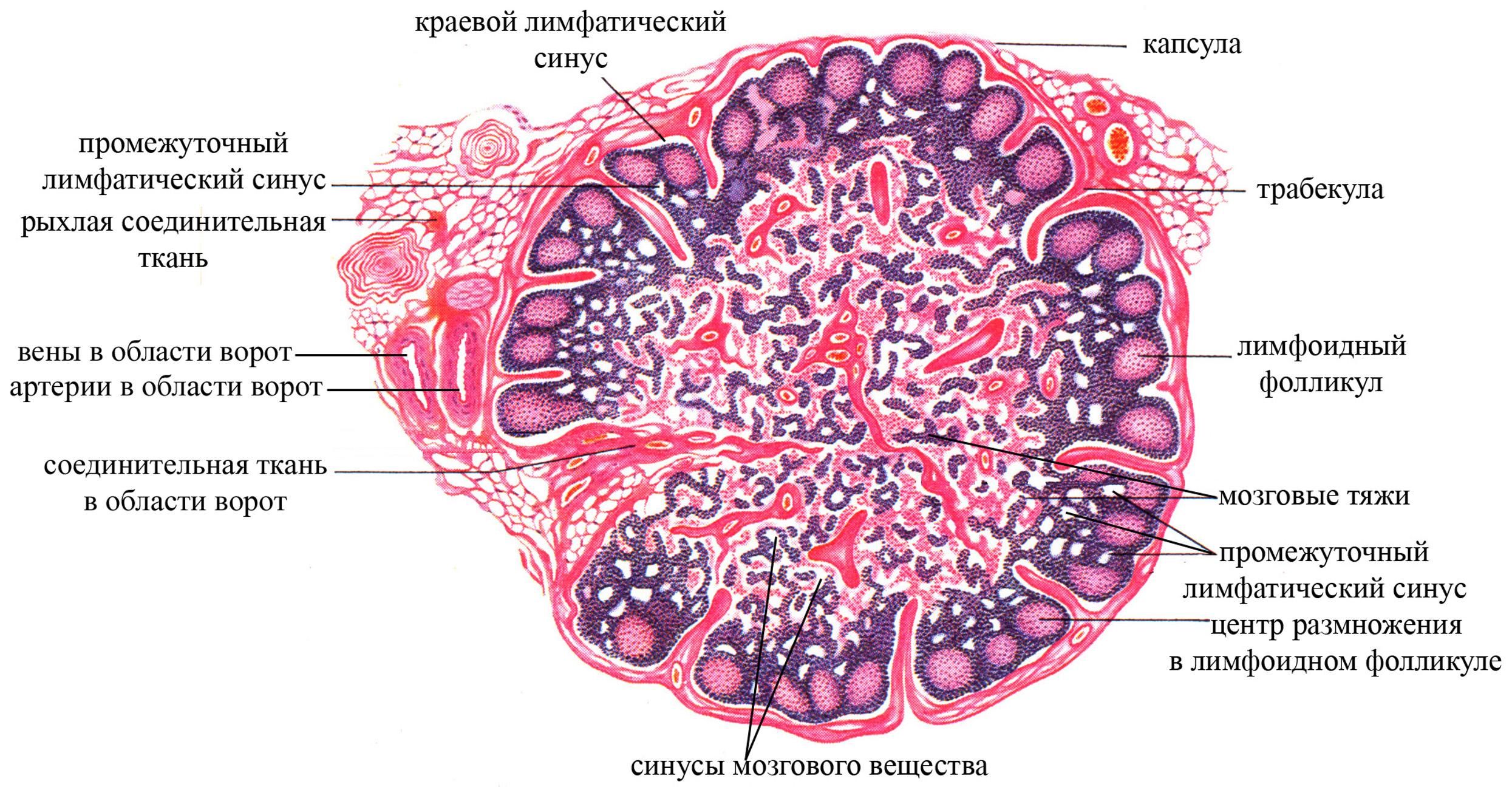
Рис. Гистоструктура лимфатического узла
В лимфатическом узле выделяют корковое и мозговое вещество, а также лимфатические синусы (полости в лимфатическом узле, определенным образом в нем локализующиеся, выстланные лимфатическим эндотелием или эндотелиоподобными ретикулярными клетками, заполнены лимфой).

Корковое вещество занимает периферическое положение в узле, имеет более плотную структуру, в периферической своей части содержит лимфатические фолликулы (называемые также первичными лимфатическими фолликулами и идентичные неинкапсулированным лимфатическим фолликулам рыхлой волокнистой соединительной ткани). Лимфатические фолликулы имеют округлую форму, диаметр до 0,5 мм, представлены лимфоидными клетками на разных этапах дифференцировки (преимущественно В-лимфоцитами, среди которых в гораздо меньшем количестве находятся Т-хелперы) и макрофагами, залегающими в ретикулярной ткани. Снаружи лимфатические фолликулы покрыты эндотелиоподобными ретикулярными клетками, лежащими на пучках ретикулярных фибрилл. В толще коркового вещества в радиальном направлении проходят соединительнотканные трабекулы (являются производными соединительнотканной капсулы узла) и лимфатические синусы, разделяя ретикулолимфоидную его структуру на отдельные дольки, имеющие форму пирамид (основание этих пирамид обращено к соединительнотканной капсуле, а вершины – к мозговому веществу и переходят в мякотные тяжи). Именно в пирамидах коркового вещества и залегают лимфатические фолликулы.
Читайте также: Ткани для детских веще
Рис. Гистоструктура коркового вещества лимфатического узла
Мозговое вещество занимает центральное положение в лимфатическом узле, представлено мякотными тяжами, которые являются продолжением коркового вещества, имеет более рыхлую структуру по сравнению с коровым веществом.
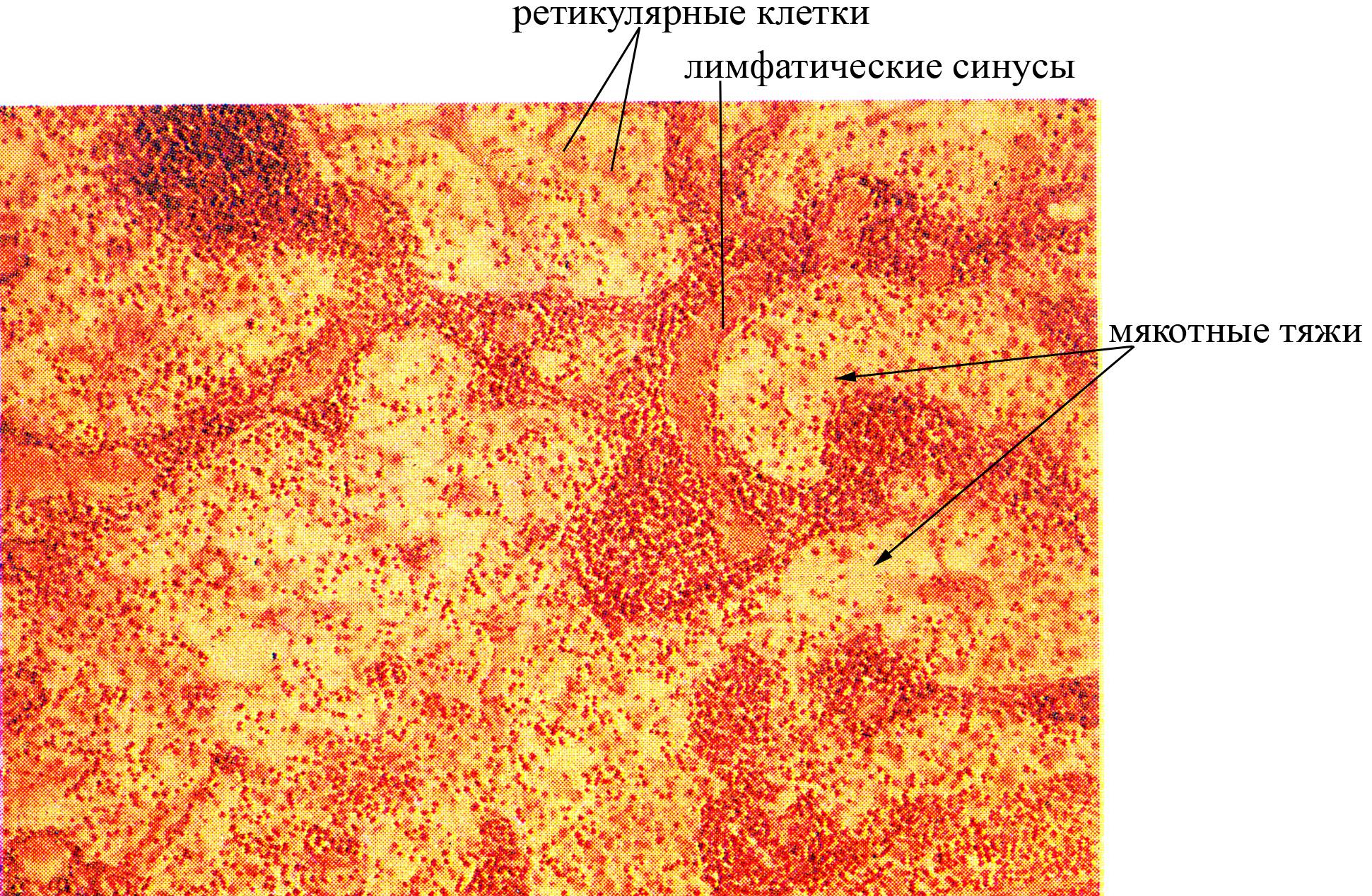
Рис. Гистоструктура мозгового вещества лимфатического узла
В основе коркового и мозгового вещества лимфатических узлов лежит ретикулярная ткань и лимфоидные клетки, находящиеся на разных этапах дифференцировки. Кроме того, и в мозговом, и в корковом веществе лимфатических узлов содержится большое количество макрофагов. Ретикулярная ткань лимфатического узла представлена дендритными ретикулярными клетками и ретикулиновыми волокнами, контактирующими между собой и образующими подобие петлистой сети, в ячеях которой локализованы лимфоидные клетки. Ретикулиновые волокна соединяются не только между собой, но и с коллагеновыми и эластическими волокнами соединительной ткани капсулы узла и его трабекул, что обеспечивает создание прочного волокнистого каркаса внутри лимфатического узла. Лимфоидные клетки коркового и мозгового вещества узла оказываются окруженными со всех сторон дендритными ретикулярными клетками, которые выполняют следующие функции:
антигенпредставляющую (связывают антигены и локализуют их на своей поверхности; благодаря большой суммарной площади своей поверхности (за счет наличия большого количества отростков) они обеспечивают поддержание высоких концентраций антигенов вокруг созревающих клеток лимфоидного ряда)
трофическую по отношению к клеткам лимфоидного ряда (принимают участие в доставке различных веществ из крови к лимфоидным клеткам и в обратном направлении), в связи с чем их образно называют «клетками-няньками» лимфоидных клеток
опорную по отношению к развивающимся лимфоидным клеткам.
Наряду с ретикулярными клетками лимфоидные клетки коркового и мозгового вещества лимфатического узла окружены большим количеством макрофагов, часть из которых обладает способностью к фагоцитозу, а часть – утратила эту способность. Макрофаги, участвующие в фагоцитозе, характеризуются наличием в своей цитоплазме большого количества фагосом, они, с одной стороны, способствуют очистке протекающей через узел лимфы от различных антигенов, а с другой – принимают участие в иммунном ответе, поскольку обеспечивают превращение корпускулярного антигена в молекулярный, который доступен для различения иммунокомпетентными лимфоцитами. Так, макрофаги после фагоцитоза антигенов частично их лизируют, после чего транспортируют на свою поверхность какие-то молекулярные компоненты этих антигенов, где они связываются с рецепторами мембраны макрофага и могут различаться определенными иммунокомпетентными лимфоцитами, коммитированными на данный антиген. Часть макрофагов, находящихся в лимфатическом узле утратила способность к фагоцитозу. Существует предположение, согласно которому такие клетки узла являются «пришлыми» макрофагами, потомками клеток Лангерганса кожи, мигрирующими в узел и приносящими сюда антигены, поступившие в организм через поверхность кожи. Причем эти макрофаги характеризуются большой продолжительностью жизни и специализируются на антигенпредставляющей функции, поскольку фиксируют на своей поверхности различные молекулярные компоненты антигенов, в том числе проникшие через кожу.
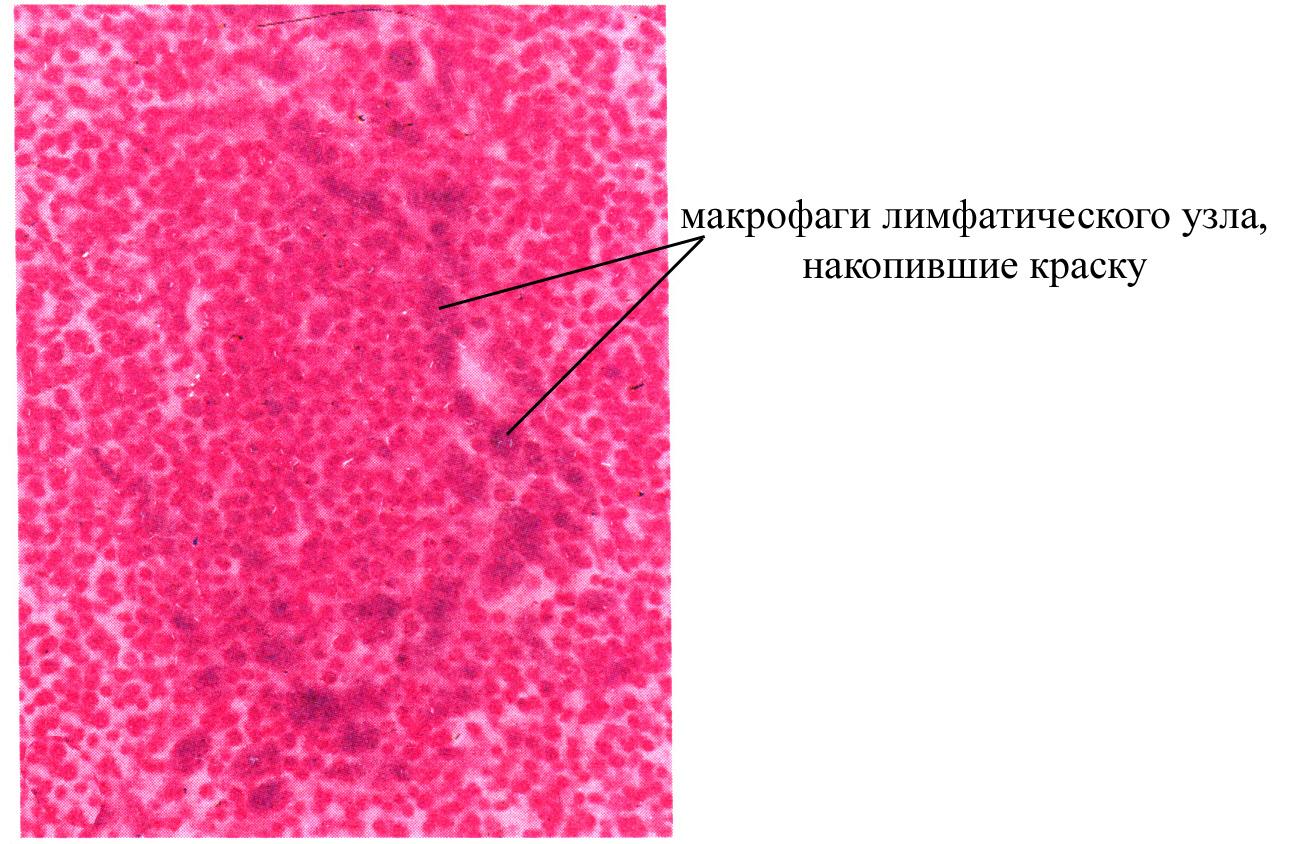
Рис. Накопление краски в макрофагах лимфатического узла (окраска гематоксилин-эозином)
Распределение популяций Т- и В-лимфоцитов в корковом и мозговом веществе лимфатического узла подчиняется определенной закономерности. Так, в периферической части коркового вещества, соответствующей месту локализации лимфатических фолликулов, преобладают в основном В-лимфоциты, а сами эти лимфатические фолликулы являются местом антигензависимой активации В-лимфоцитов, в которой участвуют Т-хелперы и образуются активированные В-лимфоциты. Часть активированных В-лимфоцитов мигрирует в мозговое вещество, где на вершинах мякотных тяжей превращается в плазматические клетки, продуцирующие антитела, а часть – дифференцируется в В-клетки памяти, которые могут оставаться в узле или поступать в кровь и лимфу и мигрировать по организму. Плазматические же клетки, в отличие от В-клеток памяти являются неподвижными и так и остаются в области мякотных тяжей, продуцируя антитела.
Следовательно, лимфатические фолликулы являются местом антигензависимого дифференцирования В-лимфоцитов, которое протекает с участием макрофагов (антигенпредставляющих клеток) и в большинстве случаев – Т-хелперов. Состояние этих фолликулов во многом зависит от проникновения определенных антигенов в узел. В частности, после антигенной стимуляции в некоторых лимфатических фолликулах появляются более светлые зоны, называемые центрами размножения. Они соответствуют месту нахождения крупных лимфоидных клеток, активно делящихся и дающих начало активированным малым лимфоцитам. Поскольку в центре узла содержатся преимущественно крупные предшественники малых лимфоцитов, эта область узла имеет более светлую окраску, обусловленную относительно большим объемом цитоплазмы крупных лимфоидных клеток, по сравнению с активированными малыми лимфоцитами. Периферия же лимфоидных фолликулов представлена скоплениями преимущественно активированных малых лимфоцитов, характеризующихся меньшими размерами и объемом цитоплазмы и поэтому создающих более темную картину в связи с большей плотностью ядер. В отсутствии антигенов в узле большая часть первичных фолликулов не имеет центров размножения. Эти центры отсутствуют также до момента рождения и у «безмикробных» животных. Таким образом, лимфатический узел представляет собой очень динамичную структуру, в которой, в зависимости о проникающих антигенов, происходит возникновение центров размножения в определенных лимфатических фолликулах на фоне временного «молчания» других.
Наряду с заселением В-лимфоцитами периферической части коркового вещества лимфатического узла, многочисленные их популяции встречаются и в мозговом веществе (среди ретикулярной ткани мякотных тяжей). В-лимфоциты мозгового вещества в большинстве своем являются активированными в результате контакта с определенными антигенами малыми лимфоцитами, мигрирующими из коркового вещества узла и находящимися на завершающих стадиях созревания. Они могут превращаться либо в В-лимфоциты памяти, либо в плазматические клетки (как правило, на вершине мякотных тяжей), продуцирующие в основном иммуноглобулины класса G. Наряду с В-лимфоцитами в ячеях ретикулярной ткани мозгового вещества содержаться и макрофаги. Внутри мякотных тяжей коркового вещества проходят артериолы и капилляры, содержащие поры в эндотелии, через посредство которых активированные В-лимфоциты могут поступать в кровоток. Снаружи мякотные тяжи, подобно лимфатическим фолликулам покрыты эндотелиоподобными ретикулярными клетками, лежащими на ретикулярных волокнах.
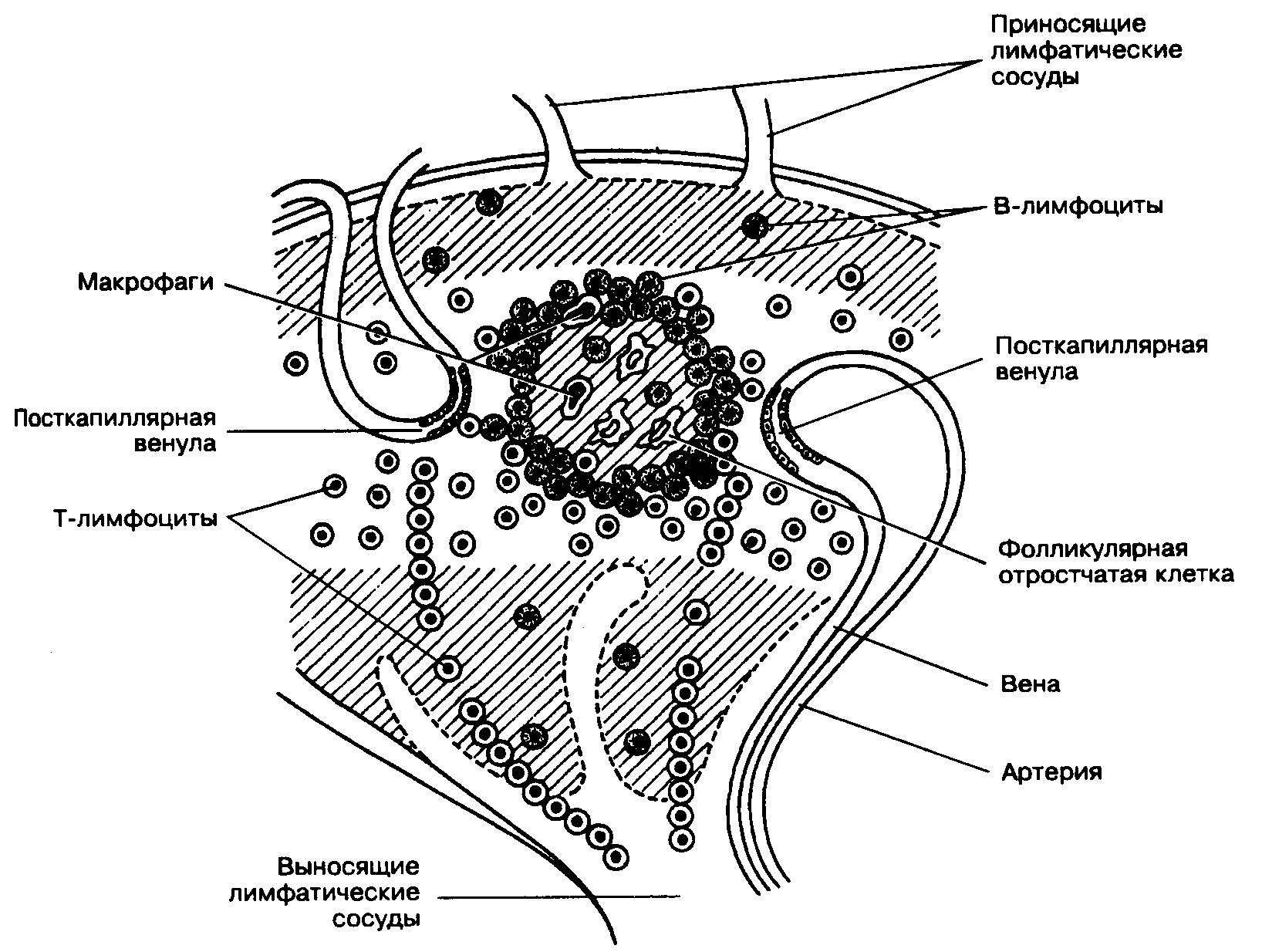
Рис. Схема распределения Т- и В-лимфоцитов в лимфатическом узле

Рис. Локализация паракортикальной (Т-зоны) в лимфатическом узле
Наконец, зона узла, соответствующая средней и внутренней части коркового вещества и граничащая с мозговым веществом, называется тимусзависимой зоной узла, поскольку заселена преимущественно Т-лимфоцитами. Иногда эту зону узла называют еще паракортикальной, считая в таком случае истинной корой узла периферическую его часть, содержащую лимфатические фолликулы (т.е. тимусзависимая зона с одной своей стороны прилежит к периферической части коркового вещества узла, а с другой – контактирует с мозговым его веществом). В этой зоне узла в ячеях ретикулярной ткани преобладают Т-лимфоциты-хелперы, и несколько реже встречаются другие популяции Т-лимфоцитов (в частности, Т-киллеры). Тимусзависимая зона узла получила свое название в связи с тем, что заселяется она малодифференцированными Т-лимфоцитами, приносящимися из тимуса, и после удаления тимуса она запустевает из-за убыли Т-лимфоцитов. В этой зоне происходит антигензависимая пролиферация Т-клеток и дифференцировка в определенные эффекторные клетки (Т-киллеры, Т-хелперы и т.д.). Пересадка аллотрансплантанта вблизи места расположения лимфатического узла сопровождается резким увеличением тимусзависимой зоны, в связи с поступлением в узел чужеродных антигенов аллотрансплантанта и вызванной этим антигензависимой пролиферацией и активацией малодифференцированных Т-лимфоцитов, поступивших из тимуса и коммитированных на данные антигены. Наряду с ретикулярными и лимфоидными клетками в тимусзависимой зоне узла присутствуют и макрофаги, в том числе утратившие способность к фагоцитозу (т.н. интердигитирующие клетки). Интердигитирующие клетки обладают многочисленными пальцевидными отростками, вдавливающимися из одной клетки в другую, они мигрируют с током лимфы из кожи и являются производными внутриэпидермальных макрофагов (клеток Лангерганса), доставляющих в лимфатический узел антигены, проникшие через кожу. Функциональное значение таких интердигитирующих клеток состоит в том, что они специализируются на синтезе гликопротеидов примембранных слоев (т.е. гликопротеидов гликокаликса), которые способны сорбировать и сохранять антигены на плазматических мембранах, а также индуцировать пролиферацию Т-лимфоцитов.
Читайте также: Для чего ткань в опыте опускали именно в раствор йода
Таким образом, распределение Т- и В-лимфоцитов в лимфатических узлах подчиняется следующей закономерности:
В-лимфоциты располагаются преимущественно в наружной части коркового вещества лимфатического узла и в области мякотных тяжей мозгового вещества. Причем В-лимфоциты коркового вещества в большинстве своем представляют собой малодифференцированные лимфоидные клетки, находящиеся на разных стадиях антигензависмой дифференцировки. Тогда как тяжи узла заселены преимущественно зрелыми малыми В-лимфоцитами, заканчивающими свою дифференцировку и превращающимися либо в плазматические клетки (неподвижные клетки, продуцирующие антитела), либо в В-лимфоциты памяти
Т-лимфоциты преобладают в средней и внутренней области коркового вещества узла и представляют собой лимфоидные клетки, находящиеся на разных этапах антигензависимой активации.
Наряду с соединительнотканной стромой, ретикулярной и лимфоидной тканями, типичной морфологической особенностью лимфатического узла являются синусы (в переводе с латинского означает полое пространство), заполненные лимфой (лимфатические синусы). В лимфатическом узле различают следующие синусы:
краевой синус, располагается непосредственно под соединительнотканной капсулой узла, выстлан лимфатическим эндотелием (уплощенные эндотелиальные клетки), не имеющим базальной мембраны, но контактирующим в некоторых местах с коллагеновыми и ретикулиновыми волокнами. Причем со стороны соединительнотканной капсулы эндотелиальная выстилка синуса является сплошной, тогда как со стороны лимфатических фолликулов корокового вещества – прерывистой. В краевой синус лимфатического узла поступает лимфа, приносимая по афферентным лимфатическим сосудам, несущим лимфу из тканей и проникающим в узел со стороны наружной его поверхности (поверхности, противоположной воротам узла). Краевой синус содержит много свободных клеток, преимущественно макрофагов и лимфоцитов, которые могут проникать сюда из подлежащего коркового вещества лимфатического узла. Благодаря многочисленным макрофагам и лимфоцитам краевой синус обеспечивает фагоцитоз различных антигенов, поступающих с лимфой, а, следовательно, принимает участие в начальной фильтрации и очистке лимфы.
корковые и промежуточные корковые синусы, отходящие радиально от краевого синуса и проходящие в толще коркового вещества между ретикулярно-лимфоидными его компонентами, имеющими форму пирамид. В этих синусах в протекающую лимфу попадает еще некоторое количество лимфоцитов из коркового вещества узла
мозговые синусы, являются продолжением синусов коркового вещества в мозговом веществе узла; проходят между мякотными тяжами, по этим синусам лимфа направляется к выносящим лимфатическим сосудам, покидающим узел в области его ворот.

Рис. Краевой синус лимфатического узла (вид при сканирующей электронной микроскопии)
Стенка всех синусов устроена по общему принципу: она образована лимфатическим эндотелием или ретикулярными эндотелиоподобными клетками, не имеет базальной мембраны, укреплена в некоторых местах ретикулиновыми волокнами и характеризуется наличием крупных просветов между своими клетками, позволяющими клетками проходить в полость синуса или в обратном направлении в ткань узла. Обычными типами клеток внутри синусов являются лимфоциты и макрофаги, обеспечивающие очистку лимфы.
Кровоснабжение лимфатического узла осуществляется артериями, входящими в узел в области ворот. Эти артерии и более мелкие отходящие от них сосуды (мелкие артерии, артериолы) проходят в толще узла в составе соединительнотканных трабекул и мякотных тяжей и достигают коркового вещества узла. Причем артериолы до уровня лимфатических фолликулов в корковом веществе, как правило, не доходят, а разветвляются на капиллярную сеть на границе коркового и мозгового вещества. Таким образом, в периферической части коркового вещества лимфатического узла кровоснабжение обеспечивают в основном капилляры, которые переходят в посткапиллярные венулы. Посткапиллярные венулы следуют в корковом веществе в противоположном по отношению к капиллярам направлении (а именно в направлении из коркового вещества в мозговое, т.е. кровеносная сеть совершает обратный ход и возвращается в мозговое вещество). Посткапиллярные венулы переходят в мозговое вещество узла, где впадают в более крупные вены. Венозная кровь из лимфатического узла выносится, как правило, по общей его вене, покидающей узел в области ворот. Важной особенностью посткапиллярных венул коркового вещества лимфатического узла является относительно большая высота эндотелиальных клеток (гораздо большая по сравнению со многими другими венулами организма), в связи с чем такой эндотелий называют кубическим и даже высоким призматическим, а также наличие большого количества пор между эндотелиальными клетками. Функциональное значение такого высокого призматического эндотелия посткапиллярных венул состоит в том, что его клетки, после того как лимфоцит проник между ними, со стороны просвета сосуда (выше места его нахождения лимфоцита в стенке венулы) смыкаются и препятствует просачиванию жидкости. Таким образом, высокий призматический эндотелий венул осуществляет хоминг лимфоцитов, препятствуя при этом выходу плазмы из венулы в узел.

Рис. Миграция лимфоцитов сквозь стенку посткапиллярной венулы (схема)

Рис. Миграция лимфоцитов сквозь стенку посткапиллярной венулы (сканирующая электронная микроскопия)
Лимфоциты, покинувшие посткапиллярные венулы, в большинстве своем поступают в лимфатические синусы или лимфоретикулярную ткань коркового и мозгового вещества, где могут контактировать с различными антигенами, принесенными лимфой из тканей, и в результате такого контакта активироваться. Кроме того, часть, вышедших из посткапиллярных венул лимфоцитов с током лимфы выносятся из узла и возвращаются обратно в кровь (рециркуляция лимфоцитов).
Развитие лимфатических узлов в эмбриональный период. Лимфатические узлы возникают в конце 2-го – начале 3-го месяца внутриутробного развития из мезенхимных клеток, образующих скопления в определенных областях кровеносных и лимфатических сосудов. Мезенхимные клетки дают начало соединительнотканной капсуле узла и трабекулам, а также ретикулярной ткани. К 16-й неделе зачатки лимфатических узлов начинают заселяться лимфоцитами. Причем В-лимфоциты вселяются раньше Т-лимфоцитов, главным образом в центральные участки лимфатического узла (будущее мозговое вещество), а затем в самый поверхностный (в дальнейшем – корковое вещество) слой. Т-лимфоциты вселяются в промежуточную зону между корковым и мозговым веществом (паракортикальную или тимусзависимую зону). На 16-й неделе в лимфатическом узле образуются лимфоидные фолликулы и мозговые тяжи, а также появляются ретикулярные волокна. к концу эмбриогенеза в лимфатических узлах заканчивается формирование всех структур – коркового вещества с лимфатическими фолликулами, мозговых тяжей, лимфатических синусов, Т- и В-зон. Однако до момента рождения (и соответственно начала контакта лимфоцитов узла с антигенами в его лимфатических фолликулах отсутствуют центры размножения, а плотность Т-зоны оказывается гораздо меньше, чем в постнатальном периоде).
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом