
Перед обсуждением процессов нормальной миелинизации в головном мозге человека необходимо понимать строение миелина и его функции в центральной нервной системе. Миелин имеется и в центральной, и в периферической нервных системах. В ЦНС он находится преимущественно в белом веществе (хотя некоторые его количества имеются и в сером), как раз придавая ему такой цвет. Миелин работает по типу электрического изолятора: скорость проведения потенциала действия возрастает в 10-100 раз по сравнению с немиелинизированными волокнами. Юлия Эдгар и Джессика Гарберн (2004) показали, что отсутствие главного белка миелина (PLP/DM20) в олигодендроцитах приводит к серьезным нарушениям аксонального транспорта у мышей с моделью наследственной спастической параплегией. Также было выяснено, что миелин, вероятно, играет роль в регуляции как состава ионов, так и объема жидкости вокруг аксона.
Миелинизация – процесс образования билипидного миелинового слоя вокруг аксона. Данный процесс обеспечивает в дальнейшем быструю передачу информации, необходимую для когнитивной, поведенческой, эмоциональной функций. Миелинизация начинается во время эмбрионального периода и продолжается после рождения.
Миелин является модифицированным расширением отростков олигодендроглиальных клеток. Олигодендроцит является ключевой клеткой в миелинизации ЦНС и является преобладающим типом нейроглии в белом веществе. Миелиновая оболочка состоит из множественных сегментов миелина, которые обвиваются вокруг аксона. Данная оболочка способна проводить потенциал действия с его увеличением по аксону благодаря перехватам Ранвье, которые представляют собой немиелинизированные участки, содержащие натриевые ионные каналы – они и ответственны за ускорение проведения ПД по аксону. Миелин также находится в некотором симбиозе с аксоном. Миелин участвует в метаболических превращениях своих компонентов и содержит большое количество миелиновых ферментов. Миелин также играет роль в переносе ионов, что способствует буферизации ионов вокруг аксона.
Один олигодендроцит может обеспечивать миелиновой оболочкой до 40 волокон. Миелин примерно на 70% состоит из липидного компонента и на 30% – из белкового. Основной белок миелина (MBP) составляет 30% от всей белковой фракции, протеолипидный белок (PLP) – 50%, а фосфодиэстеразы циклических нуклеотидов – 4%. Липиды миелина в своем составе содержат холестерол, фосфолипиды, гликосфинголипиды.
Миелин и МРТ
Не существует МРТ методики или технологии, которая могла бы визуализировать напрямую миелиновый бислой. Миелин оценивается качественно на основании Т1- и Т2-ВИ, МР-спектроскопии, диффузно-тензорной визуализации (DTI). В клинической практике традиционная анатомическая визуализация является основой из-за своей легкости выполнения. Количественная оценка миелина может быть получена в многокомпонентном релаксационном (MCR или multicomponent relaxation) анализе. MCR-анализ – это объемно-взвешенное суммирование микроскопических компартментов воды. С помощью этого метода определяются два домена воды: медленно-релаксирующий домен, включающий свободную внутри- и внеклеточную воду, и быстро-релаксирующий домен, включающий в себя воду, заключенную в липидном бислое миелина.
На данный момент стандартные МРТ-последовательности не способны специфично подсчитывать количество миелина. Они позволяют оценить изменения в плотности и размерах аксонов, изменения в мембранной структуре, а именно содержание белков, липидов и воды. DTI – ненадежный показатель общего количества миелина, но все же дает информацию о его возможных изменениях.
Джеймс Баркович выделяет две отдельные популяции молекул воды, которые играют главную роль в формировании сигнала от миелина на МРТ – это те молекулы, которые находятся в миелиновой оболочке, и молекулы, находящиеся вне ее. На анатомических изображениях миелин имеет гиперинтенсивный сигнал по отношению к серому веществу на Т1- и гипоинтенсивный на Т2-ВИ. На Т1-ВИ гиперинтенсивность относительно коры определяется вероятнее всего наличием большого количества гликолипидов (особенно галактоцереброзидов) и холестерола в миелиновой оболочке. На Т2-ВИ гипоинтенсивность обусловлена уменьшенным содержанием воды.
Какая ткань имеет миелин
Взаимодействия между глиальными и нервными клетками отчетливо проявляются в процессах развития и структурной организации нервных волокон. Нервным волокном называется отросток нервной клетки, окруженный глиальной оболочкой.
Непосредственно сам отросток называют еще осевым цилиндром, а клетки глиальной оболочки — нейролеммоцитами. Различают миелиновые (мякотные) и безмиелиновые (безмякотные) нервные волокна.
В безмиелиновых нервных волокнах отростки нервных клеток погружены в углубления на поверхности нейролеммоцитов, имеющих вид желоба. Погруженный в тело глиальной клетки нервный отросток ограничен как собственной плазмолеммой, так и внешней мембраной нейролеммоцита. Он как бы подвешен на двухлистковой ее складке. Эти складки мембран (своеобразные ультраструктурные «брыжейки») называют мезаксонами. Безмиелиновые волокна могут включать несколько осевых цилиндров.
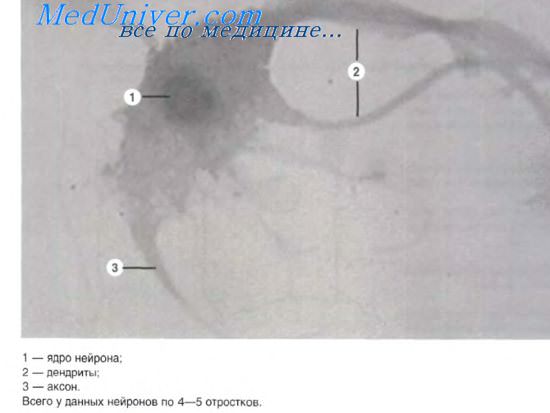
Миелиновое нервное волокно состоит из нервного отростка и нейролеммоцитов (шванновских клеток). Осевой цилиндр не просто погружен в цитоплазму нейролеммоцита, а окружен спиральной слоистой оболочкой (миелином), образованной наматыванием мезаксонов нейролеммоцитов при их вращении вокруг отростка нервной клетки. В миелиновой оболочке обнаружены липиды, щелочной белок миелина, маркерный белок S100 и др.
Высокое содержание липидов (почти 2/3 массы миелина) выявляется при обработке препаратов четырехокисью осмия, окрашивающей миелиновую оболочку в темно-коричневый цвет. По ходу миелинового волокна имеются сужения — узловые перехваты (перехваты Ранвье). Они соответствуют границе смежных нейролеммоцитов. Каждый межузловой сегмент оболочки волокна представлен одним нейролеммоцптом. Миелиновые волокна толще безмиелиновых. Скорость проведения нервного импульса по ним составляет 5-120 м/с, тогда как по безмиелиновым волокнам импульс проводится со скоростью 1-2 м/с.
Сложные взаимоотношения между нервными и глиальными клетками складываются при формировании чувствительных нервных окончаний (рецепторов) и двигательных нервных окончаний (эффекторов).
Нервные окончания — концевой аппарат нервных волокон, формирует межнейрональные контакты, или синапсы, рецепторные (чувствительные) окончания и двигательные (эффекторные) окончания.
Синапс (от synapsis — соединение) — специализированный для передачи нервных импульсов контакт между двумя нейронами или между нейроном и эффектором. Процессы возбуждения нейронов, возникновение импульсов и распространение их по отросткам связаны с изменениями в плазмолемме. Она является структурной основой возникновения и передачи потенциалов действия. Плазмолемма имеет существенные особенности строения и функции в участках, входящих в состав синапсов.
Межнейрональные синапсы бывают нескольких видов: аксосоматические (между аксоном одного нейрона и телом другого нейрона); аксодендритические (между аксоном одного нейрона и дендритом другого нейрона); аксоаксональные (между аксонами двух нейронов). Описаны также синапсы соматосоматические, дендродендритические и др.
Все синапсы по механизму передачи импульсов между нервными клетками подразделяются на 3 типа: синапсы с химической передачей, электротонические и смешанные синапсы. Типичный синапс с химической передачей состоит из пресинаптической и постсинаптической частей, а также синаптической щели. Пресинаптическая часть включает концевое расширение аксона, ограниченное пресинаптической мембраной. Специфическими структурами этой части являются синоптические пузырьки, содержащие нейромедиаторы. Пузырьки бывают со светлым и электронно-плотным содержимым и называются в связи с этим агранулярными и гранулярными.
По форме они подразделяются на круглые и уплощенные. На внутренней поверхности пресинаптической мембраны расположены конусовидные электронно-плотные образования — пресинаптические уплотнения. В цитоплазме пресинаптической части имеются митохондрии. Синаптическая щель размером 20-30 нм содержит филаменты, связывающие наружные слои плазмолеммы контактирующих нейронов.
Читайте также: Набивная ткань что это такое 4 буквы
Постсинаптическая часть в составе плазмолеммы второго нейрона имеет рецепторы к медиатору, который выделяется в синаптическую щель при деполяризации мембраны первого нейрона. Внутренняя поверхность постсинаптической мембраны характеризуется наличием электронно-плотного слоя цитоплазмы — постсинаптические уплотнения.
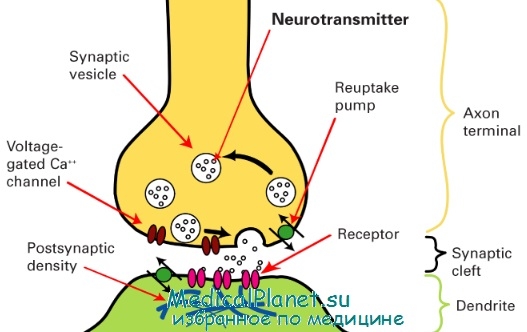
Схема строения синапса
ОСНОВНОЙ БЕЛОК МИЕЛИНА. СТРОЕНИЕ, СВОЙСТВА, ФУНКЦИИ, РОЛЬ В ДИАГНОСТИКЕ ДЕМИЕЛИНИЗИРУЮЩИХ ЗАБОЛЕВАНИЙ
Ключевые слова: основной белок миелина, онтогенез, рассеянный склероз, гидроцефалия, опухоли головного мозга, демиелинизирующие заболевания.
ВВЕДЕНИЕ. Молекулярная организация миелина. Основная информация о структуре миелина получена с помощью рентгено-структурного анализа и электронной микроскопии [1–10]. Уникальной морфологической особенностью миелина является то, что он формируется в результате спирального обвития отростков олигодендроглиоцитов в центральной нервной системе и шванновских клеток на периферии, вокруг аксонов нейронов [2, 7, 10–13]. Таким образом, миелин представляет собой своеобразную мембрану, состоящую из липидного бислоя и белков, связанных с ним. В образовании миелиновой оболочки и структуре миелина ЦНС и периферической нервной системы (ПНС) имеются отличия. При формировании миелина ЦНС один олигодендроглиоцит имеет связи с несколькими сегментами миелина нескольких аксонов; при этом к аксону примыкает отросток олигодендроглиоцита, расположенного на некотором расстоянии от аксона, а внешняя поверхность миелина соприкасается с внеклеточным пространством. Шванновская клетка при образовании миелина ПНС формирует спиральные пластинки миелина и отвечает лишь за отдельный участок миелиновой оболочки между перехватами Ранвье. Цитоплазма шванновской клетки вытесняется из пространства между спиральными витками и остается только на внутренней и наружной поверхностях миелиновой оболочки [2, 12].
Среди белков миелина выделяют так называемые внутренние (intrinsic) и внешние (extrinsic) белки [14]. Первые прочно связаны с мембраной, проходя сквозь нее, в то время как другие, расположенные поверхностно, связаны слабее. Подобная мембрана является асимметричной по химическому составу и электрическому заряду. Ее экстрацеллюлярная поверхность богата углеводными остатками гликопротеинов и гликолипидов, при этом С-конец гликопротеинов находится на цитоплазматической стороне мембраны, тогда как полисахаридный остаток экспонирован на экстрацеллюлярной поверхности. Расстояние между разнонаправленными гидрофильными группировками липидов в мембране миелина составляет 4,5-5,0 нм, в то время как расстояние между соседними витками спирали — 3,0-5,0 нм. Толщина мультиламеллярного образования, сформированного исключительно липидами, составляет 1,5-3,0 нм [2, 14–16].
Процесс формирования миелина отростками глиальных клеток сопровож-дается вытеснением цитоплазмы таким образом, что цитоплазматические поверх-ности мембраны плотно соприкасаются друг с другом, образуя так называемую главную плотную линию (major dense line). Плотный контакт наружных поверхностей мембран, образующихся при спиралевидном обвитии отростков миелинобразующих клеток вокруг аксонов нейронов, способствует формирова-нию, так называемой межпромежуточной линии (interperiod line) [2, 12]
Одной из биохимических характеристик, которая отличает миелин от других биологических мембран, является высокое соотношение липид/белок. Белки составляют от 25 до 30% массы сухого вещества миелиновой оболочки. На долю липидов приходится приблизительно 70-75% от сухой массы белого вещества ЦНС млекопитающих; в миелине спинного мозга соотношение липиды:белки выше [12, 15]. Из общего количества липидов на долю холестерола приходится около 28%, 43% – на фосфолипиды и 29% составляют галактолипиды. Известно, что липиды оказывают существенное влияние на конформационные характеристики белков, входящих в состав мембраны миелина; последние в свою очередь влияют на свойства липидов [2, 12].
Основной функцией миелина является быстрое проведение нервного импульса по аксонам, которые он окружает. Мембраны клеток, формирующих миелин, плотно соприкасаются, что обеспечивает высокое сопротивление и малую емкость, обеспечивая, таким образом, аксону эффективную изоляцию и предотвращая продольное распространение импульса. Миелин прерывается только в области перехватов Ранвье, которые встречаются через правильные промежутки длинной примерно 1 мм. В связи с тем, что ионные токи не могут проходить сквозь миелин, вход и выход ионов осуществляется лишь в области перехватов. Это ведет к увеличению скорости проведения нервного импульса. Таким образом, по миелинизированным волокнам импульс проводится приблизительно в 5-10 раз быстрее, чем по немиелинизированным [2, 12].
Помимо передачи нервного импульса, миелин участвует в питании нервного волокна, а также выполняет структурную и защитную функции
Строение, физико-химические свойства. При электрофоретическом анализе в ПААГ с SDS экстрактов миелиновой оболочки человека [1, 17, 18] определяется до 10 полос. Основную полосу (около 50% всех белков), составлял протеолипидный белок (PLP) с молекулярной массой 30 кДа.
Вторыми в количественном соотношении (около 30%) были белки, входящие в так называемую группу основных белков миелина (ОБМ), соответствующие молекулярным массам от 17 до 21,5 кДа. Данную группу составили 3 изоформы ОБМ с молекулярными массами 21,5 кДа, 18,5 кДа и 17,5 кДа. Первая из них кодируется 7 экзонами; в результате делеции 2 экзонов синтезируется белок молекулярной массой 18,5 кДа. При кодировании же изоформы массой 17,5 кДа были подвержены делеции 5 экзонов [18–21]. Показано, что ген ОБМ человека расположен на 18 хромосоме и имеет 3 промоторные области, с которых начинается считывание информации [21].
У различных видов животных было также показано наличие нескольких изоформ данного белка [23, 24]. Так, ОБМ крысы включает в себя 4 изоформы с молекулярными массами 21,5, 18,5, 17,0 и 14,0 кДа. Интересно, что изоформы ОБМ массами 21,5 и 18,5 кДа кодируются экзонами, комплементарными человеческим, за небольшим исключением, касающимся незначительных перестановок последовательностей. В изоформе ОБМ с массой 17,0 кДа делеции подвержен 6 экзон. Делеция 2 и 6 экзонов наблюдается при кодировании изоформы белка массой 14,0 кДа. Эти две изоформы ОБМ крысы, таким образом, не имеют человеческих аналогов. Кроме того, доказано наличие изоформ ОБМ с молекулярными массами 21,5, 17,0 и 14,0 кДа в ткани ЦНС мыши [25]. Низкомолекулярные изоформы белка образуются путем делеции участка хромосомы, кодирующего аминокислотные последовательности в области С-конца. Недавние исследования [26] показали наличие ОБМ с молекулярной массой 21,5 кДа в мозге барана, при этом было доказано, что он не является предшественником низкомолекулярных изоформ белка. Кроме того, ОБМ с молекулярной массой 13,5 кДа был идентифицирован в мозге серебряного карася [27].
В настоящий момент полностью установлена аминокислотная последовательность [17, 26, 28, 29] 18,5 кДа изоформы ОБМ человека [30], морской свинки [31], свиньи [32]. Проводятся исследования по определению аминокислотной последовательности ОБМ кролика, быка, обезьяны [33].
Интересен тот факт, что человеческий ОБМ имеет вариации последовательностей аминокислот в 46 и 47 положении. Здесь может находиться глицин, как самостоятельно, так и в сочетании с серином [30]. Согласно более ранним исследованиям, у больных с рассеянным склерозом отмечалась замена серина на глицин в 44-49 положениях [34].
Показано, что среди млекопитающих имеется значительная степень гомологии между аминокислотными последовательностями ОБМ [35, 36].
Рядом авторов была показана высокая степень гомологии – порядка 80-90% [29, 35, 36] аминокислотной последовательности ОБМ у различных видов животных. Так, аминокислотные последовательности ОБМ человека и быка имеются различия аминокислотных остатков лишь в нескольких положениях, в то время как ОБМ крысы отличаются от ОБМ человека и быка положением 40 аминокислотных остатков в середине полипептидной цепи (от 118 до 157 остатка).
ОБМ содержит необычайно высокий процент (приблизительно 25%) основных аминокислот (аргинин, лизин и гистидин), равномерно распределенных по всей полипептидной цепи, что и обусловливает очень высокую изоэлектрическую точку ОБМ (рI=12-13) [37, 38]. Основная изоформа ОБМ с молекулярной массой 18,5 кДа) в дальнейшем может быть субфракционирована в щелочной среде по зарядам. Подобная неоднородность заряда увеличивается в результате фосфорилирования и дезаминирования in vivo. В процессе дальнейшего выделения белка происходит дополнительное дезаминирование и деградация в области С-конца. Такая гетерогенность зарядов может варьировать у различных индивидуумов в зависимости от возраста и патологического состояния организма [39].
Фосфорилирование ОБМ. Процесс фосфорилирования является основным источником гетерогенности. Процесс фосфорилирования ОБМ в оптическом нерве крысы возрастает в процессе развития [40]. Фосфорилирование может способствовать приобретению молекулой белка гидрофильных свойств и уменьшать величину положительного заряда. Таким образом, фосфорилированная форма ОБМ должная в наименьшей степени взаимодействовать с анионными фосфолипидами. Это отражается в снижении способности фосфорилированных форм белка к индукции агрегации пузырьков [41]. Кроме этого, и способы модификации, обусловливающие изменения его заряда, влияют на данную способность, а также изменяют организацию липидного слоя [42].
Наличие в миелине протеинкиназной и фосфорилазной активности объясняют способностью регулировать чрезмерное фосфорилирование ОБМ. Протеинкиназа миелина, выделенного из мозга человека, способна активироваться как кальцием или сАМР [43], так и фосфатидилинозитол-4-фосфатом [44]. Наличие высоких концентраций фосфатидилинозитолкиназы отмечается в миелине, выделенном из головного мозга быка [45].
Данный фермент может быть вовлечен в регуляцию механизма контроля чрезмерного фосфорилирования белка. Остатки белка, подвергающиеся фосфорилированию в условиях in vitro, зависят от вида протеинкиназы [46]. Протеинкиназа С важна в качестве регулятора клеточного числа. ОБМ является благоприятным субстратом для действия данного фермента [47]. Некоторые участки этого белка являются специфическими ингибиторами активности протеинкиназы С. Так, было продемонстрировано, что деградация миелина может возрастать при ослаблении регуляторного контроля, обеспечиваемого данным ферментом. Миелиногенез осуществляется при адгезии олигодендроцита к субстрату. Этот процесс сопутствует активации фосфорилирования ОБМ протеинкиназой С [48]. Кроме того, процесс фосфорилирования ОБМ также регулируют ганглиозиды [49]. Фосфорилирование, катализируемое протеинкиназой С, стимулируется ганглиозидами; в то время, как фосфорилирование других участков белка, катализируемое сАМР-зависимой протеинкиназой, ими тормозится. Ганглиозиды подавляют также активацию протеинкиназы С диацилглицеролом. Участки молекулы ОБМ, подвергающиеся фосфорилированию in vitro, были выявлены методом высокоэффективной хроматографии высокого давления. Фосфорилирование in vivo происходит по-разному у различных видов млекопитающих, так, у быка, оно осуществляется в 97 и 165 положениях; у кролика – в 7, 56, 96, 113, 163 положениях. Кроме того, было выявлено, что в развивающемся мозге мыши, процессы фосфорилирования начинаются на самых ранних стадиях [50], что подтверждает предположение о непосредственном их участии в механизмах миелинизации.
Метилирование ОБМ. Другое преобразование белка, происходящее посттранскрипционно, заключается в метилировании одиночного аргининового остатка в 107 положении. Процесс катализируется специфическим ферментом аргинин-метилтрансферазой. Экспрессия данного фермента регулируется тиреоидными гормонами [51].
Степень метилирования ОБМ значительно варьирует в процессе развития. На исключительную важность метилирования ОБМ указывает тот факт, что инициация демиелинизации возможна при ингибировании биосинтеза S-аденозил-L-метионина циклолейцином, а формирование компактной структуры миелина подавляется синефунгином – ингибитором метилтрансферазы. S-аденозил-L-метионин является донором метильных групп для аргининового остатка ОБМ [51]. У мышей линии jimpy, характеризующихся гипомиелинизацией, отмечается более низкий, по сравнению с нормальными, уровень ОБМ-специфичной аргинин-N-метилтрансферазы. Процесс миелинизации не может быть успешно завершен при нормальном уровне синтеза ОБМ потому, что метилирование белка не протекает в достаточной степени и становится невозможным его интеграция в миелиновую мембрану. Недавние исследования показали, что у этих мышей страдает и уровень синтеза ОБМ, а также полностью отсутствует синтез PLP. Отсутствие последнего может критически влиять на способность ОБМ внедряться в миелиновую мембрану [52].
У другой линии мутантных мышей shiverer, нарушение процесса миелинизации происходит в результате иных механизмов [53]. Мыши shiverer являются мутантными аутосомными рецессивными мышами, с дефицитом содержания миелина в ЦНС. Данная мутация заключается в делеции 20 т.п.о. гена, которая приводит к отсутствию ОБМ [21, 54]. Гомозиготные мыши являются отличной поведенческой моделью, у которых наблюдается около 12 дня постнатального развития дрожание, конвульсии и ранняя смерть. Морфологический анализ показал практически полное отсутствие миелина в ткани мозга со слабо миелинизированными аксонами. Миелин у этих мышей был аномальным, представлял собой неплотную оболочку, что косвенным образом свидетельствовало о том, что ОБМ необходим для формирования плотности оболочки [55]. Напротив, миелин ПНС у мышей shiverer был качественно и количественно нормальным, с нормальной толшиной и структурой, относительно небольшие аномалии миелина наблюдались на рентгенограммах седалищного нерва у взрослых мышей shiverer [56]. Биохимические изучения миелина у мышей shiverer показали некоторые изменения в количестве липидов. Однако, отсутствие у таких мышей ОБМ не нарушает формирование компактного миелина в ПНС, которое, возможно, компенсирует другой белок миелина – Pо [57].
Известно, что все изоформы ОБМ могут подвергаться ацетилированию N-конца. Однако и свободный С-конец способен подвергаться действию карбоксипептидаз. ОБМ подвержен действию различных протеолитических ферментов. Гидролиз белка под действием катепсина D ускоряется в присутствии кислых липидов. Этот процесс может быть ограничен как ферментативно, так и неферментативно при деградации ОБМ в процессе изоляции. Этот метод основан на хроматографическом удалении примесей протеолитических ферментов и исключении неферментативной деградации при высоких значениях рH и температур [58].
Показано, что минорные фракции ОБМ содержат ковалентно связанный в области серинового остатка в 54 положении фосфатидилинозитолбисфосфат. В процессе экспериментального аллергического энцефаломиелита степень связывания фосфатидилинозитолбисфосфата сначала снижается, а затем возрастает [59].
Изучение структурной организации молекулы ОБМ в растворе проводились с помощью ЯМР-спектроскопии, флуоресцентной спектроскопии, калориметрии, а также с применением иммунологических методов анализа [60–62].
В растворе ОБМ имеет тенденцию к самоассоциации, которая играет важную роль в поддержании компактной структуры миелина. Белок в растворе представлен смесью мономер-гексамерных форм с преобладанием цепей с b-структурой [63]. Преобладание b-структуры и формирование a-структуры индуцируется в фрагментах ОБМ при переносе из водной фазы в трифторэтанол [64].
Взаимодействие ОБМ с липидами. ОБМ как мембранный компонент, имеет значительное сродство к липидам, с которыми в комплексе и выделяется [8, 65]. Кроме того, будучи поликатионом, ОБМ формирует наиболее стабильные комплексы с анионными липидами [2, 66]. Но, несмотря на то, что степень связывания ОБМ значительно выше с кислыми липидами, она достаточно выражена и в отношении цвиттер-ионов, особенно – со сфингомиелином. ОБМ также взаимодействует с фосфатидилэтаноламином при рН 7,2, где этот липид находится преимущественно в цвиттер-форме. Удивительна способность ОБМ формировать нерастворимые комплексы с SDS как выше, так и ниже изоэлектрической точки [67].
При формировании комплексов ОБМ с основной массой анионных фосфолипидов, играют роль в равной степени как гидрофобные, так и электростатические взаимодействия [68]. Наиболее выраженное влияние на стабилизацию липидов ОБМ оказывает при интеграции в липидные хвосты [69, 70].
ОБМ также способствует освобождению глюкозы из многослойных липосом, содержащих GM4 ганглиозид, который является специфическим маркером миелина и олигодендроглии в ЦНС [71].
Как было показано методом рентгеноструктурного анализа, при формировании миелина происходит погружение большей части ОБМ в слой липидных головок. Известно, что ОБМ ускоряет формирование мультислоя фосфатидилглицерола [72] и индуцирует формирование многослойной мембранной структуры с двойной периодичностью бислоя. Холестерол может увеличивать связывание протеина с поверхностью бислоя, открывая каналы между диацил-липидными головками [73].
Smith предположил, [74] а работы Yohg [75] и Cheifetz [41] подтвердили, что ОБМ является стимулятором везикулярной агрегации, которая служит моделью молекулярных взаимодействий, имеющих важное значение для формирования компактной структуры миелина и его стабилизации в мультиламелярной структуре мембраны. ОБМ способен индуцировать агрегацию пузырьков, содержащих только фосфатидилхолин, однако степень агрегации значительно усиливается кислыми фосфолипидами. Агрегация фосфолипидных пузырьков под влиянием ОБМ значительно усиливается в присутствии алифатических альдегидов [2]. Высказываются предположения, что возрастание уровня алифатических альдегидов может быть связано с демиелинизирующими заболеваниями [76]. Холестерол увеличивает способность ОБМ к стимуляции агрегации пузырьков фосфатидилсерина [77].
Brady et. al. [72], а также Sridhara et. al. [78] показали некоторые различия в свойствах ОБМ, выделенного из нормального миелина и ОБМ, выделенного из ткани мозга больных рассеянным склерозом (РС). ОБМ нормального миелина более активен в отношении индукции формирования многослойной структуры с фосфатидилглицеролом [72] и инициации агрегации везикул фосфатидилхолина [78]. Независимо от источника выделения ОБМ (нормальный миелин или миелин при РС), было показано, что компоненты белка, имеющие одинаковый заряд, в равной степени способны индуцировать формирование липидного мультислоя. Также было показано, что отличия ОБМ из нормального миелина и из миелина при РС, вызваны различием зарядов изомеров, входящих в состав ОБМ [66].
ОБМ в филогенезе и онтогенезе. Как филогенетически, так и онтогенетически, появление ОБМ может быть связано с дифференцировкой олигодендроглиоцитов и шванновских клеток. В процессе эволюции нервной системы позвоночных дифференцировка глии на клетки двух типов совпадает с появлением миелинизированных волокон, а также с ограничением способности к функциональной регенерации.
Сравнительные исследования ОБМ из ткани мозга различных биологических видов животных показали значительное межвидовое сходство. ОБМ выделен и очищен, а также описаны его свойства из ткани мозга человека, быка, крысы, мыши, морской свинки [1, 26, 29, 30, 32].
Обнаружение филогенетического сходства гликопротеинов у различных биологических видов дало основание провести исследование этого белка в процессе онтогенеза. Так, Grever и et. al. [11] проводили с помощью иммуногистохимического и Northern blott анализа количественное определение ОБМ в ткани спинного мозга плодов человека с 12 до 24 недели гестации. Развитие структуры миелина исследовали с помощью электронной микроскопии. Тридцать восемь образцов спинного мозга плодов были получены после прерывания внематочной беременности. В период между 12 и 18 неделями гестации показано 15,8-кратное увеличение синтеза ОБМ мРНК. С 18 по 24 недели гестации синтез ОБМ мРНК увеличился в 2,2 раза.
Параллельно проведенный иммуноблот-анализ показал 90,5-кратное увеличение ОБМ (от 0,147 нг/мг до 13,3 нг/мг) между 12 и 18 неделями гестационного развития и приблизительно 11,5-кратное увеличение между 18 и 24 неделями гестации (с 13,3 до 154 нг/мг ткани). Иммуноцитохимический анализ также показал повышение степени окрашивания на ОБМ с увеличением гестационного возраста. У плода с гестационным возрастом 12 недель ОБМ определялся во всех трех боковых канатиках спинного мозга, с 18 недель гестационного развития ОБМ экспрессировался во всем белом веществе мозга, за исключением боковых кортикоспинальных путей и ростральных уровней fasciculus gracilis. Авторы также исследовали структуру миелина с помощью электронной микроскопии. В 12 недель гестации миелиновая оболочка представляла собой редкую неуплотненную пластину, в 18 недель гестации наблюдались отдельные точки компактного миелина, которые экспрессировали ОБМ, в 24 недели гестации компактный миелин наблюдался во всем белом веществе спинного мозга. Таким образом, авторами было продемонстрировано количественное увеличение экспрессии ОБМ, связанное с формированием миелина в течение второго триместра беременности.
Weidenheim и соавт. [79, 80] с помощью иммуногистохимического анализа определяли ОБМ в ткани спинного мозга (в цервикальном, грудном и люмбосакральном уровнях) и стволовой части головного мозга эмбрионов человека на протяжении первого и второго триместров беременности. С 9-10 недель гестации ОБМ-позитивные олигодендроглиоциты наблюдались по периферии эпиндимы. В переднем и боковом канатиках экспрессия ОБМ олигодендроглиоцитами отмечалась с 10-12 недель гестационного развития. В заднем канатике ОБМ детектировался на более поздних сроках гестационного развития, чем в антилатеральном белом веществе. ОБМ с 10 недели гестации обнаруживался в основном по срединной линии ствола мозга и экспрессировался латеральнее на протяжении второго триместра беременности. Таким образом, авторы заключили, что ОБМ присутствует с 10 недель гестации антилатерально в цервикальной области спинного мозга и срединной линии стволовой области мозга и экспрессируется в рострально-хвостовом направлении от антилатеральной к задней области. Исключением, однако, является область fasciculus gracilis, которая имела большее количество ОБМ-положительных клеток на поясничном уровне, чем в более ростральных областях.
Аналогичные результаты были получены и другими исследователями [13, 81, 82].
Таким образом, приведенные выше факты свидетельствуют об общем явлении эмбриогенеза человека: начало синтеза нейроспецифических антигенов приходится на 10-18 недели гестации.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом
- Правообладателям
- Политика конфиденциальности