
4) эластичная базальная мембрана = толстые эластичные волокна, расположенные вдоль сосуда + коллагеновые волокна по окружности сосуда + поперечные связи.

Рис.7.4. Зависимость напряжения σ
стенок кровеносных сосудов
Механические свойства кровеносных сосудов определяются главным образом свойствами коллагена, эластина и гладких мышечных волокон. Содержание этих составляющих сосудистой ткани изменяется по ходу кровеносной системы: отношение эластина к коллагену в общей сонной артерии – 2:1, а в бедренной артерии – 1:2. С удалением от сердца увеличивается доля гладких мышечных волокон, в артериолах они уже являются основной составляющей сосудистой ткани. Общая зависимость напряжения σ стенок кровеносных сосудов от величины деформации ε представлена на рис.7.4.
В физиологических условиях ткани кровеносных сосудов представляют собой нелинейный упругий малосжимаемый анизотропный материал. При этом механические свойства стенки определяются главным образом свойствами средней сосудистой оболочки, состоящей из коллагена, эластина и гладких мышечных волокон. При низком давлении решающую роль в процессе деформирования играют волокна эластина, при физиологическом давлении – эластина и коллагена, при высоком давлении – коллагена.
Для артериальных сосудов модуль упругости Е изменяется в широких пределах: от 0,06 до 0,7 МПа. Это связано с тем, что под влиянием нервных и химических воздействий клетки гладких мышечных волокон сокращаются. Благодаря сокращению гладких мышц изменяются размеры кровеносных сосудов и их механические свойства. Если до сокращения мышечных волокон модуль упругости Е=0,01 МПа, то при сокращении (стимуляции) Е=0,1 МПа.
С увеличением расстояния от сердца предельная деформация, соответствующая разрыву стенки артерий, уменьшается. Наибольшую прочность имеет общая сонная артерия (предел прочности в осевом направлении σ=2 МПа, предельная деформация ε=0,61).
С возрастом жёсткость стенок увеличивается, а прочность уменьшается.
Для оценки жёсткости стенки можно воспользоваться расчётной схемой тонкостенной цилиндрической оболочки с использованием параметра A, называемого характеристикой растяжимости. Растяжимость – величина, обратная жёсткости, – определяет общее перемещение под действием суммарной единичной силы. Для стенок кровеносных сосудов растяжимость характеризуется изменением объёма в зависимости от давления. Характеристика растяжимости стенки определяется формулой:
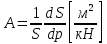
где: S – площадь отверстия (просвета сосуда), р – избыточное давление.
Если материал стенки изотропный, то величина A может быть рассчитана по формуле:
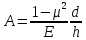
где: Е – модель упругости, μ – коэффициент Пуассона (для несжимаемого материала μ=0,5), d – средний диаметр стенки, h – средняя толщина стенки.
Упругость артериальных стенок обусловлена наличием в них эластина. В грудной аорте содержание эластина достигает 30% от сухой массы стенки, в брюшной аорте и других артериях – до 15%.
Деформацию сосуда в целом можно рассматривать как результат действия давления изнутри на упругий цилиндр.
Рассмотрим цилиндрическую часть кровеносного сосуда длиной l, с толщиной стенок h и радиусом внутренней части r. Сечения вдоль и поперек оси цилиндра показаны на рис.7.5. Две половины цилиндрического сосуда взаимодействуют между собой по сечениям стенок цилиндра (заштрихованные области на рис.7.5,а).
Общая площадь этого «сечения взаимодействия» равна 2hl. Если в сосудистой стенке существует механическое напряжение σ, то сила взаимодействия двух половинок сосуда равна:
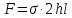
Эта сила уравновешивает силы давления на цилиндр изнутри (они показаны стрелками на рис. 7.5,б).
Читайте также: Офисное кресло loftyhome template ткань серый
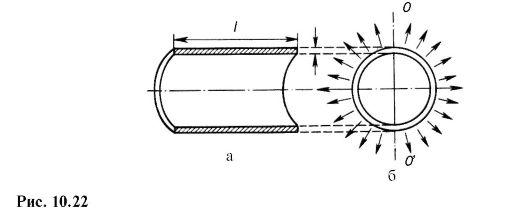
Рис. 7.5. Сечения цилиндрического сосуда
вдоль продольной оси (а) и поперёк оси (б).
Силы направлены под разными углами к горизонтальной плоскости (на рисунке). Для того чтобы найти их равнодействующую, следует просуммировать горизонтальные или вертикальные проекции. Проекцией в этом случае будет прямоугольник, одна сторона которого – это длина сосуда l, а друга – внутренний диаметр сосуда d=2r. Тогда равнодействующая сила давления жидкости будет иметь вид:


Приравнивая (7.2.1) и (7.2.2), получаем , откуда:

Из (7.2.3) видно что в капиллярах (r→0) напряжение почти отсутствует (σ→0).
Формула (7.2.3) выражает зависимость четырёх величин, и поэтому установить некоторые зависимости с помощью этой формулы затруднительно. Кроме того, уравнение (7.2.3) не включает модуль упругости – основную механическую характеристику упругого тела, поэтому целесообразно преобразовать эту формулу в каком-либо приближённом варианте. Например, попытаться найти зависимость p=f(r).
Продифференцируем (7.2.3) как функцию двух переменных (σ=f(p,r):

Продифференцирован уравнение Гука (σ=Eε), получим:

Применительно к цилиндрическому сосуду элементарное изменение относительной деформации можно представить следующим образом:

Подставляя (7.2.6) в (7.2.5) получим:

Приравнивая (7.2.4) и (7.2.7), находим:
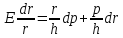
Преобразуем это уравнение:


Если Е велико, а r мало, то тогда , и можно получить приближённую формулу:

Уравнения (7.2.9) и (7.2.10) могут быть использованы для нахождения связи между давлением и радиусом кровеносного сосуда, а также модуля упругости. При решении вопроса о распространении пульсовой волны количественные соотношения получаются также на основе этих уравнений (формула (7.11), лекция 7, часть 1).
Как правило, в стенках артерий больше эластичных волокон и меньше коллагеновых, чем в стенках вен. Вены же, напротив, более богаты коллагеновыми волокнами. Капилляры имеют эндотелиальный слой, но их стенки лишены мышечной и соединительной ткани. Они относительно пассивны, и их поведение определяется преимущественно процессами, происходящими в примыкающих артериолах и венулах. Лимфатические сосуды по строению сходны с венами, но отличаются от них меньшей толщиной и большей проницаемостью.
Мышечные ткани
Мышечные ткани — это ткани, для которых способность к сокращению является главным свойством. Мышечные ткани составляют активную часть опорно-двигательного аппарата (пассивной частью являются кости, соединения костей).
Общими свойствами всех мышечных тканей является сократимость и возбудимость. К данной группе тканей относятся гладкая, поперечнополосатая скелетная и поперечнополосатая сердечная мышечные ткани. Клетки мышечной ткани имеют хорошо развитый цитоскелет, содержат много митохондрий.

Гладкая (висцеральная) мускулатура
Эта мышечная ткань встречается в стенках внутренних органах (бронхи, кишечник, желудок, мочевой пузырь), в стенках сосудов, протоках желез. Эволюционно является наиболее древним видом мускулатуры.
Состоит из веретенообразных миоцитов — коротких одноядерных клеток. Между клетками имеются межклеточные контакты — нексусы (лат. nexus — связь). Благодаря нексусам возбуждение, возникшее в одной клетке, волнообразно распространяется на все остальные клетки.

Гладкая мышечная ткань отличается своей способностью к длительному тоническому напряжению, что очень важно для работы внутренних органов (к примеру мочевого пузыря), сокращается медленно, практически не утомляется. Скелетная мышечная ткань, которую мы изучим чуть позже, такой способностью не обладает — сокращается и утомляется быстро.
Осуществляется сокращение с помощью клеточных органоидов — миофиламентов, которые расположены в клетке хаотично и не имеют такой упорядоченной структуры, как миофибриллы в скелетной мускулатуре (все познается в сравнении, уже скоро мы их тоже изучим).
Особо заметим, что в гладкой мышечной ткани миофиламенты собираются в миофибриллы только во время сокращения. У таких временных миофибрилл не может быть регулярной организации, а значит ни у таких миофибрилл, ни у гладких миоцитов не может быть поперечной исчерченности.
Гладкая мышечная ткань сокращается непроизвольно (неподвластна воле человека). Работа гладких мышц обеспечивается вегетативной (автономной) нервной системой. К примеру невозможно по желанию сузить или расширить бронхи, кровеносные сосуды, зрачок.

Гладкая мышечная ткань называется неисчерченной, так как не обладает поперечной исчерченностью, характерной для поперечнополосатых скелетной и сердечной мышечных тканей.
Скелетная (поперечнополосатая) мышечная ткань
Скелетная мышечная ткань образует диафрагму (дыхательную мышцу), мускулатуру туловища, конечностей, головы, голосовых связок.
В отличие от гладкой мускулатуры, скелетная образована не отдельными одноядерными клетками, а длинными многоядерными волокнами, имеющими до 100 и более ядер — миосимпластами. Миосимпласт (греч. sim — вместе + plast — образованный) представляет совокупность слившихся клеток, имеет длину от нескольких миллиметров до нескольких сантиметров (соответствует длине мышцы).
Внутри миосимпласта находится саркоплазма, снаружи миосимпласт покрыт сарколеммой. Сократительные элементы — миофибриллы (лат. fibra — волоконце) — длинные тяжеобразные органеллы в миосимпласте (около 1400).

Характерная черта данной ткани — поперечная исчерченность, выражающаяся в равномерном чередовании светлых и темных полос на мышечном волокне. Это происходит потому, что границы саркомеров в соседних миофибриллах совпадают, вследствие чего все волокно приобретает поперечную исчерченность. Теперь самое время изучить микроскопическую основу мышцы — саркомер.
Саркомер (от греч. sarco — мясо (мышца) + mere — маленький)
Саркомер — элементарная сократительная единица поперечнополосатых мышц, структурная единица миофибриллы. В состав саркомера (и миофибриллы в целом) входят миофиламенты (лат. filamentum — нить) двух типов, которые обеспечивают сократимость мышечной ткани.
Саркомер состоит из актиновых (тонких) и миозиновых (толстых) филаментов, которые образованы главным образом белками актином и миозином. Сокращение происходит за счет взаимного перемещения миофиламентов: они тянутся навстречу друг другу, саркомер укорачивается (и мышца в целом).

Источником энергии для сокращения служат молекулы АТФ. К тому же невозможно представить сокращение мышц без участия ионов кальция: именно они связываются с тропонином, что приводит к изменению конформации тропомиозина (тропонин и тропомиозин — регуляторные белки между нитями актина), за счет чего становится возможно соединение актина и миозина. При сокращении мышц выделяется тепло (сократительный термогенез).
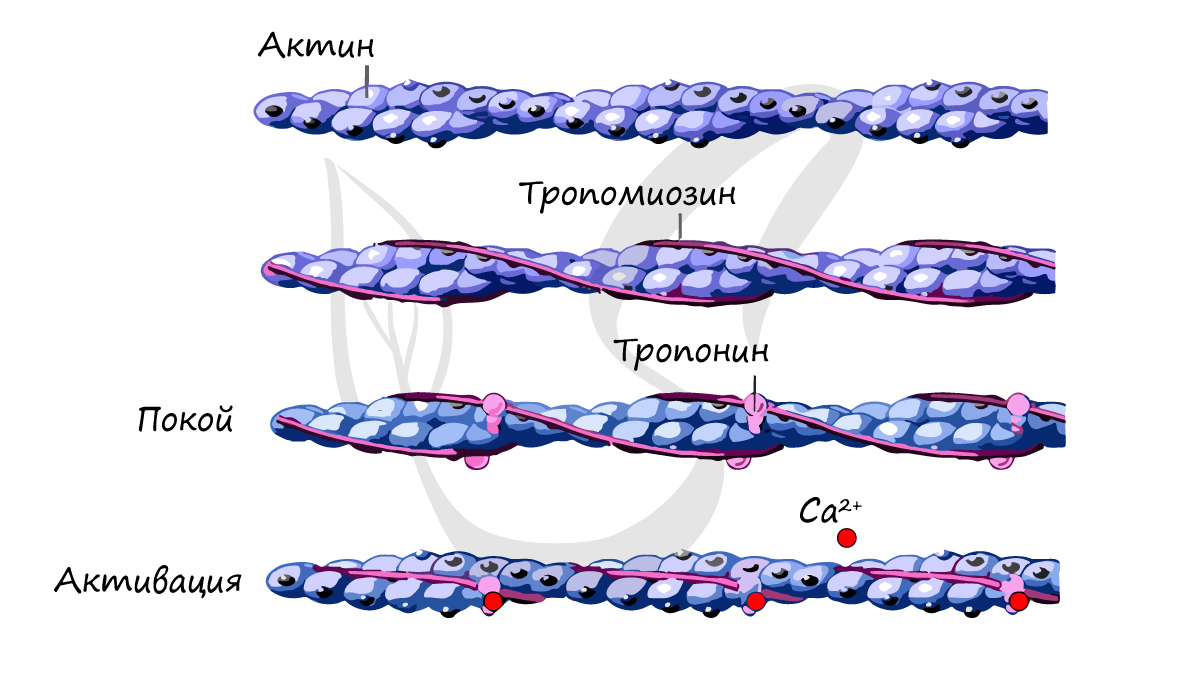
Замечу, что трупное окоченение (лат. rigor mortis) — посмертное затвердевание мышц — связано именно с ионами кальция, которые устремляются в область низкой концентрации (в саркоплазму миосимпласта), способствуя связыванию актина и миозина.
После смерти в мышце перестает синтезироваться АТФ, ее уровень быстро снижается. Как следствие этого перестает функционировать Ca-АТФаза — насос, выкачивающий ионы Ca из саркоплазмы в саркоплазматический ретикулум (мембранная органелла мышечных клеток (сходная с ЭПС), в которой запасаются ионы Ca).
В саркоплазме повышается концентрация ионов Ca — замыкаются мостики между актином и миозином, однако разомкнуться они уже не могут, в связи с чем наблюдается стойкая мышечная контрактура (лат. contractura — стягивание, сужение): конечности очень сложно разогнуть или согнуть.

Вернемся к скелетным мышцам. Имеется еще ряд важных моментов, о которых нужно знать.
В процесс возбуждения вовлекается изолированно один миосимпласт, соседние миосимпласты (волокна) не возбуждают друг друга, в отличие от гладких миоцитов, где возбуждение предается между соседними клетками через нексусы. Скелетные мышцы сокращаются быстро и быстро утомляются (у гладких мышц фазы сокращения и расслабления растянуты во времени, мало утомляются) .
Скелетные мышцы сокращаются произвольно: они подконтрольны нашему сознанию. К примеру, по желанию мы можем изменить скорость движения руки, темп бега, силу прыжка. Мышцы покрыты фасцией, крепятся к костям сухожилиями, и, сокращаясь, приводят в движение суставы.

Сердечная поперечнополосатая мышечная ткань
Сердечная мышечная ткань образует мышечную оболочку сердца — миокард (от др.-греч. μῦς «мышца» + καρδία — «сердце»). Миокард — средний слой сердца, составляющий основную часть его массы. При работе сердечная мышечная ткань не утомляется.

Сердечная мышечная ткань состоит из кардиомиоцитов — одиночных клеток, имеющих поперечную исчерченность. Соединяясь друг с другом, кардиомиоциты образуют функциональные волокна.
Этот тип мышечной ткани удивительным образом сочетает свойства двух предыдущих, изученных нами, тканей (возбудимость, сократимость) и имеет одно новое уникальное свойство — автоматизм.
Автоматизм — способность сердечной мышечной ткани возбуждаться и сокращаться самопроизвольно, без влияний извне. Это легко можно подтвердить, наблюдая сокращения изолированного сердца лягушки в физиологическом растворе: сокращения сердца в нем будут продолжаться несколько десятков минут после отделения сердца от организма.

Места контактов соседних кардиомиоцитов — вставочные диски (в их составе находятся нексусы), благодаря которым возбуждение одной клетки передается на соседние, таким образом волнообразно охватываются возбуждением и сокращаются новые участки миокарда.
Большое число контактов между кардиомиоцитами обеспечивает высокую эффективность и надежность проведения возбуждения по миокарду. Сокращается эта ткань непроизвольно, не утомляется.
На рисунке или микропрепарате узнать данную ткань можно по центральному положению ядер в клетках, поперечной исчерченности, наличию вставочных дисков и анастомозов (греч. anastomosis — отверстие) — мест соединений боковых поверхностей функциональных волокон (кардиомиоцитов).

В норме возбуждение проводится по проводящей системе сердца от предсердий к желудочкам (однонаправленно). Участок сердечной мышцы, в котором генерируются импульсы, определяющие частоту сердечных сокращений — водитель сердечного ритма.
Автоматизм возможен благодаря наличию в миокарде особых пейсмекерных (англ. pacemaker — задающий ритм) клеток, которые также называют водителями ритма. Они спонтанно генерируют нервные импульсы, которые охватывают весь миокард, в результате чего осуществляется сокращение. Именно благодаря водителям ритма сердце лягушки продолжает биться, будучи полностью отделенным от тела.
Ответ мышц на физическую нагрузку
Физические нагрузки приводят к гипертрофии мышц (от др.-греч. ὑπερ- чрез, слишком + τροφή — еда, пища) — в них увеличивается количество мышечных волокон, объем мышечной массы нарастает.

В условиях гиподинамии (от греч. ὑπό — под и δύνᾰμις — сила), то есть пониженной активности, мышцы уменьшаются вплоть до полной атрофии (греч. а – «не» + trophe – питание). В худшем случае волокна мышечной ткани перерождаются в соединительную ткань, после чего пациент становится обездвиженным.

Необходимо отметить, что сердечная мышечная ткань также дает ответную реакцию на чрезмерную нагрузку: сердце увеличивается в размере, нарастает масса миокарда. Причиной могут быть генетические заболевания, повышенное артериальное давление. Гипертрофия сердца — состояние, требующее вмешательства врача и наблюдения за пациентом.
В большинстве случае гипертрофия сердца обратима, а у спортсменов наблюдается так называемая физиологическая гипертрофия (вариант нормы).

Происхождение мышц
Мышцы развиваются из среднего зародышевого листка — мезодермы.

© Беллевич Юрий Сергеевич 2018-2022
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом
- Правообладателям
- Политика конфиденциальности