
ЭЛЕКТРОТОНИЧЕСКИЕ ЯВЛЕНИЯ (синоним электротон физиологический) — изменение возбудимости и проводимости в тканях организма под действием постоянного электрического тока.
Впервые электротонические явления были обнаружены Э. Пфлюгером в 1859 году в эксперименте на нервно-мышечном препарате. Раздражая нервный ствол постоянным электрическим током, он обнаружил, что при замыкании цепи постоянного тока возбуждение в нерве возникало только под катодом, а в момент размыкания — только под анодом. Впоследствии установленная зависимость получила название закона электротона Пфлюгера, или полярного закона (см.). Э. Пфлюгер установил также изменение возбудимости под катодом и анодом во время прохождения постоянного электрического тока через возбудимую ткань. В дальнейшем Б. Ф. Вериго описал снижение проводимости под катодом, получившее название катодической депрессии. Изучая парабиотическое состояние нервов, Н. Е. Введенский обнаружил колебания возбудимости на некотором расстоянии от полюсов действующего постоянного тока, которые были им названы периэлектротоном. Изучение электротонических явлений было продолжено советскими физиологами Л. Л. Васильевым, Д. С. Воронцовым, В. С. Русиновым и др.

При действии постоянным электрическим током на нервное волокно или мышцу непосредственно под катодом (отрицательным полюсом) и в соседних участках ткани повышается возбудимость (катэлектротон), что проявляется снижением пороговой силы тестирующего раздражителя; под анодом (положительным полюсом), а также в соседних участках возбудимой ткани, наоборот, происходит снижение возбудимости (анэлектротон). Если постоянный электрический ток действует на ткань достаточно долго, то характер изменения возбудимости извращается: под катодом начальное повышение возбудимости сменяется понижением (катодическая депрессия), а под анодом пониженная возбудимость постепенно повышается (аподическая экзальтация). С появлением метода прямой регистрации мембранных потенциалов клеток (см. Микроэлектродный метод исследования) стало возможным изучить механизм изменения возбудимости при действии постоянным электрическим током на возбудимую ткань. Прохождение постоянного электрического тока через живые возбудимые клетки сопровождается изменениями поляризации их мембраны как в сторону уменьшения (деполяризация), так и в сторону увеличения (гипер-поляризация), а также изменением величины критического уровня деполяризации. Подобные подпороговые нераспространяющиеся изменения мембранного потенциала, регистрируемые вблизи от раздражающего электрода, получили название электротонических потенциалов. Электротонические изменения мембранного потенциала не связаны с активными изменениями проницаемости для ионов. Электротон имеет чисто физическую природу, определяется пассивными характеристиками мембраны и кабельными свойствами возбудимого нервного или мышечного волокна (см. Нервные волокна). При непродолжительном действии постоянного тока повышение возбудимости под катодом является прямым следствием деполяризации мембраны и приближения уровня порогового потенциала к критическому уровню деполяризации. Снижение возбудимости под анодом происходит в результате увеличения потенциала покоя (гиперполяризация) и отдаления его от уровня критической деполяризации мембраны. Длительное действие постоянного тока на возбудимую ткань приводит не только к сдвигу мембранного потенциала, но и к изменению критического уровня деполяризации. Под катодом происходит уменьшение этих величин, но по абсолютному значению изменения критического уровня деполяризации больше, чем мембранного потенциала, что приводит к возрастанию порогового потенциала и, следовательно, к снижению возбудимости. Под анодом наблюдается увеличение мембранного потенциала и критического уровня деполяризации, но вследствие большего увеличения критического уровня поляризации происходит уменьшение порогового потенциала, что приводит к увеличению возбудимости. При продолжительном действии подпороговым постоянным током критический уровень поляризации под катодом уменьшается настолько, что пороговый потенциал становится больше исходного значения, и в результате снижается амплитуда потенциала действия вплоть до полного его исчезновения, то есть развивается католическая депрессия Вериго (рис.). В соответствии с мембранной теорией возбуждения (см.) причиной наблюдаемых электротонических явлений является инактивация проницаемости ионов натрия через мембрану, обусловленная ее продолжительной деполяризацией. Одновременно значительно повышается проницаемость мембраны для ионов калия.
Изменение возбудимости под полюсами действующего постоянного тока сопровождается изменением проводимости. Под катодом при повышения возбудимости увеличивается скорость проведения нервного импульса по нервному волокну, а во время католической депрессии скорость проведения уменьшается и развивается полный блок проведения. Под анодом снижение возбудимости приводит к падению скорости проведения возбуждения.
Возможность изменения возбудимости нервов и мышц с помощью постоянного электрического тока применяется в экспериментальной нейрофизиологии для изучения мембранных механизмов возбуждения. В неврологической и нейрохирургической практике воздействие полюсами постоянного тока на возбудимые ткани используется для направленного изменения функционального состояния нервов, мышц и центральной нервной системы.
Библиогр.: Латманизова Л. В. Очерк физиологии возбуждения, М., 1972; Общая физиология нервной системы, под ред. П. Г. Костюка, Л., 1979; Ходоров Б. И. Общая физиология возбудимых мембран, М., 1975; Pfluger Е. F. W. Untersuchungen uber die Pbysiologie des Electrotonus, B., 1859.
17.6 Возбудимость, методы её оценки. Изменения возбудимости при действии постоянного тока (электротон, катодическая депрессия, аккомодация).
Возбудимость– это способность возбудимых тканей на действие раздражителя отвечать возбуждением, которое проявляется в виде биоэлектрического процесса и специфической ответной реакции.
Подпороговое пассивное поведение клеточной мембраны называется электротоническим, или электротоном.
Читайте также: Как сделать маску худи из крипипасты из ткани
Закон физиологического электротона(2 закон Пфлюгера) – закон об изменении физиологических свойств тканей при прохождении через них постоянного тока:
При прохождении через ткань постоянного тока в области катода возбудимость и проводимость повышаются, а лабильность снижается(физиологический Кат-электротон), в области анода возбудимость и проводимость понижается, а лабильность повышается – физиологический АН-электротон.
Дополнение Вериго– при прохождении через ткань сильного или длительного постоянного тока повышенная возбудимость в области катода сменяется пониженной – катодическая депрессия, а пониженная возбудимость в области анода сменяется повышенной – анодическая экзальтация. ИТОГ: Возбудимость клетки не зависит ни от величины мембранного потенциала, ни от величины критического уровня деполяризации, а определяется их соотношением: чем меньше между ними разница, тем выше возбудимость и наоборот
Закон Дюбуа-Раймона: Раздражающее действие постоянного тока не зависит ни от силы тока (его плотности), ни от длительности действия(времени), а определяется скоростью его нарастания: чем быстрее меняется ток во времени, тем большей раздражающей силой он обладает.
Аккомодация ткани– явление приспособления (снижения возбудимости) ткани к действию медленно-нарастающего раздражителя.
18.7 Соотношения фаз изменения возбудимости при возбуждении с фазами потенциала действия.
1) местный ответ — физиологический катэлектротон. 2) высоковольтный пик — катодическая депрессия 3) следовая деполяризация — катэлектротон 4) следовая гиперполяризация — анэлектротон
Когда ткань возбуждается генерирует ПД, то временно в ней меняется возбудимость: сначала ткань становится совершенно невозбудимой (абсолютная рефрактерность) любой по силе стимул не способен вызывать в ней новый приступ возбуждения, эта фаза обычно наблюдается во время пика ПД, затем происходит постепенное восстановление возбудимости до исходного состояния (фаза относительной рефрактерности) – в этот момент раздражитель может вызвать возбуждение (генерацию нового ПД), но для этого он должен быть выше порогового (исходного). Затем в фазу следовой негативности возбудимость повышается (супервозбудимость или фаза экзальтации). В этот момент подпороговые раздражители могут вызвать возбуждение. В тканях, в которых ярко проявилась следовая гиперполяризация наблюдается еще одна фаза- субнормальности (фаза сниженной возбудимости).
19.8 Строение и классификация синапсов. Механизм передачи сигналов в синапсах ( электрических и химических) Ионные механизмы постсинаптических потенциалов, их виды.
Синапсом называют образование, обеспечивающее передачу возбуждения от одной структуры к другой.
Синапсы по способу передачи сигнала: I.Химические (передача сигналов с помощью медиатора)
II.Электрические (нексусы) – нет у высш животных и у человека
II. Периферические : 1) Соматические (нервно-мышечные)
2) Вегетативные: 1. Ганглионарные
2. Эффекторные(симпатические и парасимпатические)
Синапсы по характеру действия на воспринимающую структуру: I. возбуждающие
По местоположению выделяют нервно-мышечные синапсы и нейронейрональные, последние в свою очередь делятся на аксосоматические, аксоаксональные, аксодендритические, дендросоматические.
В химическом синапсе выделяют 3 элемента :
пресинаптическую область (пресинаптическая терминаль);
Пресинаптическая областьпредставляет собой демиелинизированный терминальный участок отростка нервной клетки. По форме пресинаптическая терминаль напоминает собой колбу, прилежащую основанием к участку мембраны возбудимой клетки. Наиболее существенной чертой пресинаптической области является скопление пресинаптических пузырьков диаметром 50 мм (везикул), в которых содержится медиатор (химическое соединение, которое является материальным носителем сигнала к реципиентской клетке). Кроме везикул с медиатора в пресинаптической области содержится большое количество митохондрий и лизосом, что свидетельствует о высокой активности обменных процессов в этой области. Кроме того, в этой области обнаружены предшественники медиаторов и продукты их метаболизма.
Синаптическая щель. В химических синапсах составляет от 20 до 50 мм. Здесь содержаться вода, электролиты, олигосахариды, ферменты, участвующие в расщеплении медиатора.
Постсинаптическая область. Включает субсинаптическую мембрану (участок постсинаптической мембраны, имеющий специальный аппарат -рецепторы, характеризующиеся сродством к медиатору. В этой же области имеются химически-чувствительные ионные каналы. Собственно постсинап-тическая мембрана — участок постсинаптической мембраны, которая содержит потенциал-зависимые ионные каналы и на которой происходит генерация постсинаптических потенциалов.
Современные представления о механизмах передачи возбуждения в синапсе.
ПД распространяется по нервному волокну к пресинаптической области.
Изменение проницаемости мембраны пресинаптического образования к ионам Са ++ и поступление Са ++ в пресинаптическом образовании.
Движение везикул с активным медиатором пресинаптической области к пресинаптической мембране и выделение медиатора в синаптическую щель методом экзоцитоза.
Движение медиатора к субсинаптической мембране постсинаптической области и взаимодействие с соответствующими рецепторами мембраны.
Далее возможны два варианта: 1- либо само взаимодействие медиатора с рецептором приводит к активации ионных каналов субсинаптической мембраны (это имеет место в некоторых холинэргических синапсах; 2 — либо вначале активируется вторичный посредник, а затем уже включается цепь биохимических процессов, приводящая к изменению проницаемости ионных каналов. Изменение проницаемости ионных каналов приводит к формированию местных токов и генерации на постсинаптической мембране постсинаптического потенциала.
В зависимости от того по отношению к какому иону меняется проницаемость мембран, возможны два варианта постсинаптических потенциалов — возбуждающий, деполяризационный (ВПСП) и тормозной, гиперполяризационный (ТПСП).
В основе ВПСП лежит повышение проницаемости мембраны по отношению к ионам Nа + , а ТПСП- по отношению к ионам К + иCl — . Характер активности тех или иных каналов определяетсяхимической структуроймедиатора, особенностью рецепторного образования, а также связанного с ним вторичного посредника.
Читайте также: Шелк армани свойства ткани
Электрические синапсы обладают односторонним проведением возбуждения. Это легко доказать при регистрировании электрического потенциала на синапсе: при раздражении афферентных путей мембрана синапса деполяризуется, а при раздражении эфферентных волокон — гиперполяризуется. Оказалось, что синапсы нейронов с одинаковой функцией обладают двусторонним проведением возбуждения (например, синапсы между двумя чувствительными клетками), а синапсы между разнофункциональными нейронами (сенсорные и моторные) обладают односторонним проведением. Функции электрических синапсов заключаются прежде всего в обеспечении срочных реакций организма. Этим, видимо, объясняется расположение их у животных в структурах, обеспечивающих реакцию бегства, спасения от опасности и т. д.
Электрический синапс сравнительно мало утомляем, устойчив к изменениям внешней и внутренней среды. Видимо, эти качества наряду с быстродействием обеспечивают высокую надежность его работы.
9. Законы раздражения. Полярный закон. Физиологический электротон. Като-дическая депрессия.
Полярный закон раздражения (закон Пфлюгера)

При замыкании цепи постоянного электрического тока возбуждение возникает только под катодом, а при размыкании — только под анодом
Доказать правильность закона можно при помощи опыта, в котором меняется расположение катода и анода на поврежденном и неповрежденном участках нерва, иннервирующего мышцу
Прохождение постоянного электрического тока через мембрану вызывает изменение мембранного потенциала покоя.
Так, при замыкании цепи возле катода скапливается его «-» заряд, который уменьшает «+» заряд наружной поверхности мембраны.
Разность потенциалов (между наружной и внутренней поверхностями мембраны) уменьшается, и мембранный потенциал изменяется в сторону уровня критической деполяризации, т.е. формируется деполяризация(возбудимость повышается).
Достижение критического уровня приводит к возникновению пикового потенциала (потенциала действия)
При замыкании цепи возле анода скапливается его «+» заряд.
Он увеличивает «+» заряд мембраны и величину мембранного потенциала
Мембранный потенциал удаляется от критического уровня, превышает значение потенциала покоя и формируетгиперполяризацию (возбудимость уменьшается)
При размыкании цепи, прекращение поступления дополнительного «+» заряда от анода приводит к уменьшению(восстановлению) заряда наружной поверхности мембраны.
Мембранный потенциал, уменьшаясь, приближается к критическому уровню
Формируется деполяризация (возбудимость увеличивается).
После достижения критического значения развивается пиковый потенциал
(Поскольку размыкание происходит после замыкания, а, следовательно, на фоне гиперполяризации и пониженной возбудимости, то для возникновения ПД необходим раздражитель, превышающий по силе пороговый — это анодно-размыкательный эффект)
При размыкании возле катода прекращается накопление его «-» заряда
Заряд наружной поверхности мембраны увеличивается (восстанавливается), мембранный потенциал, увеличиваясь, удаляется от критического уровня и возбуждение не возникает
Закон физиологического электротона
Действие постоянного электрического тока на ткань сопровождается изменением ее возбудимости
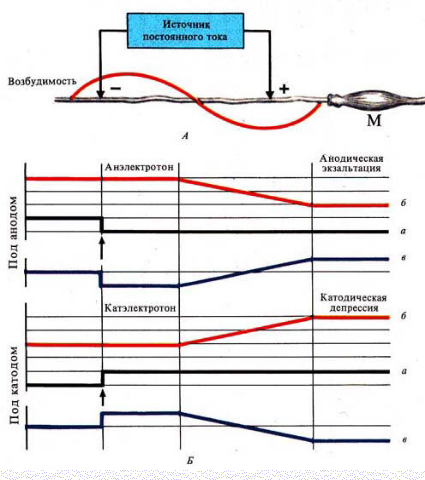

Различают 3 вида физиологического электротона или изменения возбудимости:
Катэлектротон — изменение возбудимости под катодом.
В момент замыкания под катодом формируется деполяризация и возбудимость повышается.
По мере удаления от катода количество его «-» зарядов, а следовательно и выраженность деполяризацииуменьшается.
В результате возбудимость уменьшается, но она остается выше, чем в состоянии покоя
Анэлектротон — изменение возбудимости под анодом.
В момент замыкания под анодом формируется гиперполяризация и возбудимость снижается.
По мере удаления от анода количество его «+» зарядов, а, следовательно, и гиперполяризация уменьшается.
В результате возбудимость увеличивается, но она остается ниже, чем в состоянии покоя
Периэлектротон — обратное изменение возбудимости вне электротонических областей.
В зоне прекращения действия катода возбудимость снижается.
В зоне прекращения влияний анода она, наоборот, увеличивается
При длительном прохождении постоянного тока через ткань происходит извращение измененной возбудимости
При длительном замыкании под катодом происходит:
·инактивация Na проницаемости
·повышение уровня критической деполяризации
·увеличение порога деполяризации
·снижение вначале повышенной возбудимости
Это явление получило название катодическая депрессия
При длительном замыкании под анодом происходит:
·уровень критической деполяризации снижается
·порог деполяризации уменьшается
·повышается вначале сниженная возбудимость
Это явление получило название анодическая экзальтация
Этот закон необходимо учитывать в медицинской практике
С одной стороны, он может быть использован, если требуется заблокировать проведение возбуждения по нервной или мышечной ткани (катодическая депрессия) или повысить возбудимость (анодическая экзальтация)
С другой стороны, необходимо помнить о возможности привыкания к длительному действию раздражителей, в частности, фармацевтических препаратов, влияющих на возбудимость мембраны.
Структура миофибрилл и ее изменения при сокращении.Миофибриллы представляют собой сократительный аппарат мышечного волокна. В поперечнополосатых мышечных волокнахмиофибриллы-разделены на правильно чередующиеся участки (диски), обладающие разными оптическими свойствами. Одни участки анизотропны,т. е. обладают двойным лучепреломлением. В обыкновенном свете они ^выглядят темными, а. в поляризованномг— прозрачными в продольном направлении й непрозрачными в поперечном. Другие участки в Обыкновенной свете выглядят светлыми — они изотропны,т. е. не ; обладают двойным’ лучепреломлением (рис. 34, а) . Анизотропные участки обозначают буквой А,изотропные — буквой I.В середине диска Аразличается светлая полоска Н, посередине диска / — темная «полоска -представляющая собой тонкую мембрану, сквозь поры которой проходят миофибриллы. Благодаря наличию этой опорной структуры параллельно расположенные однозначные диски отдельных фибрилл внутри одного волокна во время сокращения не смещаются по,отношению друг к другу.
Читайте также: Если ткань не прокрашена
Современные представления о структуре миофибриллярного аппарата основываются на исследованиях структуры мышечного волокна-при помощи электронной микроскопии, рентгеноструктурного анализа, фазово-контрастной и интерференционной микроскопии в сочетании с гистохимическими методами.
Установлено, что каждая миофибрилла мышечного волокна диаметром около 1 мкм состоит в среднем из 2500 протофибрилл,представляющих собой удлиненные полимери-, зованные молекулы белков миозина и актина; Миозиновые протофибриллы, или, как, их принято обозначать, нити, вдвое толще актиновых. Их диаметр,примерно 10 нм. В состоянии покоя мышечного волокна нити расположены в миофибрилле таким образом, ^что тонкие длинные актиновые нити входят своими концами в промежутки между толстыми и более короткими миозиновыми нитями (рис. 34, б). Благодаря этому диски / состоят только из актиновых нитей, а диски А — из нитей миозина, а, возможно, еще и другого белка.


Рис. 34. Структура миофибриллы (схема), Показаны диски А и 1-полоски Zи Н(а). Взаимное расположение толстых (миозиновых) и тонких ; (актиновых) нитей в расслабленной (б) й сокращённой (в) миофибрилле.
Светлая полоска Н представляет собой узкую зону, свободную от актиновых нйтей. Мембрана Z,проходя через середину диска /, скрепляет,между собой эти Нити. Важным компонентом ультрамикроскопической-структуры миофибрилл являются также многочисленные поперечные мостики, .соединяющие между собой миозиновые и актиновые нити. При сокращений мышечного волокна указанные нити не укорачиваются, а начинают «скользить» друг по другу: актиновые нити вдвигаются в промежутки между миозиновыми, в результате чего диски / укорачиваются, а диски/1 сохраняют свой размер. Почти исчезает светлая полоска //, так как актиновые нити при сокращении сближаются друг с другом своими концами (рис. 34, в). Причиной «скольжения» является химическое взаимодействие между актином и миозином в присутствии ионов Са 2+ и АТФ. Наблюдается своегорода ч химическое «зубчатое колесо», как 6^,1 протягивающее одну Группу нитей по другой. Роль «зубчиков» в этом процессе приписывают поперечным мостикам, обеспечивающим взаимодействие активных центров белков миозиновых и актиновых нитей.
Это глобулярный белок, состоящий из одного полипептида, который полимеризуется с другими молекулами актина и образует две цепи, обвивающие друг друга (рис. 4-4 А). Такая двойная спираль представляет собой остов тонкого филамента. На каждой молекуле актина есть участок связывания миозина. В покоящемся мышечном волокне взаимодействие между актином и миозином предотвращают два белка — тропонин и тропомиозин (рис. 4-4 Б).

Тропомиозин — стержневидная молекула из двух полипептидов, обвивающихся друг около друга; молекула соответствует в длину примерно семи мономерам актина. Цепи из молекул тропомиозина, уложенные конец в конец, располагаются вдоль всего тонкого филамента. Молекулы тропомиозина частично прикрывают участки связывания каждой молекулы актина, мешая контакту миозина с актином. В таком блокирующем положении молекула тропомиозина удерживается тропонином.
Тропонин — гетеротримерный белок. Он состоит из тропонина Т (отвечает за связывание с одиночной молекулой тропомиозина), тропонина С (связывает ион Са 2+ ) и тропонина I (связывает актин и ингибирует сокращение). Каждая молекула тропомиозина связана с одной гетеротримерной молекулой тропонина, которая регулирует доступ к участкам связывания миозина на семи мономерах актина, прилегающих к молекуле тропомиозина.
Это единое название большой семьи протеинов, имеющих определенные отличия в клетках разных тканей. Миозин присутствует у всех эукариотов. Около 60 лет назад было известно два типа миозина, которые сейчас называют миозин I и миозин II. Миозин II был первым из числа открытых миозинов, и именно он принимает участие в мышечном сокращении. Позднее были открыты миозин I и миозин V (рис. 4-4 В). В последнее время показано, что миозин II участвует в мышечном сокращении, тогда как миозин I и миозин V вовлечены в работу подмембранного (кортикального) цитоскелета. В настоящее время идентифицировано более 10 классов миозина. На рисунке 4-4 Г показано два варианта схемы строения миозина, который состоит из головки, шейки и хвоста. Молекула миозина состоит из двух больших полипептидов (тяжелых цепей) и четырех меньших (легких цепей). Эти полипептиды составляют молекулу с двумя глобулярными «головками», которые содержат оба вида цепей, и длинным стержнем («хвостом») из двух переплетенных тяжелых цепей. Хвост каждой молекулы миозина располагается вдоль оси толстого филамента, а две глобулярные головки выступают по бокам, их иначе называют поперечными мостиками. На каждой глобулярной головке находятся по два участка связывания: для актина и для АТФ. Участки связывания АТФ обладают также свойствами фермента АТФазы, гидролизующей связанную молекулу АТФ.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом