
Для характеристики протекания отдельных ПД используется понятие лабильность. Лабильность – это скорость развития ответа на раздражитель (отдельных ПД). Чем выше лабильность тем больше ПД может произвести ткань в единицу времени. Мерой лабильности является наибольшее количество импульсов, которое ткань может генерировать в единицу времени. Максимальный ритм возбуждения лимитируется длительностью периода абсолютной рефрактерности. Если рефрактерность длиться 0,5 мс, то максимальный ритм составляет 1000 импульсов в секунду и выше.
Самой высокой лабильностью обладает нервная ткань. Она способна генерировать до 1000 импульсов в секунду. Мышечная ткань способна проводить до 500 импульсов в секунду. Наименьшей лабильность обладают синапсы. При этом в максимальном ритме ткань не может функционировать долго. В естественных условиях ткани реагируют на возбуждение в более низком ритме, который может сохраняться длительный период времени. Формируется этот ритм через период супернормальности и поэтому называется оптимальным. Так, у нервного волокна он составляет 500 импульсов в секунду, у мышцы 200 импульсов в секунду.
В ходе ритмического возбуждения лабильность может увеличиваться или уменьшаться. Снижение лабильности ведет к развитию процессов торможения, а ее увеличение определяет свойства ткани усваивать новые более высокие ритмы импульсации. Усвоение более высокого ритма связано с выкачиванием ионов Nа + из цитоплазмы, когда возбуждение проникает внутрь клетки. Таким образом мышцы способны усваивать более частый ритм импульсов поступающих к ним от нервных волокон.
В практической медицине используют эти фундаментальные знания
Местные анестетики блокируют натриевые каналы. Натрий не входит в нервные волокна, значит возбуждение не возникает (боль не распространяется).
Кардиологи управляют входом Кальция (лечение аритмий, гипертонии)
Физиология нервного волокна. Механизмы проведения нервного импульса в миелиновых и безмиелиновых волокнах.
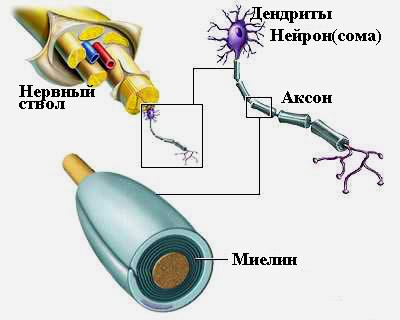
Нервные волокна формируют нерв или нервный ствол, состоящий из нервных волокон, заключенных в общую соединительнотканную оболочку. Поверхность осевого цилиндра нервного волокна покрыта плазматической мембраной, которая выполняет главную роль в возникновении и проведении возбуждения.
Информационная — передача ПД от тела нейрона на периферию и обратно.
Трофическая – трафик нутриентов, органелл.
Генетическая детерминация направления роста.
Классификация нервных волокон.
По строению нервы делятся на:
Миелиновые волокна имеют прерывистую оболочку, которая образована сегментами миелина длиной 1—2 мм. Участок между двумя сегментами называется перехватом Ранвье. На поверхности миелин покрыт Швановскими клетками. Миелин обладает высоким сопротивлением и поэтому выполняет изоляционную функцию и кроме того он принимает участие в обмене веществ нервного волокна. Возбуждение по миелиновому волокну распространяется только через перехваты Ранвье так как там много натриевых каналов.
Безмиелиновые волокна обладают такой же структурой, но не имеют миелина. их поверхность покрыта Швановской клеткой.
Если перерезать нервное волокно, то его периферический конец через некоторое время теряет способность проводить возбуждение и дегенерирует. Перерождение периферического конца нервного волокна изучал Валлер (в 19 столетии) поэтому назвали Валлеровское перерождение. Миелин претерпевает жировое перерождение и превращается в жировые капли. Центральный конец нервного волокна способен регенерировать. На нем образуется колба роста, которая растет к периферии (в сутки от 0,4 до 4,5 мм) и достигает соответствующего органа или ткани. Таким образом, восстанавливается их иннервация. Так, первые признаки восстановления иннервации мышц могут появляться через 5—6 недель.
По направлению передачи информации (центр — периферия) нервы подразделяются на:
Эфферентные по физиологическому эффекту делятся на:
Двигательные (иннервируют мышцы).
Сосудодвигательные (иннервируют сосуды).
Секреторные (иннервируют железы).
По характеру влияния на эффекторный орган нейроны делятся на:
пусковые (переводят ткань из состояния физиологического покоя в состояние активности)
корригирующие (изменяют активность функционирующего органа).
По функциональным свойствам (строению, диаметру волокна, электровозбудимости, скорости развития потенциала действия, длительности различных фаз потенциала действия, по скорости проведения возбуждения) нервные волокна разделили на группы А, В и С.
Группа А неоднородна. Волокна этого типа в свою очередь делятся на подтипы: А-альфа, А-бета, А-гамма, А-дельта.
А-альфа – самые толстые волокна покрыты миелиновой оболочкой. Их диаметр от 12 до 22 мкм. Скорость распространения возбуждения по такому волокну составляет от 70 до 120 мс, ПД самый короткий. Эти волокна проводят возбуждение от моторных нервных центров спинного мозга к скелетным мышцам (двигательные волокна) и от проприорецепторов мышц к соответствующим нервным центрам.
Три другие группы волокон типа А (бета, гамма, дельта) имеют меньший диаметр от 8 до 1 мкм и меньшую скорость проведения возбуждения от 5 до 70 м/с. Волокна этих групп относятся преимущественно к чувствительным, проводящим возбуждение от различных рецепторов (тактильных, температурных, некоторых болевых рецепторов внутренних органов) в ЦНС. Исключение составляют лишь гамма-волокна, значительная часть которых проводит возбуждение от клеток спинного мозга к интрафузальным мышечным волокнам.
А-бета – диаметр от 8 до 13 мкм. Скорость распространения возбуждения от 40 до 70 мс.
А-гамма – диаметр от 4 до 8 мкм. Скорость распространения возбуждения от 15 до 40 мс.
А-дельта – диаметр от 1 до 4 мкм. Скорость распространения возбуждения от 5 до 15 мс.
К волокнам типа В относятся миелинизированные преганглионарные волокна вегетативной нервной системы. Их диаметр — 1- 3 мкм, а скорость проведения возбуждения — 3-18 м/с, ПД длиться 1—2 мс. У эти волокон отсутствует период супернормальности.
К волокнам типа С относятся безмиелиновые нервные волокна малого диаметра — 0,5-2,0 мкм. Скорость проведения возбуждения в этих волокнах не более от 0,5 до 3 м/с, ПД длиться 2 мс. Большинство волокон типа С — это постганглионарные волокна симпатического отдела вегетативной нервной системы, а также нервные волокна, которые проводят возбуждение от болевых рецепторов, некоторых терморецепторов и рецепторов давления.
Таким образом, скорость проведения возбуждения по нерву пропорциональна его диаметру.
Нервы практически не утомляются. Они способны очень длительное время проводить импульсы. Механизмы проведения нервного импульса в
Сравнительная характеристика потенциала действия и локального ответа
ЛО возникает в ответ на действие раздражителя подпороговой силы
ПД возникает в ответ на действие раздражителя пороговой силы
ЛО пропорционален силе подпоpогового раздражителя. ЛО зависит от силы раздражителя до тех пор, пока деполяризация, вызванная этим раздражителем, не достигнет своего критического уровня. В этот момент ЛО перестает быть ЛО, а превращается в ПД.
Читайте также: Роза из подкладочной ткани своими руками
ПД не зависит от силы раздражителя и подчиняется закону «все или ничего»— если раздражитель подпороговой силы, ПД нет, возникает только ЛО, если раздражитель пороговый, то возникает ПД и дальнейшее увеличение силы раздражителя не изменяет величину ПД, она стандартна для каждой клетки.
ЛО может суммироваться до тех пор, пока изменения мембранного потенциала не достигнут КУД
ПД не суммируется, потому, что ПД — это максимальный ответ, на который способна клетка.
ЛО не передается по мембране, потому что при развитии ЛО не происходит пеpезаpядки мембраны (pевеpсии потенциала).
ПД передается по мембране, потому, что при ПД происходит перезарядка мембраны, на короткое время она становится электроположитеьной. Следовательно, возникает разность потенциалов между возбужденным (+) и невозбужденным (-) участками нервного волокна.
Мы рассмотрели изменение потенциала мембраны во время ЛО и ПД. Если вспомнить о том, что возбудимость клетки (способность к ответу) определяется наличием и величиной мембранного потенциала, то становится ясно, что при его колебаниях изменяется и возбудимость во время развития ЛО и ПД. Действительно, ВОЗБУДИМОСТЬ МЕМБРАНЫ в различные фазы одиночного цикла возбуждения не одинакова.
Если принять уровень возбудимости в условиях физиологического покоя за норму, то в ходе развития одиночного цикла возбуждения можно наблюдать ее циклические колебания (рис 11).

Рисунок 11 Изменение возбудимости во время ПД
На рисунке 11: а – нормальная возбудимость, б – повышенная возбудимость во время деполяризации, в – абсолютная и относительная рефрактерность, г — повышенная возбудимость во время деполяризации, д – сниженная возбудимость во время гиперполяризации, е – восстановление возбудимости при восста новлении МПП.
В период развития начальной деполяризации возбудимость незначительно повышается. Во время развития полной деполяризации и инверсии заряда (ПД) возбудимость падает до 0. Время, в течение которого мембрана невозбудима, называется периодом АБСОЛЮТНОЙ РЕФРАКТЕРНОСТИ. В это время даже очень сильный раздражитель не может вызвать возбуждение клетки. Рефрактерность обусловлена инактивацией натриевых каналов — при таком состоянии мембраны натриевые каналы не могут быть открыты (рис 12), кроме того, положительный заряд клетки препятствует входу натрия. В фазе восстановления МП возбудимость повышается, но она еще ниже нормального уровня, это период первичной ОТНОСИТЕЛЬНОЙ РЕФРАКТЕРНОСТИ. Во время реполяризации, когда повышается проницаемость мембраны для ионов калия (рис.12) наступает период повышенной (по сравнению с нормальной) возбудимости — период ЭКЗАЛЬТАЦИИ, клетка еще частично деполяризована.

Рисунок 12 Изменение проницаемости для натрия и калия во время ПД
Во время развития гиперполяризации мембрана снова становится менее возбудима, чем в покое. Этот период повторного снижения возбудимости называется периодом вторичной ОТНОСИТЕЛЬНОЙ РЕФРАКТЕРНОСТИ. Возбуждение может возникнуть только в том случае, если сила раздражения значительно превысит пороговую. После этого возбудимость восстанавливается (МПП) и клетка готова к осуществлению следующего цикла возбуждения.
Фазы измененной возбудимости, сопровождающие развитие ПД, определяют возможность ритмической активности клетки. Максимально возможный ритм активности лимитируется длительностью фазы абсолютной рефрактерности и часто обозначается как ФУНКЦИОНАЛЬНАЯ ЛАБИЛЬНОСТЬ. Чем дольше протекает реполяризация, тем реже способна клетка воспринимать новый возбуждающий импульс, тем ниже функциональная лабильность этой клетки.
У нервных волокон лабильность достигает 1000 импульсов в секунду.
Задумаемся над тем, для чего нужно знать так подробно механизмы возбуждения клеток? Возбудимость тканей может изменяться, поэтому необходимо знать причины изменения возбудимости и результаты изменения, т.е. к чему это может привести. Например, возбудимость гладкомышечных клеток сосудов повышается при накоплении в них ионов кальция (положительный ион), при повышении активности симпатичекой нервной системы. Такое повышение возбудимости приведет к тому, что гладкие мышцы сосудов будут сокращаться в ответ на минимальные дозы сосудосуживающих веществ, например, адреналина, ангиотензина, следовательно, к повышению артериального давления. Возбудимость гладкомышечных элементов сосудов повышается и при накоплении в них ионов натрия, что часто наблюдается при увеличении концентрации гормона надпочечников альдостерона. Повышение возбудимости эндокринных клеток может привести к тому, что в крови увеличится содержание гормонов. Большое значение играет изменение возбудимости клеток миокарда и в нормальной деятельности сердечной мышцы, и в возникновении патологии.
Причиной изменения возбудимости, кроме нарушения внутриклеточной концентрации ионов, могут быть метаболические процессы, связанные, например, с нарушением синтеза АТФ, в этом случае возбудимость может снизиться, т.к. нарушатся процессы реполяризации клеток. Кроме того, ионные каналы это белки, которые чувствительны к действию различных ядов и фармакологических препаратов, следовательно, зная их свойства, можно воздействовать на возбудимость клеток.
Лабильность.
Для характеристики протекания отдельных ПД используется понятие лабильность. Лабильность – это скорость развития ответа на раздражитель (отдельных ПД). Чем выше лабильность тем больше ПД может произвести ткань в единицу времени. Мерой лабильности является наибольшее количество импульсов, которое ткань может генерировать в единицу времени. Максимальный ритм возбуждения лимитируется длительностью периода абсолютной рефрактерности. Если рефрактерность длиться 0,5 мс, то максимальный ритм составляет 1000 импульсов в секунду и выше.
Самой высокой лабильностью обладает нервная ткань. Она способна генерировать до 1000 импульсов в секунду. Мышечная ткань способна проводить до 500 импульсов в секунду. Наименьшей лабильность обладают синапсы. При этом в максимальном ритме ткань не может функционировать долго. В естественных условиях ткани реагируют на возбуждение в более низком ритме, который может сохраняться длительный период времени. Формируется этот ритм через период супернормальности и поэтому называется оптимальным. Так, у нервного волокна он составляет 500 импульсов в секунду, у мышцы 200 импульсов в секунду.
В ходе ритмического возбуждения лабильность может увеличиваться или уменьшаться. Снижение лабильности ведет к развитию процессов торможения, а ее увеличение определяет свойства ткани усваивать новые более высокие ритмы импульсации. Усвоение более высокого ритма связано с выкачиванием ионов Nа + из цитоплазмы, когда возбуждение проникает внутрь клетки. Таким образом мышцы способны усваивать более частый ритм импульсов поступающих к ним от нервных волокон. Например, после длительного похода солдаты возвращаются домой очень усталые, где их встречают с музыкой и у них появляются дополнительные силы. Это явление связано с усвоением мышц более высокого ритма поступающего от нервных центров.