

Более века костный мозг рассматривали частью костей, селезенку — пищеварительной системы, тимус относили к эндокринным железам, лимфоузлы (ЛУ) — к лимфатической системе. Это нашло свое отражение в Базельской (1895), Йенской (1935) и Парижской (1955) анатомических номенклатурах. Но в литературе можно было найти и другие понятия: «кроветворные органы» — млечные пятна, лимфоидные узелки, ЛУ и селезенка (Иванов Г.Ф., 1949); «сосудистые органы», где кровеносное или лимфатическое русло проходит по участку малодифференцированной соединительной ткани — костный мозг, ЛУ, селезенка (Заварзин А.А., 1938). Первая Международная гистологическая номенклатура (1970) содержала раздел «Органы кроветворения» — костный мозг, селезенка и тимус, ЛУ были отнесены к лимфатической системе. В новой Международной анатомической терминологии (Нью-Йорк, 1998) впервые выделен раздел «Лимфоидная система» — костный мозг, тимус, селезенка, глоточное лимфоидное кольцо (миндалины), лимфатический узел, а также лимфоидные узелки и бляшка, термин «лимфатическая система» исключен. В 2008 г. вышла новая Международная гистологическая терминология, она также содержит раздел «Лимфоидная система» — красный костный мозг, тимус, лимфоидный узелок, временное и постоянное периферические лимфоидные скопления, лимфатический узел, селезенка, миндалина. Но красный костный мозг и селезенка являются смешанными по строению кроветворными органами с преобладанием миелоидной ткани, образуются в связи с венозными синусоидами и синусами. Тимус, миндалины закладываются как эпителиомезенхимные скопления клеток, позднее они преобразуются в лимфоэпителиальные органы. ЛУ возникают как переплетения кровеносных и лимфатических сосудов, соединительная ткань между ними преобразуется в лимфоидную. Я считаю, что следует различать «кроветворные органы», а их разделять на смешанные, миелоидно-лимфоидные (красный костный мозг, селезенка) и лимфоидные (тимус, ЛУ, миндалины). Особенности строения миелоидно-лимфоидных органов — главным образом миелоидные (особенно красный костный мозг), экстралимфатические (паренхима не связана с лимфатическим руслом), синусоидные (венозные синусоиды как пути оттока элементов крови), периартериальные (по локализации лимфоидных элементов). Лимфоидные органы можно разделить на следующие группы по разным признакам:
1) функционально-генетическая классификация (пригодна для всех кроветворных органов):
— первичные или центральные (красный костный мозг; тимус);
— вторичные или периферические (селезенка; ЛУ, миндалины и др.);
2) морфо-функциональная классификация:
— экстралимфатические (тимус, миндалины, а также лимфоидные узелки и предузелки);
— лимфатические (ЛУ, а также лимфоидные бляшки и узелки).
Лимфоидные органы содержат посткапиллярные венулы с высокими эндотелиоцитами — пути рециркуляции лимфоцитов между первичными и вторичными лимфоидными органами.
Тимус занимает особое место в системе рециркуляции лимфоцитов (Хэм А., Кормак Д., 1983; Сапин М.Р., Этинген Л.Е., 1996). Наиболее простой вариант:
1) корковое вещество содержит кровеносные капилляры, которые заканчиваются в посткапиллярных венулах с высокими эндотелиоцитами мозгового вещества;
2) отток Т-лимфоцитов происходит из мозгового вещества тимуса через посткапиллярные венулы с высокими эндотелиоцитами.
Более сложный, дискуссионный вариант:
1) лимфоциты поступают в корковое вещество тимуса и уходят из него через капилляры;
2) мозговое вещество тимуса связано с паренхимой вторичных лимфоидных органов через посткапиллярные венулы с высокими эндотелиоцитами.
При этом также возможны варианты соотношения коркового и мозгового вещества тимуса:
1) они не взаимосвязаны (нет перехода лимфоцитов между ними);
2) взаимосвязаны (часть лимфоцитов переходит в мозговое вещество);
3) часть лимфоцитов возникает не в корковом веществе, а в мозговом. Тимус первого варианта близок к ЛУ, во втором варианте это относится к мозговому веществу тимуса, тогда как его корковое вещество напоминает селезенку в части путей поступления лимфоцитов в орган.
Особый вопрос — пути поступления антигенов или иных стимуляторов лимфоцитопоэза в органы, иные очаги кроветворения. По этому признаку их можно разделить на три группы:
1) кровеносные сосуды — красный костный мозг, тимус, селезенка;
2) лимфатические пути — ЛУ, лимфоидные бляшки и узелки;
3) тканевые каналы — миндалина, лимфоидные узелки и предузелки.
В современных Международных анатомической и гистологической терминологиях основополагающим признаком объединения совершенно разных очагов кроветворения в единую лимфоидную систему стала их иммунопоэтическая функция, что представляется необоснованным с морфологической и с генетической точек зрения. Такую совокупность указанных органов и структур можно определять только как лимфоидный аппарат (подобная ситуация с эндокринными железами). Тем более, что красный костный мозг, лимфоидные бляшки, узелки и предузелки не являются самостоятельными органами. К собственно лимфоидным органам можно отнести тимус, ЛУ и, возможно, миндалины.
Лимфоидная ткань кроветворный орган
С эмбриологической точки зрения кровяная ткань берет свое начало в первичном сосудистом ареале, мезенхимальных островках Вольфа и Пандера, от которых происходят также сердце и сосуды. В следующей главе (г) описаны фазы образования кроветворных органов, соответственно первичная мезобластичсская, затем гепатолиенальная и, на последних месяцах, окончательная костно-мозговая фаза кроветворения. Первичная мезенхимальиая клетка, содержащая информацию о кроветворной пролиферации и дифференциации восстанавливает ряды крови, как материнскими стимулами, так и стимулами, отправляемыми дифференцирующимися плодными тканями.
Привлекает внимание многосторонний потенциал первичной мезенхимальной клетки, которая порождает ретикулярную строму селезенки, узлов, печени и костей.
В послезародышевой жизни мезенхимальная клетка дифференцируется в клетки-штамм, различной направленности в костном мозге и лимфатических органах. Эта ориентация определяется в период зачаточного развития и является результатом продолжительной филогенетической эволюции. На протяжении этого развития костно-мозговая миелоидная ткань находится в зависимости от мезенхимальных, сосудисто-соединительнотканных и костных структур, составляющих частный микроклимат, способствующий дифференцированной пролиферации.
Филогенетически лимфоидная ткань развивается по плану иерархической структурной организации, причем исходно она появляется в центральных, а затем и в периферических лимфоидных органах (Берчану). Центральными органами являются вилочковая железа, у животных, и сумка Фабриция — у птиц, при том эти два органа способствуют «целенаправленности» иммунологически компетентных клеток, носящих характер лимфоцитов Т и Б. План их организации общий и создается вокруг эпителиальных образований эндотелиальной природы, берущих свое начало в пищеварительном тракте. Эти образования составляют эпителиальную ретикулярную строму, переплетающуюся с мезенхимальной стромой, как, впрочем, во всех лимфоидных органах.
Дифференциацию иммуных клеток, равно как и способность реагировать на антигенный стимул определяет их эмбриологическое происхождение, в тесной связи с эпителием, внутридермальной природы (Defendi, Good).
Функциональное различие, требуемое для дифференцированного развития по типу миелоидных или лимфоидных клеток определяется этими структурными взаимосвязями с разновидной стромой: эпителиальным эндотелием, для лимфоидной системы и костносуставными образованиями — для миелоидной. Еще не полностью выявлены взаимоотношения, налаживающиеся при создании этих двух систем. Точно установлена роль вилочковой железы в дифференциации лимфоидной системы, но не выведены размеры ее участия в функциях костного мозга. Однако из патологии известно, что опухоли вилочковой железы определяют медуллярную аплазию, но отсутствует доказательство взаимозависимости функций вилочковой железы и костного мозга при нормальном состоянии.
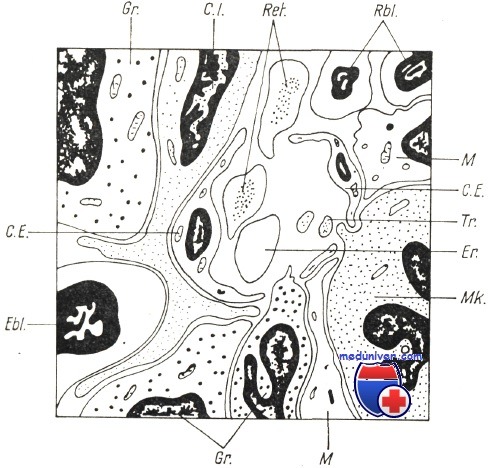
Строение костномозгового синуса.
Ebl—эритробласт; СЕ.— эндотелиальиая клетка; Gr.— гранулоцит; C.I.— интерстициальная клетка; М — макрофаг; Tr — тромбоцит; Mk — мегакариоцит; Еr— эритроцит.
На рисунке изображена функциональная взаимосвязь центральных лимфоидных органов — вилочковой железы и сумки Фабриция — и периферических лимфоидных органов — лимфатических узлов с различной топографической организацией лимфоцитов Т и Б. Лимфоциты Б, а по результатам некоторых исследований и лимфоцаты т находятся и в костном мозге, где образуют мелкие лимфоидные очаги, при этом гуморальный иммунитет находится под контролем лимфоцитов В. Поскольку морфологически клетки-штамм не отличаются от мелких лимфоцитов нам не известно являются ли костномозговые лимфоциты иммунологически компетентными клетками или клетками-штамм.
Проведенными после 1965 г. работами Yoffey и сотр. привели доказательства о том, что лимфоцит это клетка иммуного и в то же время кроветворного восстановления. Методом культур в полунепроницаемых камерах, Григориу и сотр. доказали, что периферические лимфоциты восстанавливают эритробласты. Однако последние данные, полученные в результате анализа колоний в пробирке (Metcalf u Moore) и прижизненно (Мс Colluch) выявили наличие двух различных клеток-штамм, имеющих характеристику лимфоцитов — одну из них в костном мозге, восстанавливающую миелоидные клетки, а другую в лимфоидных органах и периферической крови, восстанавливающую лимфоидные клетки. Эти аргументы выступают в пользу дуалистическоой теории о кроветворении.
Однако не ислючена мысль о способности различной дифференциации той же клетки, в зависимости от структурного, ганглиевого или медуллярного микроклимата, тем самым подтверждая неунистическую гипотезу (Loutit).
В 1979 г. Philips и сотр. впрыскивая клетки-штамм с хромосомными маркерами после облучения проследили возрождение этих клеток у других облученных животных. В связи с этим авторы заключили о наличии полипотентной клетки-штамм (КШп), из которой образуются клетка-штамм для лимфоидных линий Т и В (КШл) и клетка-штамм для миелоидной линии (КШм).
Дифференциацию по той или иной из этих линий определяют условия микроклимата и выделение специфических факторов для организации той или мной из этих линий (Boggs и сотр., Lord и сотр.). При послезародышевой жизни в остальных органах кроветворение не представляется возможным без специфического микроклимата равно как и там, где существуют угнетатели кроветворения. Однако его можно индуцировать при одновременном переносе питательной сети ретикулярной стромы (Humar и сотр.).
Читайте также: Краску для ткани в пензе
Все же функциональные связи и взаимообусловленность возможны в силу собственно структуры костного мозга, по сравнению со структурой ганглиев. В обеих структурах имеется ретикулярная строма с наличием ретикулярных и макрофаговых клеток; существуют также лимфоидные структуры, такие как паренхима ганглиев, но и лимфоидная ткань — в виде мелких фолликулов — и в костном мозге. Нам не известно выполняют ли последние и непосредственную роль в кроветворении посредством определенных общих клеток-штамм или путем стимулирующего трофического влияния на пролиферацию и дифференциацию (Берчану).
Частная патология крови выявит, что заболевания лимфоидного и миелоидного рядов различны. Глубокое поражение стромы и изменение первичной ретикулярной клетки определяет, тем не менее, нарушение обеих систем. Так, при острой недифференцированной лейкемии, рассеянной ретикулосаркоме (PC), нагрузочном ретикулезе, опухолевом гистиоцитозе поражаются обе системы с тяжелыми нарушениями и синдромами недостаточности кроветворения и иммунитета. Структурные связи в костном мозге обусловливают лимфоидные злокачественные метаплазии, смещение миелоидных клеток. При хронической лимфатической, острой лимфобластической лейкемиях или рассеянной лимфосаркоме также наблюдается смещение кроветворной ткани.
Однако существуют патологические аргументы, говорящие о дифференцированной структурной организации этих двух систем. Так, первичная или вторичная аплазия костного мозга не сопровождается аплазией лимфоидной системы; аплазия лимфоидной системы при иммунодефицитных заболеваниях в принципе не поражает кроветворную миелоидную систему. Тем не менее существуют — в настоящее время хорошо известные в иммунопатологии — комплексные иммунодефицитные заболевания, одновременно поражающие обе системы. Так, синдром ретикулярной агенезии и определенные заболевания дисгаммаглобулинемией сопровождаются одновременным нарушением иммуной лимфоидной системы и расстройствами процессов возрождения зернистых и тромбоцитных клеток (Берчану).
Функциональная взаимосвязь в норме, равно как и взаимообусловленность при патологии крови и иммуноклеточной системы говорят в пользу дифференциации миелоидной и лимфоидной систем. Однако эту дифференциацию следует рассматривать, в основном, как результат определенной степени приспособительного отбора по сравнению с условиями среды и структурного микроклимата в послезародышевой жизни. Вместе с тем организационная, генетическая и регенеративная взаимосвязи в зародышевой жизни свидетельствуют о структурном и функциональном единстве организации крови, рассматриваемой как высокодифференцированная мезенхимальная ткань.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Лимфоидная ткань кроветворный орган
2. а) Среди данных элементов — и лимфоциты — особый класс лейкоцитов, обеспечивающий иммунные реакции.
локализацию кроветворения в эмбриональном и в постэмбриональном периодах жизни;
последовательность развития всех форменнных элементов крови,
гистологическое строение центральных органов кроветворения —
строение периферических органов кроветворения — лимфоузлов, селезёнки, лимфоидной системы слизистых оболочек ,
а также общие вопросы иммуногенеза.
20.1. Локализация кроветворения
20.1.1. Кроветворение у эмбриона
мезобластический,
печёночный и
медуллярный .
б) При этом надо заметить, что данное подразделение несколько условно, поскольку
«этапы» не следуют строго друг за другом,
а в значительной степени перекрываются.
I. Мезобластический этап
| Рисунки: А — связь сосудов желточного мешка и тела зародыша; Б — кровяной островок в стенке желточного мешка. |
|
| Локализация | |
1. а) Впервые кроветворение начинается в стенке желточного мешка (1).
б) Здесь появляются скопления мезенхимных клеток — кровяные островки.

1. Центральные клетки (3) кровяных островков
округляются и
внутри сосудов, т.е. интраваскулярно , вступают в т.н. мегалобластический эритропоэз :
2. Образующиеся первичные эритроциты
имеют большой размер ,
часто содержат ядра,
содержат особый вид гемоглобина — т.н. Hb эмбриона.
Позднее в желточном мешке
начинается нормобластический эритропоэз — образовани е обычных эритроцитов ( нормоцитов );
вне сосудов ( экстраваскулярно ) образуются первичные лейкоциты (причём, только гранулоциты );
ч асть стволовых клеток (1-ой генерации) выходит в кровь и переносится в зачат о к печени.
II. Печёночный этап
2. Отличительные черты таковы:
а) п роцесс (в т.ч. эритропоэз) происходит экстраваскулярно — вокруг капилляров, врастающих в печёночные дольки ;
б ) о бразуются все форменные элементы крови ;
имеют обычный размер и
содержат другой (нежели мегалоциты) вид гемоглобина — фетальный (Hb F ).
III. Медуллярный этап
1. Названные стволовые клетки (2-й генерации) оседают в зачатках
тимуса,
лимфоузлов,
селезёнки и
красного костного мозга.
2. а) Все эти органы (а не только красный костный мозг, как следует из названия этапа) включаются в кроветворение на медуллярном этапе ; причём,
кроветворение в них происходит экстраваскулярно,
эритроциты (если они образуются в органе) содержат, в основном, HbF и в меньшей степени HbA ( гемоглобин взрослых);
перечисленные органы остаются органами кроветворения также после рождения.
1. а) Так, вскоре красный костный мозг начинают покидать предшественники Т-лимфоцитов .
б) Своё антиген не зависимое с озревание они заканчива ют в тимусе.
1. а) Вначале в лимфоузлах и селезёнке образуются
все виды форменных элементов крови.
б) Такая способность сохраняется
в лимфоузлах до 15-й недел и развития,
а в селезёнке — до рождени я.
2. а) Затем эти органы (а также лимфоидн ая систем а слизистых оболочек) тоже концентрируются лишь на одной функции (если говорить о кроветворении) —
антиген зависимом созревании В- и Т-лимфоцитов.
з десь образуются лимфатические узелки ;
в последних оседают В- и Т-лимфоциты из, соответственно, красного костного мозга и тимуса ;
п осле антигенной стимуляции соответствующие клоны лимфоцитов вступают в активную пролиферацию и в дальнейшую дифференцировку.
1. Вначале в красном костном мозгу тоже образуются все клетки крови,
а затем, как отмечалось, его начинают покидать предшественники Т-лимфоцитов.
2. Таким образом, у взрослого красный костный мозг сохраняет способность образовывать
все виды клеток крови, кроме Т-лимфоцитов,
а также предшественники Т-лимфоцитов.
| 1. Препарат — срез зародыша курицы. Окраска гематоксилин-эозином. | |
| 1. а) Этот препарат уже встречался нам в п. 6.3.2.2. б) В данном случае он иллюстрирует кроветворение в стенке желточного мешка. | |

20.1.2. Органы кроветворения у взрослых
Теперь уточним местоположение и основную функцию органов кроветворения у взрослого человека.
20.1.2.1. Центральные органы кроветворения
Как уже отмечалось, к центральным органам кроветворения относятся
1. а) Локализация — губчатое вещество
плоских и губчатых костей, а также
эпифизов трубчатых костей.
б) У детей (до 12-18 лет) — также диафизы трубчатых костей
( где потом красный костный мозг замещается на жёлтый костный мозг ) .
2. а) Консистенция — полужидкая;
поэтому из красного костного мозга приготовляют как срезы, так и мазки.
3. а) Функция : в красном костном мозгу, как говорилось выше, происходят все стадии созревания
эритроцитов,
гранулоцитов,
моноцитов,
тромбоцитов и
В-лимфоцитов (нестимулированых).
1. Локализация — за грудиной.
2. Масса: максимальная величина — к 14-15 годам — 35-40г ;
затем масса органа постепенно снижается.
завершается созревание Т-лимфоцитов и происходит их пролиферация ,
о дновременно элиминируются те Т-лимфоциты, которые настроены против собственных антигенных детерминант организма.
20.1.2.2. Периферические органы кроветворения
лимфоидную систему слизистых оболочек,
многочисленны е лимфатические узлы (4) , располагаю щиеся по ходу лимфатических сосудов , и
селезёнку (5).
2. Общая масса лимфоидной ткани во всех этих образованиях сравнима с массой печени или головного мозга.
миндалина языка,
две нёбные миндалины (3.А ),
две трубные миндалины (3.Б),
глоточная миндалина;
одиночные (солитарные) лимфатические фолликулы,
а также их скопления (пейеровы бляшки) (3.В),
в) в стенке червеобразного отростка — лимфатические узелки (3.Г) ,
б) Именно здесь происходит встреча лимфоцитов (В- и Т-клеток) с антигенами — чужеродными молекулами (которые могут находиться либо в растворённом состоянии, либо на поверхности клеток).
I. Два типа кроветворения — миело- и лимфопоэз
образование лимфоцитов (отвечающих за иммунный ответ) стоит несколько особняком от образования других клеток крови
(хотя и то, и другое начинается в одном органе — красном костном мозгу).
2. Соответственно, выделяют два вида кроветворения:
а) миелопоэз — образование всех форменных элементов крови, кроме лимфоцитов, т.е.
эритроцитов,
гранулоцитов,
моноцитов и
тромбоцитов;
II. Два типа кроветворной ткани — миелоидная и лимфоидная
в) Но, как мы уже знаем, в миелоидной ткани , кроме миелопоэза, совершаются и важные события лимфопоэза:
созрева ние В-лимфоцит ов и
начальные стадии созревания Т-лимфоцитов.
2. а) Ткань, в которой происходит дозревание и функционирование лимфоцитов, называется лимфоидной .
III. Два компонента кроветворной ткани — стромальный и гемальный
1. Первый — стромальный компонент . Он может быть представлен:
| ретикулярной тканью (п. 9.4.1) |
— в красном костном мозгу, лимфоузлах и селезёнке , |
| рыхлой соединительной тканью | — в лимфатических фолликулах слизистых оболочек , |
| эпителиальной тканью | — в тимусе. |
2. а) Второй компонент — гемальный:
гемопоэтические (кроветворные) клетки на разных стадиях созревания.
IV. Происхождение кроветворной ткани
2. а) Клетки стромального компонента (будучи разновидностью соединительной ткани) тоже имеют такое же происхождение.
б) Исключение, как уже отмечалось, составляет тимус:
здесь строма долек представлена эпителиальной тканью.
20.2. Постэмбриональный гемоцитопоэз
а) Определив, где проходят процессы кроветворения, рассмотрим теперь сами эти процессы (в постэмбриональный период онтогенеза).
б) Иными словами, рассмотрим пути дифференцировки исходных стволовых клеток в различные виды форменных элементов крови.
Схема — постэмбриональный гемоцитопоэз.
а) Все клетки крови происходят из единого источника — стволовых клеток крови.
б) Соответственно числу разных видов форменных элементов крови, на схеме показаны
6 направлений миелопоэза и
2 направления лимфопоэза.
в) В каждом из этих путей дифференцировки различают 6 классов клеток :
I. стволовые клетки крови,
II. полустволовые клетки,
III. унипотентные клетки,
IV. бласты,
V. созревающие клетки,
VI. зрелые клетки.
20.2.2. Гемопоэтические клетки классов I-III
20.2.2.1. Фрагмент общей схемы
20.2.2.2. Общие свойства клеток классов I-III
Приведённые на схеме гемопоэтические клетки первых трёх классов имеют следующие общие свойства.
попадать в кровь и
после циркуляции вновь выселяться в кроветворные органы.
друг от друга морфологически не отличаются,
а отличаются только по поверхностным антигенам.
часть дочерних клеток полностью идентична материнским (т.е. пополняет пул клеток того класса, к которому принадлежали родительские клетки),
и лишь другая часть подвергается дифференцировке (превращается в клетки последующих классов) .
II. Определение способности к образованию колоний
| Рисунки — А — селезёнка облучённых мышей с колониями гемопоэтических клеток, Б — обычная селезёнка. |
|
| Способность к образованию колоний определяют следующим образом. – | |
1. Мышей-реципиентов облучают такой дозой радиации, при которой погибают все их гемопоэтические клетки.
б) Каждый из них — колония (клон) клеток, развившихся из одной гемопоэтической клетки класса I, II или III.
20.2.2.3. Особенности клеток классов I, II и III
I. Класс I: стволовые клетки крови
2. При этом они являются полипотентными: могут давать начало всем форменным элементам крови.
3. На первом этапе их дифференцировки образуются полустволовые клетки двух видов :
предшественники миелопоэза и
предшественники лимфопоэза.
II. Класс II: полустволовые клетки
Клетки класса II имеют три принципиальные особенности. —
возможности дальнейших превращений для каждой из них уже ограничена.
по двум или более различным направлениям.
1. а) Из вышеприведённых схем следует, что к полустволовым клеткам относятся
предшественники миелопоэза и образующиеся из них клетки следующей стадии развития —
а также предшественники лимфопоэза.
2. Итого — 5 видов клеток, где КоЕ — т.н. колониеобразующие клетки (единицы)
1. В обозначениях полустволовых КоЕ буквы после чёрточки показывают, в какие клетки крови способны дифференцироваться данные КоЕ:
КоЕ-ГнЭ — по двум направлениям —
в нейтрофильные гранулоциты ( Гн) и
в эритроциты ( Э) ;
КоЕ-ГМ — по четырём направлениям —
во все три вида гранулоцитов ( Г ) ( нейтрофилы, эозинофилы , базофилы), а также
в моноциты ( М ) ;
КоЕ-МГЦЭ — по двум направлениям —
в мегакариоциты ( МГЦ) — источники тромбоцитов –
и в эритроциты ( Э ) (напомним: последние могут образовываться также из КоЕ-ГнЭ).
Превращение предшественников миелопоэза в тот или иной из трёх перечисленных видов КоЕ происходит под действием регуляторов:
эритропоэтин (синтезируемый в почках, лёгких и печени) стимулирует образование КоЕ-ГнЭ,
лейкопоэтин — образование КоЕ-ГМ
и тромбопоэтин — образование КоЕ-МГЦЭ.
III. Класс III: унипотентные клетки
2. Поэтому естественно, что (по числу разных видов форменных элементов крови) имеются 8 видов унипотентных клеток — предшественники
1) эритроцитов ( КоЕ-Э ),
2) нейтрофильных гранулоцитов ( КоЕ-Гн ) — и т.д. — вплоть до
7) предшественников В-лимфоцитов и
8) предшественников Т-лимфоцитов.
20.2.2.4. Дополнительные замечания
I. Гомобластический и гетеробластический типы кроветворения
митотические деления ;
одновременно происходящее постепенное сужение потенций развития клеток.
1. Однако в обычных условиях начальные стадии гемопоэза протекают с небольшой интенсивностью,
и содержание клеток классов I-III (а также класса IV) в костном мозгу очень низко.
2. Преобладают же (и значительно) клетки последующих стадий развития — обычно
той последней стадии, на которой клетки ещё способны делиться.
1. Напротив, в экстремальных ситуациях (на пример, после острой кровопотери)
дифференцировочные деления начальных клеток ряда ускоряются,
а расход зрелых форм увеличивается.
2. Это приводит к перераспределению соотношения клеточных форм в костном мозг у :
доля поздних форм снижается,
а доля ранних форм повышается.
3. В таком случае говорят о гетеробластическом типе кроветворения:
в заметном количестве присутствуют клетки нескольких стадий.
Вместе с тем, надо понимать, что принципиальной разницы между этими «типами» кроветворения нет :
в обоих случаях фунционирует вся «вертикаль» гемопоэза ,
и в обоих случаях достигается стационарное состояние (т.е. постоянство количества клеток) каждой клеточной формы.
II. Особенности лимфопоэза
1. а) Что касается лимфопоэза , то на одной из его ранних стадий происходит уникальный процесс ( п. 21.2.2.2) —
перестройка геномной области , кодирующей фрагменты иммуноглобулинов,
и результирующее образование в каждой клетке лишь одного полного гена иммуноглобулина.
2. В итоге, каждая клетка приобретает способность синтезировать и нести на поверхности
иммуноглобулины (антитела) только к одному виду (из множества возможных) антигенных детерминант .
3. В силу случайности процесса геномной перестройки,
образуется большое число разных клеток, отличающихся по своей антигенной специфичности.
1 . а) Последующие деления клеток приводят к образованию клонов лимфоцитов с разной антигеной специфичностью.
б) По некоторым оценкам, число таких клонов близко к 10 7 .
2 . а) Прежде полагали, что дифференциация на клоны происходит только в эмбриональный период.
б) По альтернативным представлениям, это совершается постоянно —
в красном костном мозгу
и, возможно, в тимусе (если в него попадают стволовые или полустволовые клетки).
20.2.3. Гемопоэтические клетки класса IV
20.2.3.1. Фрагмент общей схемы
2,а. Здесь впервые изменяется морфология клеток (за счёт начала специфических синтезов):
от клеток классов I-III (похожих на малые лимфоциты) бласты отличаются
большим размером,
более светлым ядром и светлой цитоплазмой,
появлением в цитоплазме первых продуктов специфических синтезов.
б) Несмотря на последнее обстоятельство, между собой (т.е. «по горизонтали» ) бластные клетки морфологическически практически неразличимы.
3. а) В отличие от предыдущих клеток, бласты не способны к самоподдержанию.
при их делениях образуются только более дифференцированные клетки,
а клетки, подобные родительским, не воспроизводятся.
20.2.4. Завершающие стадии миелопоэза
20.2.4.1. Общая характеристика
а целым рядом последовательно переходящих друг в друга клеток.
не только «по вертикали » — между смежными клетками каждого ряда,-
но и «по горизонтали» — между клетками различных направлений дифференцировки.
зрелых форменных элементов крови.
I. Клетки класса V (созревающие клетки)
б) Базофильный эритробласт
б) Отсюда — резкая базофилия цитоплазмы.
в) Полихромато —
фильный
эритробласт

1. а) Затем создаётся ситуация, когда в цитоплазме присутствуют одновременно
и базофильные компоненты ( рибосомы ),
и оксифильные (новосинтезированный гемоглобин ).
б) Поэтому цитоплазма становится полихроматофильной — приобретает серовато-розовый цвет.
2. а) Полихроматофильные эритробласты — последние из делящихся клеток эритроидного ряда.
г) Оксифильный эритробласт

продолжается накоплени е в цитоплазме гемоглобина ,
а содержани е рибосом снижается.
б) Потому цитоплазма оказывается оксифильной,
т.е. розовой при обычном методе окраски.
2. а) Размер клетки и объём ядра уменьшаются.
б) При этом ядро уплотняется (становится гиперхромным ).
II. Клетки класса VI (зрелые клетки)

(окраска крезиловой синькой)
1. На стадии ретикулоцита клетка у млекопитающих уже не имеет ядра.
2. В цитоплазме появляется зернисто-сетчатая субстанция (п. 8.2) , включающая
ор ганеллы, участвующие в синтезе белка
( свободные рибосомы), и
клетка превращается в эритроцит.
I. Фрагмент общей схемы
II,а. Промиелоциты — первые клетки класса V
| 1. Промиелоциты (нейтрофильные, эозинофильные и базофильные) ещё друг от друга практически не отличаются : | ||
 |
|
|
| нейтрофильные | эозинофильные | базофильные |
| 2. Все они имеют следующие признаки: | ||
округлые ядра,
голубую (при окраске по Романовскому) цитоплазму,
а в цитоплазме — азурофильную зернистость ; её образуют первичные (неспецифические, или промиелоцитарные) гранулы (п. 8.3.2.1).
II,б. Миелоциты — первые клетки, содержащие специфические гранулы
в нейтрофильных
миелоцитах —
в эозинофильных миелоцитах —
в базофильных миелоцитах —
1. а) Ядра у миелоцитов по-прежнему округлые.
б ) Дальнейшее же созревание клеток проявляется, главным образом, в изменении структуры и формы ядра.
2 . а ) В результате этих изменений ядра клеток потеряют способность к делениям.
б ) Поэтому миелоциты —
последние клетки гранулоцитопоэтических рядов, способные делиться.
3. Как полихроматофильные эритробласты, в норме они являются
преобладающим типом среди клеток соответствующего ряда.
II,в-д. Клетки завершающих стадий развития
не делятся,
обнаруживаются в крови,
содержат ядро специфической формы.
2. При этом обнаруживаемые в крови метамиелоциты называются юными гранулоцитами.
3. А форма ядра закономерно меняется —
у метамиелоцитов она бобовидная,
у палочкоядерных клеток ядро похоже на толстую изогнутую палочку без перемычек;
у сегментоядерных клеток ядро состоит из нескольких сегментов, разделённых узкими перетяжками.
ядро — круглое, большое;
в цитоплазме нет гранул.
I. Созревающие клетки класса V

2. а) Поэтому объём ядра и клетки в целом значительно увеличивается.
б) Это позволяет легко обнаружить данные клетки (и зрелые мегакариоциты) среди гемопоэтических клеток.
2. А в цитоплазме появляется демаркационная мембранная система (вид им ая под электронным микроскопом):
она разделяет цитоплазму на фрагменты — будущие «тромбоциты» .
II. Зрелые форменные элементы класса VI

б) П осле этог о фрагменты цитоплазмы отделяются в виде тромбопластинок («тромбоцитов» ) .
2. Остающаяся ядросодержащая часть мегакариоцита может
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом
- Правообладателям
- Политика конфиденциальности
Мастерица © 2023
Информация, опубликованная на сайте, носит исключительно ознакомительный характер